В состав ДНК входят тимин, цитозин, аденин и гуанин.
В состав РНК – урацил, цитозин, аденин и гуанин.
НК содержат пентозы.
Азотистые основания представляют 2 класса пиримидины, пурины.
В первичной структуре ДНК нуклеотиды связываются между собой фосфодиэфирными мостиками, которые связывают 5’ гидроксильную группу одного нуклеотида и 3’ гидроксильную группу другого нуклеотида, т.е. состав молекулы ДНК представляет сахарный остаток фосфорной к-ты.
| Белки | Нуклеиновые к-ты |
| Молекулярная масса 100кД | Несколько тысяч кД |
| 4 уровня структурной организации | 3 уровня структурной организации |
| Выполняют много ф-ций | 1ф-ция – хранение и передача генетической информации. |
| Характерна разнообразная структура | 1структура – 2правазакрученная спираль. |
Исследование Э.Чаргоффа и других ученых сделавших след. выводы: 1)нуклеотидный состав ДНК разных тканей одного и того же вида одинаков. 2)нуклеотидный состав ДНК у разных видов различен. 3)нуклеотидный состав не зависит от возраста и питания. 4)в составе ДНК число остатков аденина всегда равно числу остатков остатков тимина, а число остатков гуанина равно числу остатков цитозина.
Вторичная стр-ра ДНК.В 1944 Эвери, Маклеод, Маккарти доказали, что ДНК – носитель генетической информации. 1953 – Уотсон и Крик предложили теорию двойной спирали, которая позволяет объяснить механизмы хранения, трансформации и реализации генетической информации.
Нуклеотиды связаны в молекуле ДНК фосфорноэфирной связью. Взаимодействие двух полинуклеотидных цепочек молекулы ДНК обеспечивается комплементарностью. Основную роль в этом играют водородные связи, так же важную роль играют гидрофобные взаимодействия, стэкинг взаимодействия, дипольные взаимодействия π-связей.
Молекулы ДНК представляют двойную правозакрученную спираль. При этом плоскости азотистых оснований лежат перпендикулярно оси спирали. На внешней стороне двойной спирали расположены остатки дезоксирибозы и фосфорной к-ты.
Полиморфизм – это разнообразие формы молекулы ДНК. Стандартная конформация мол-лы ДНК – это В-конформация. Основания располагается перпендикулярно оси спирали на расстоянии 0,36 нм. А-конформация получается из В-конформации при ее дегидротации. Z -конформация – это левозакрученная спираль. С-тип.
Денатурация происходит при действии химических факторов и физических факторов. Денатурированный белок приобретает свою нативную структуру – ренатурация. Денатурация при действии температуры – плавление ДНК. Температура плавления зависит от нуклеотидного состава.
Гиперхромный эффект – увеличение оптической плотности ДНК при плавлении ДНК. Максимальное повышение оптической плотности раствора ДНК при длине волны 260 нм приблизительно равно 80%, при полном распаде ее до мононуклеотидов.
Хроматин состоит из многих витков ДНК, присоединенных к гистонам – белкам основной природы. В период между делениями, хромосомы практически неразличимы в световом микроскопе как обособленные структуры, хотя материал, из которого они состоят, окрашивается некоторыми основными красителями и потому назван хроматином. На этой стадии хромосомы представляют клубок длинных тонких нитей.
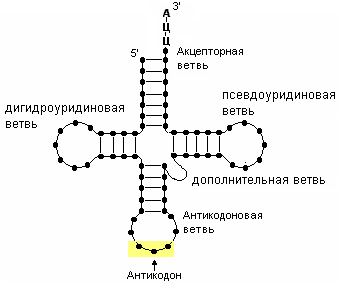 17. Нуклеосомы- структурная единица хроматина, связываются линкерными участками. Последовательность нуклеосом, соединенная гистоновым белком H1, формирует нуклеосомную нить. Гистоны – ядерные белки, на них навивается молекула ДНК, выполняющие две основные функции-они участвуют в упаковке нитей ДНК в ядре и регулируют такие ядерные процессы, как транскрипция, репликация и репарация. Существует пять различных типов гистонов H1/Н5(линкерные), H2A, H2B, H3, H4. Классификация РНК: информационная- Открыта в 1961 году Жакобом и Мано. Она составляет всего 2-3% от общего количества РНК клетки. Н есет информацию о первичной структуре белка служат матрицами для биосинтеза полипептидных цепей. Они содержат линейную последовательность кодонов, которые и определяют первичную структура белка.иРНК –одноцепочечные молекулы. Одна молекула иРНК может кодировать одну или несколько полипептидных цепей.Если иРНК несет информацию об одной полипептидной цепи, то ее называют моноцистронной (эукариот), если о двух или более – полицистронной (прокариот). В нерабочем состоянии м-РНК свернута в клубок, связана с белком; а во время функционирования цепь расправляется. Матричные РНК синтезируются на ДНК в ядре. Процесс называется транскрипция (списывание); транспортная- обеспечивает расшифровку ген. инф., записанной в иРНК и доставку АК к месту синтеза белка. На 3’-конце всех тРНК находится тринуклеотидная последовательность Ц-Ц-А. Более половины оснований тРНК образуют внутрицепочечные пары по принципу комплементарности. Т.о., формируется вторичная структура- клеверный листок.
17. Нуклеосомы- структурная единица хроматина, связываются линкерными участками. Последовательность нуклеосом, соединенная гистоновым белком H1, формирует нуклеосомную нить. Гистоны – ядерные белки, на них навивается молекула ДНК, выполняющие две основные функции-они участвуют в упаковке нитей ДНК в ядре и регулируют такие ядерные процессы, как транскрипция, репликация и репарация. Существует пять различных типов гистонов H1/Н5(линкерные), H2A, H2B, H3, H4. Классификация РНК: информационная- Открыта в 1961 году Жакобом и Мано. Она составляет всего 2-3% от общего количества РНК клетки. Н есет информацию о первичной структуре белка служат матрицами для биосинтеза полипептидных цепей. Они содержат линейную последовательность кодонов, которые и определяют первичную структура белка.иРНК –одноцепочечные молекулы. Одна молекула иРНК может кодировать одну или несколько полипептидных цепей.Если иРНК несет информацию об одной полипептидной цепи, то ее называют моноцистронной (эукариот), если о двух или более – полицистронной (прокариот). В нерабочем состоянии м-РНК свернута в клубок, связана с белком; а во время функционирования цепь расправляется. Матричные РНК синтезируются на ДНК в ядре. Процесс называется транскрипция (списывание); транспортная- обеспечивает расшифровку ген. инф., записанной в иРНК и доставку АК к месту синтеза белка. На 3’-конце всех тРНК находится тринуклеотидная последовательность Ц-Ц-А. Более половины оснований тРНК образуют внутрицепочечные пары по принципу комплементарности. Т.о., формируется вторичная структура- клеверный листок.
В ней выделяют: 1)дигидроуридиловую ветвь; 2)псевдоуридиловую ветвь, содержащую минорные азотистые основания псевдоуридина; 3) антикодоновую ветвь, в центре которой находится антикодон (комплементарен кодону); 4)дополнительную ветвь; 5)акцепторную ветвь с универсальной 3'-концевой последовательностью ЦЦА, служащей акцептором остатка аминокислоты; рибосомальная- входит всостав рибосом, определяют их структуру, обуславливает количество синтезируемого белка, занимают большую часть РНК в клетке 80-85%. Рибосомы представляют собой - крупный рибонуклеопротеидный комплекс с молекулярной массой около 2,5 мДа, состоящий из белков и молекул рРНК. Уэукариот они представлены 80S частицами, у прокариот – 70S . Рибосомы всех известных организмов построены из большой и малой субчастиц: прокариотические – 30S и 50S, а эукариотические – 40S и 60S. рРНК играют важную роль в структуре и биосинтетической функции рибосом.
Макромолекулярная структура РНК: линейный полимер нуклеотидов. Каждый мономер - нуклеотид - представляет собой фосфорилированный N-гликозид, построенный из остатка пятиуглеродного сахара - пентозы, несущего фосфатную группу (сложноэфирная связь) и азотистое основание при первом углеродном атоме (N-гликозидная связь). Азотистых оснований в РНК четыре вида: два пуриновых - аденин (А) и гуанин (G) -и два пиримидиновых - цитозин (С) и урацил (U).
18.Функции нуклеиновых кислот: хранение, передача и реализация генетической информации, так же участвуют в катализе некоторых химических реакций, осуществляют регуляцию реализации генетической информации, выполняют структурные функции и т.д. Роль хранителя генетической информации у большинства организмов (эукариот, прокариот, некоторых вирусов) выполняют двухцепочечные ДНК. Только у некоторых вирусов хранителем генетической информации являются одноцепочечные ДНК или одноцепочечные, а также двухцепочечные РНК. Генетическая информация записана в генах. Гены несут информацию о первичной структуре белков и о структуре некоторых типов РНК. Генетическая информация передается от родителей к потомкам. В результате реализации генетической информации происходит синтез белка: ДНК ® РНК ® белок . Основная догма в процессе размножения клеток генетическая информация передается от одного поколения клеток другому. Все клетки получают одинаковую информацию.
РЕГУЛЯТОРНЫЙ ГЕН (ген-регулятор)- это ген, регулирующий производство других генов. Регуляторные гены активизируют и угнетают группу соседних генов, называемую ОПЕРОН, которая функционирует как единое целое. Встречающиеся обычно у БАКТЕРИЙ, опероны отвечают за формирование ФЕРМЕНТОВ, регулирующие различные процессы обмена веществ. Оперон – группа следующих друг за другом генов, экспрессия которых одновременно регулируется определёнными участками молекулы ДНК-оператор.
Это возможно вследствие того, что перед делением клетки осуществляется репликация ДНК, в результате образуются две идентичные молекулы ДНК, которые и передаются потомкам.
Транскрибируемый участок ДНК ограничен со стороны 3’-конца промотором – участком с которым связывается РНК-полимераза, со стороны 5’-конца – терминатором – участком, в котором прерывается синтез РНК. Последовательность ДНК, ограниченная промотором и терминатором, представляет собой единицу транскрипции – транскриптон. Транскриптон прокариот называют опероном.
Механизм репликации. В основе лежит принцип комплементарности. В результате образуются дочерние молекулы НК, нуклеотидные последовательности которых идентичны между собой и с материнской молекулой. Типы репликации: 1)Полуконсервативный – вновь синтезированная молекула НК состоит из одной материнской и одной дочерней полинуклеотидных цепей НК.2)Консервативный – вновь синтезированная молекула НК состоит только из дочерних полинуклеотидных последовательностей.3)Дисперсный – вновь синтезированная полинуклеотидная цепь НК состоит из фрагментов дочерних и материнских полинуклеотидных последовательностей. Дляпрокариот и эукариот характерен полуконсервативный тип репликации. У вирусов встречаются все три типа репликации.
Механизм репликации требует наличие следующих компонентов: 4-х нуклеотидов; АТФ и Mg ; ДНК-полимераза(1-го типа, присоединяется очередной нуклеотид к 3’-гидроксильной группе растущей полинуклеотидной цепи); затравка (матричная молекула ДНК с разорванной в нескольких местах цепью).
19. Экспрессия генов включает транскрипцию (коп.инф. с ДНК на синтезируемую РНК) и послед. трансляцию (синтез на матрице РНК соотв. белка), репликацию (удвоение мол. ДНК при делении кл-ки). Возможен поток инф. и в напр. от РНК к ДНК, этот процесс носит название обратная транскрипция. Инф. от белков НК не передается. Ген - единица насл. инф., занимающая опред. положение в геноме и контрол. выполнение опр. ф-ций в ор-ме. Гены – уч-ки ДНК, кодир. полип.цепи или РНК. Белок, сост. из разл. полип.цепей, код. неск. генами.Гены могут быть уникальными – предст. одной копией – и повт-ся.Все гены можно разделить на две группы: 1)Конститутивные гены («гены домашнего хозяйства») постоянно включены: они функц. на всех стадиях онтогенеза и во всех тканях (тРНК, рРНК, ДНК-полимеразы, РНК-полимеразы). 2) Индуцибельные гены («гены роскоши») могут включ. и выкл-ся. Они контр.ход онтогенеза и гены, опред. ст-ру и ф-ции комп-в кл-ки и целостного орг-ма. Включ. индуц. генов назыв. индукцией, выкл. – репрессией. Генет. код св-ва:1)Триплетность. Кодон сост. из посл-ти, состоящей из трех нуклеотидов. Число кодирующих послед.из четырех нуклеотидов по три равно 43=64, из них 61 – смысловых кодонов, - 3 нонсен-кодона. 2)Выражденность. Каждый из 61 смысловых триплетов кодирует вкл. в белок одну из 20 станд. АК. Вкл. одной и той же АК в белок определяет неск. кодонов. 3)Специфичность (однозначность).Каждому кодону соотв. только одна опред. АК. 4)Неперекрываемостькода след.за иниц. кодоном АУГ триплеты читаются посл-но не перекрываясь без каких либо пропусков вплоть до стоп-кодона, на кот. синтез полип. цепи завершается. 5)Универсальность кода т.е. смысл кодов одинаков для всех орг-в . В пр-се транскр. выделяют три стадии: инициацию, элонгацию и терминацию. На стадии иниц. РНК-полимераза, взаимод. с промотором, вызывает расхождение цепей ДНК и начинает синтез мол. РНК. В ходе элон. новая синтезир. цепь РНК образует короткие отрезки гибридной двойной спирали ДНК-РНК, кот.необходимы для правильного считывания цепи ДНК. Как только РНК-полимераза достигнет терминирующих послед., запускается последняя стадия – терминация. По ее завершению происх. освоб. вновь синтез.цепи РНК.
20. Механизм трансляции. Разл-ют несколько стадий биосинтеза белка: активация АК, инициация, элонгация и терминация. Синтез белка хар-тся тем, что между матрицей (иРНК) и продуктом (белком) нет комплем-го соответствия. Для расшифровки нукл. послед.необходим генет. код. Он устан. соотв-ие между нукл. послед.иРНК и полип. цепью. Единицей ген. кода является кодон. Кодон предст. собой послед-ть, сост. из трех нуклеотидов, т.е. триплет. Три кодона служат сигналом для остановки синтеза полип.цепи - терминирующие, или нонсенс-кодоны. Каждому кодону (из 61) соотв. строго опред. АК. Тран-ия иРНК начинается с иниц-го триплета – АУГ. Далее расшифр-ся каждый послед.триплет в напр-ии от 5’-конца молекулы иРНК к 3’-концу. Закан-ся синтез полип-да на 1 из 3-х термин-их кодонов. Синтез полип.цепи нач-ся с N -конца.Активация АК 1.На этой стадии каждая из 20 АК присоединяется к опр-ой тРНК. При этом исп-ся энергия АТФ. 2.Эти р-ции катализ-ся 20 разл. аминоацил-тРНК-синтетазами. 3.Аминоацил-тРНК-синтетазы способны узнавать 3 разл. субстрата: АТФ, АК-у и тРНК.4.Каждая аминоацил-тРНК-синтетаза способна узнавать только 1 опр. АК и соотв-ие ей тРНК, в кл-е имеется неск. изоакцепторных тРНК5.Они прис-ют АК остаток к 2’- или 3’-гидроксильной группе 3’- концевого нукл-да. 6.Р-ция прот. в 2 стадии
Инициация белкового синтеза. Иниц-ия нач-ся с присоед. к малой субъ-це рибосомы иРНК и 1-ой аминоацил-тРНК (аа-тРНК), антикодон кот.ком-рен иниц. кодону АУГ. После связывания антикодона тРНК с иниц. кодоном происх. присоед. большой субъединицы рибосомы. Образ. инициирующий комплекс, в кот. иниц. а-тРНК находится в Р (пептидильном)-центре, а А (аминоацильный) – центр свободен.Для осущ. инициации трансляции необходима энергия. Ее поставляет ГТФ. Энергия высв-ся при гидролизе ГТФ до ГДФ и фосфата. Элон. белкового синтеза. В пр-се элон. происходит наращивание полип.цепи. В А-центре происх. связывание след.аа-тРНК, антикодон кот. ком-рен след. кодону. Далее происх.образов. пепт. связи за счет пептидилтрансферазной р-ции. В рез-те образ.дипетидил-тРНК. Катализирует этот процесс рРНК большой субъединицы рибосомы.
Далее рибосома пер-ся на один кодон в напр. от 5’- к 3’-концу мРНК. Дипептидил-тРНК из А-центра перем-ся в Р-центр А-центр освобождается, здесь оказывается следующий кодон. Иниц. тРНК покидает рибосому. Рибосома готова к новому циклу элон. Далее присоед. третий АК остаток, потом – четвертый и т.д. В пр-се транс.затрачивается энергия, освоб. при гидролизе ГТФ до ГДФ и фосфата. Терминация.Терм. белового синтеза наступает, как только в А-центре окажется один из термин. кодонов: УАГ, УГА, УАА. В этом пр-се участвуют специф. белки – ф-ры терм. В рез-те терм. происх.гидролит. отщепление полипептида от тРНК, тРНК отдел. от рибосомы, рибосома диссоциируют на субъединицы. Поставщиком энергии для терминации синтеза белка явл. ГТФ. Рибосомы. Риб-мы предст. собой - крупный рибонуклеопротеидный комплекс с молек-ой массой около 2,5 мДа, сост-ий из белков и молекул рРНК. Риб-мы прокарио-их и эукариот-их организмов разл-ся по размерам.
21.Хим с-в биомембран. Стр-ра и ф-ции липидов. Мембрана (лат.перепонка)- тонкая плаская ст-ра толщиной 7-10нм состоящая из бислоя фосфолипидов и отделяющая цитоплазму клетки от внешней среды и клеточные органеллы др. от др. Состав мембран зависит от их типа и функций, однако основными составляющими являются Липиды и Белки, а также Углеводы (небольшая, но чрезвычайно важная часть) и вода (более 20% общего веса).
Липиды – группа органических соединений, которая характеризуются: нерастворимостью в воде; растворимостью в неполярных растворителях (эфир, хлороформ, бензол); содержанием высших алкильных радикалов; распространенностью в живых организмах. Функции: 1. основные компоненты биологических мембран (влияют на их проницаемость, участвуют в передаче нервного импульса, создании межклеточных контактов); 2.эффективный источник энергии (непосредственное использование либо запасы жировой ткани); 3.создание термоизоляционных покровов у животных и растений; 4.защита органов и тканей от механических воздействий.
КЛАССИФИКАЦИЯ ЛИПИДОВ
A. Простые липиды: сложные эфиры жирных кислот с различными спиртами. 1.Глицериды представляют собой сложные эфиры трехатомного спирта глицерина и высших жирных кислот. 2. Воска: сложные эфиры высших жирных кислот и одноатомных или двухатомных спиртов.
Б. Сложные липиды: сложнее эфиры жирных кислот со спиртами, дополнительно содержащие и другие группы. 1.Фосфолипиды: липиды, содержащие, помимо жирных кислот и спирта, остаток фосфорной кислоты. В их состав часто входят азотистые основания и другие компоненты: а)глицерофосфолипиды (в роли спирта выступает глицерол); б) сфинголипиды (в роли спирта - сфингозин). 2. Гликолипиды (гликосфинголипиды). 3.Стероиды.4.Другие сложные липиды: сульфолипиды, аминолипиды. К этому классу можно отнести и липопротеины.
Фосфолипиды представляют 3-атомный спирт глицерин, у которого 2-ОН группа этерифицирована остатками длинноцепочечных жирных кислот, а 3-ОН группа этерифицирована остатками фосфорной кислоты. В состав фосфолипидов входят также азотсодержащие соединения холин, этаноламин или серии. Фосфолипиды — амфифильные вещества. Они состоят из полярной «головки», в состав которой входит глицерин или другой многоатомный спирт, отрицательно заряженный остаток фосфорной кислоты и часто несущая положительный заряд группа атомов, и двух неполярных «хвостов» из остатков жирных кислот. Главная особенность фосфолипидов состоит в том, что «головка» у них гидрофильна, а «хвосты» гидрофобны. Это позволяет при нахождении в толще водной среды образовывать бислой — двойной слой фосфолипидных молекул, где гидрофильные головы с обеих сторон соприкасаются с водой, а гидрофобные хвосты упрятаны внутрь бислоя и тем самым защищены от контакта с водой.
Дата: 2019-07-24, просмотров: 383.