Обратимся к новому типу нейронных взаимодействий. Чтобы их было проще понять, я пользуюсь упрощенной схемой нейронов. Как изображено на диаграмме, нейрон А посылает все свои аксонные проекции к нейрону В , выделяя возбуждающий нейромедиатор, он изображен значком «плюс». А кружком на нейроне В обозначено клеточное тело и все дендритные веточки.
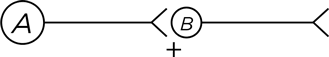
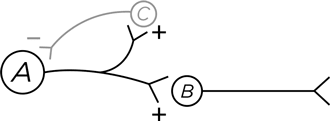
Вот другая схема. Нейрон А стимулирует нормальным образом нейрон В . Но одновременно он стимулирует и нейрон С . В этом нет ничего необычного, ведь нейрон может делить свои аксонные окончания между двумя целевыми нейронами, возбуждая их обоих. И что же делает нейрон С ? Он посылает ингибиторные проекции обратно к нейрону А , формируя петлю отрицательной обратной связи. Вспомним, как мозг любит контрасты: вопить что есть мочи, когда у него есть что сказать, и хранить напряженное молчание, когда сказать нечего. И здесь те же контрасты, но на более высоком уровне. Нейрон А генерирует несколько потенциалов действия. И как яснее всего обозначить, что разговор окончен? Глухо замолчать, что и делается с помощью петли обратной связи. Таким образом, границы сигнала на временной оси становятся более четкими[1539]. Нейрон А «определяет», насколько сильной является эта отрицательная обратная связь, – по тому, сколько из своих 10 000 нейронных отростков он посылает к нейрону С вместо В .
Повысить четкость сигнала во времени можно и по-другому.
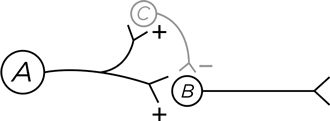
Как и в предыдущем случае, нейрон А активирует В и С . Нейрон С посылает тормозящий сигнал на нейрон В – ясно, что он придет с некоторой задержкой, уже следом за активирующим сигналом от А (потому что в петле А /С /В две синаптические передачи, а в А /В лишь одна). И что на выходе? Повышение четкости сигнала по принципу прямого торможения.
А вот еще один путь для этого, но уже за счет выделения сигнала из случайного шума, создаваемого соседними нейронами, т. е. речь идет о повышении четкости сигнала в пространстве. Перед нами сеть из шести нейронов, в которой нейрон А возбуждает В, С возбуждает D , а E возбуждает F .
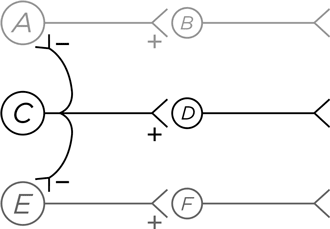
Итак, С посылает возбуждающий сигнал к D . Но вдобавок к этому его тормозящие аксоны дотягиваются до А и Е [1540]. Следовательно, если нейрон С активирован, то он одновременно возбуждает D и тормозит А и Е . С помощью такого – латерального – торможения С кричит во все горло, а А и Е молчат, закрыв рот на дополнительный замок. В этом суть пространственной очистки сигнала (не будем забывать, что диаграмма упрощенная, я на ней не нарисовал очевидных элементов – нейроны А и Е тоже посылают вбок тормозные аксоны и к С , и другим соседним нейронам в нашей воображаемой схеме).
Латеральное торможение происходит везде и всюду в сенсорных системах. Посветите в глаз тонюсеньким лучиком света. Что возбудилось – фоторецептор А, С или Е ? Благодаря латеральному торможению проще понять, что только С . И то же самое с осязанием, в результате чего можно ощутить точное место прикосновения, а не размытое «вроде бы здесь». Или ушами вы слышите, что звучит именно ля, а не ля-диез или ля-бемоль[1541].
Таким образом, мы увидели другой прием нервной системы, помогающий увеличивать контрасты. Становится понятным смысл того, что в молчащем нейроне заряд отрицательный, а не нейтральный, равный 0 мВ. Это путь к вычленению, очистке сигналов внутри одного нейрона. Обратные связи, прямое торможение, латеральное торможение… Это все способы повышения четкости сигнала в нейронной сети.
Два разных типа боли
Следующая схема включает некоторые знакомые нам теперь элементы и дает примерное представление о двух разных типах боли. Мне эта схема очень импонирует своей изящной простотой.
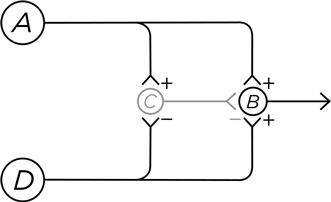
Дендриты нейрона А сидят прямо у самой поверхности кожи, и потенциал действия возникает в этом нейроне в ответ на болевое воздействие. Возбуждение от нейрона А передается на нейрон В , посылающий свои аксоны в спинной мозг, откуда вы и узнаете, что вам больно. Но нейрон А также сигнализирует нейрону С , который тормозит В . Это прямое торможение, мы уже его знаем. В итоге нейрон В ненадолго возбуждается, а затем тормозится – так происходит, когда вы переживаете момент острой боли, например, укол иголкой.
Подключим к схеме нейрон D , чьи дендриты находятся на том же участке кожи и отвечают на другой тип болевой стимуляции. Подобно соседу А , нейрон D возбуждает В , откуда сигнал идет в головной мозг. Но вместе с этим D дотягивается до С и тормозит его. И что получается? Когда нейрон D возбуждается от болевого стимула, он одновременно запрещает нейрону С тормозить нейрон В . В результате ощущается продолжительная пульсирующая боль, как при ожоге или натертости. При этом важно, что данное ощущение усиливается, поскольку потенциалы действия идут гораздо медленнее по аксону нейрона D , чем по аксону нейрона А (тут дело в миелиновых оболочках, о которых я упоминал выше, но детали здесь не слишком существенны). Поэтому сигнал боли, передаваемый нейроном D , длится дольше, хотя начинает ощущаться позже.
Два этих класса «проводки» могут взаимодействовать, и мы часто нарочно их к этому вынуждаем. Предположим, вы испытываете продолжительную ноющую боль, например от укуса насекомого. Как можно ее заглушить? Стимулировать «быстрый» аксон. Тогда на мгновение боль усиливается, но зато за счет стимуляции нейрона С линия с нейроном D на время заглушается. И в подобных обстоятельствах мы именно так и действуем. Укушенное место зудит нестерпимо, и мы расчесываем его снова и снова, потому что при этом тягучий зуд и боль на несколько минут стихают.
Для клинической практики важно знать, как срабатывают эти механизмы боли. В частности, появляется возможность разрабатывать средства от сильных хронических болей (например, в спине). С помощью микроэлектродов, имплантированных в «быструю» нейронную схему и присоединенных к стимулятору на бедре, пациент имеет возможность включать быструю линию, выключая тем самым хроническую боль. Во многих случаях это отлично срабатывает.
Таким образом, у нас имеется «линия передач» с повышенной четкостью сигнала во времени, и в ней задействовано двойное торможение – торможение тормозного нейрона, что очень круто. Как вы думаете, почему эту схему я выделяю из всех других? А потому, что она была предсказана в 1965 г. двумя великими нейробиологами, Рональдом Мелзаком и Патриком Уоллом. Они предложили ее в качестве теоретической модели: «Никто не видел подобных схем, но мы, учитывая, как срабатывает боль, полагаем, что схема должна выглядеть примерно таким образом». Последующие исследования показали, что именно так и устроены связи данной части нервной системы.
Какой из человечков?
И последняя, совершенно гипотетическая схема. Предположим, что имеется каскад из двух слоев нейронов.
Нейрон А посылает отростки к нейронам 1, 2 и 3. А нейрон В – к 2, 3 и 4 и т. п. Теперь подумаем – наделив нейроны А, В и С воображаемыми функциями, – как такая схема может работать. Пусть нейрон А возбуждается в ответ на левого человечка, В – на того, что посередине, а С – на правого.
Что может выучить нейрон № 1? Как узнавать своего конкретного человечка. Нейрон № 5 столь же узко специализирован. А на что способен нейрон № 3? Он может научиться узнавать, как одевался джентльмен Викторианской эпохи. Это нейрон, который поможет вам опознать викторианца на картинках внизу.
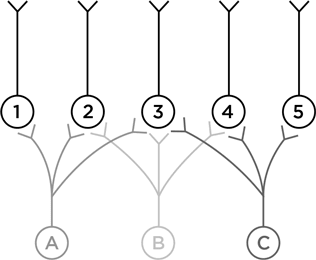
Умение нейрона № 3 неспецифичное, оно получается из перекрывания аксонов первого слоя. Нейроны № 2 и № 4 тоже неспецифичны, но их определения грубее, потому что они получают лишь по два образчика для сравнения.
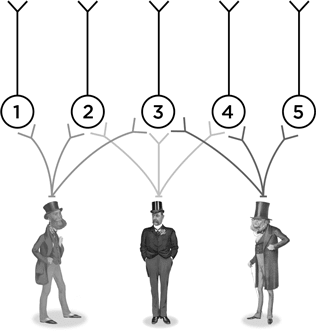
Так что нейрон № 3 располагается в точке схождения этой нейронной сети. И в самых сложно организованных участках мозга нейронные пути идут примерно так же, как на нашей воображаемой схеме, но только таких путей в тысячи тысяч раз больше. При этом нейрон № 3, вполне вероятно, окажется на периферии какой-нибудь другой схемы (к примеру, той, которую можно нарисовать перпендикулярно странице с викторианскими человечками), а нейрон № 1, напротив, играет, возможно, центральную роль в какой-то своей схеме в четвертом измерении. И так далее. Все нейроны включены в разнообразные нейронные сети.

Что это дает? Способность мыслить ассоциациями, метафорами, аналогиями, иносказаниями, символами. Связывать две несвязанные вещи, даже из разных сенсорных категорий. По-гомеровски соединять цвета моря и вина[1542]. У меня, к примеру, из-за этого тесно ассоциированы Стравинский и Пикассо, потому что на обложках альбомов Стравинского (припоминаете?) часто изображается что-нибудь из Пикассо. По той же причине прямоугольный кусок ткани, раскрашенный определенным образом, может олицетворять целый народ или идеологию.
И последнее. Мы различаемся по природе и охвату наших ассоциативных сетей. И порой маргинальные их варианты порождают интереснейшие вещи. Например, большинство из нас еще в раннем детстве научились связывать рисунок ниже с концепцией «лицо».

Но затем вступают в игру другие учителя, у которых нейронные сети проецируются шире, собственным чудны́м манером. И они показывают миру, что нечто «лицеобразное» можно увидеть и на фотографии на следующей странице.
А если спросить о последствиях подобных нетипично переплетенных сетей, то ответ будет такой – творчество.

Еще на ступеньку выше
Один нейрон, два нейрона, нейронная сеть. И мы уже готовы сделать последний шаг, подняться на уровень тысяч нейронов.
Под микроскопом срез ткани выглядит примерно так, как на картинке ниже.
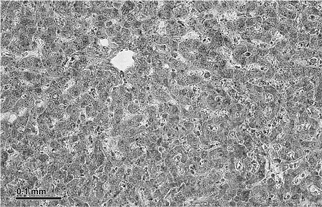
Это гомогенное поле клеток, организованных более или менее единообразно. В верхнем левом углу картинка выглядит точно так же, как и нижнем левом.
То был срез ткани печени. Один срез дает представление о всей печени. Не впечатляет.
Неужели ткань мозга столь же однообразна и скучна: такая же недифференцированная клеточная масса, заполняющая все предоставленное пространство, а нейроны рассылают свои отростки по всем возможным направлениям? Нет, у нее степень организованности фантастическая.
Другими словами, тела нейронов, занятых связанными функциями, сгруппированы вместе в строго отведенных областях мозга, а их нервные волокна, идущие к другим участкам, объединены в общий проводящий кабель. В целом это означает, что разные участки мозга заняты каждый своим особым делом . И каждый отдел мозга со своим наименованием (обычно многосложным словом на греческом или латыни) подразделяется на подотделы и подподотделы. И каждый сообщается (т. е. посылает аксоны) лишь с конкретным набором подразделений (который эти аксоны получает).
Можно свихнуться, пока выучишь все отделы и связи, и мне, к моему ужасу, встречались нейроанатомы, получавшие от этого удовольствие. Для наших же целей полезны лишь некоторые ключевые установки:
а) В любом отделе мозга содержатся миллионы нейронов. Вот некоторые уже знакомые нам наименования: гипоталамус, мозжечок, кора, гиппокамп.
б) В некоторых отделах имеются свои компактные подгруппы, которые называются «ядра». (Тут есть некоторая путаница, потому что часть клетки с ДНК тоже называется ядром. Но что поделаешь?) Вот некоторые, вероятно, совсем незнакомые вам наименования: базальное ядро Мейнерта, супраоптическое ядро гипоталамуса и еще ядро с прелестным названием – нижняя олива.
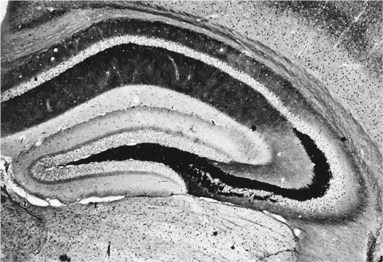
в) Как только что было сказано, тела функционально связанных нейронов собраны вместе в своих подотделах – ядрах, а кроме того, они посылают аксоны однонаправленно, в общем кабеле (в т. н. нейронные пути или нейронные тракты). Вот, например, гиппокамп.

г) Напомню, что миелиновые оболочки аксонов ускоряют перемещение вдоль них потенциала действия. Цвет миелина практически белый, поэтому нейронные тракты в мозге выглядят светлыми. Из-за этого их часто называют «белое вещество».
д) Несложно увидеть, что белое вещество и нервные волокна занимают значительную часть мозга: все его отделы посылают информацию в другие участки, зачастую весьма удаленные[1543].
е) Предположим, человек получил травму в одном из участков мозга, некоем загадочном Х . Тогда появляется возможность выяснить о данном участке нечто полезное: для этого нужно посмотреть, что у нашего человека работает неправильно. В действительности нейробиология и выросла из наблюдений за солдатами, страдающими от осколочных ранений головы. Если посмотреть на дело отстраненно, то кровавые европейские бойни XIX столетия для нейробиологов стали просто даром свыше. Получивший ранение начинает вести себя нетипичным образом. Можно ли в таком случае полагать, что участок Х в мозге отвечает за эту часть поведения? Да, но лишь когда именно в этом участке сосредоточены тела нейронов. А если в поврежденном участке имеются только нервные волокна, то, скорее всего, нужно обратить внимание на подотдел, отправляющий аксоны по этому пути, а также на тот, в который эти волокна приходят. Поэтому важно различать «нейронные ядра» и «нейронные пути».
ж) Обратимся напоследок к утверждениям, что та или иная часть мозга руководит конкретным поведением. Из примеров, приведенных мною выше, можно понять, насколько трудно определить функцию отдельного нейрона вне контекста нейронных сетей, куда он включен. И то же самое можно сказать про отдельные участки мозга. Учитывая, что каждый подотдел посылает и получает проекции из миллиона других мест, понять, кто «руководитель», удается очень и очень редко. Гораздо чаще работает вся сеть целиком, и когда говорят, что тот или иной участок «играет ключевую роль», «участвует», «опосредует», «влияет», то подразумевают именно всю сеть. Функция каждого отдельного участка мозга имеет смысл лишь в контексте его связей с другими отделами мозга.
На этом можно закончить знакомство с основами нейробиологии.
Приложение 2
Основы эндокринологии
Эндокринология – это наука о гормонах, представляющих собой, подобно нейромедиаторам из главы 2, службу передачи информации, но совершенно другую. Вспомним, что нейромедиаторы выделяются нейронными окончаниями аксонов в ответ на потенциал действия. Дальше они должны проплыть через микропространство синапса и связаться с рецепторами на дендритах второго, постсинаптического нейрона, сдвинув его заряд.
В противоположность этому гормоны являются химическими передатчиками, которые выделяются секреторными клетками (нейроны к ним тоже относятся) в различных железах. Произведенный гормон попадает в кровоток и, следовательно, может повлиять на все клетки тела, если у них найдется соответствующий рецептор[1544]. Так что уже с первых строк мы видим ключевые различия. Первое заключается в том, что нейромедиаторы напрямую влияют лишь на нейроны по ту сторону синапса, а гормоны могут повлиять на триллионы клеток тела. Второе отличие касается времени: нейромедиаторы доносят сообщение через синапс за миллисекунды; гормональный же эффект проявляется в течение часов и дней, а может продолжаться и всю жизнь (например, задумайтесь, насколько быстро проходит пубертатный период и исчезают его следы?).
Нейромедиаторы и гормоны отличаются и масштабом своих действий. Нейромедиаторы связываются с постсинаптическими рецепторами, что приводит к локальным изменениям в потоке ионов через мембрану дендритного шипика. А гормоны меняют активность конкретных белков в клетках, включают и выключают экспрессию генов, сдвигают клеточный метаболизм, заставляют клетки расти и делиться или прекратить питаться вплоть до полного истощения и смерти. Тестостерон, например, увеличивает мышечную массу, а прогестерон активирует клеточное деление в матке, за счет чего во время лютеиновой фазы ее стенки утолщаются. Еще один пример – тироидный гормон убивает клетки в хвостике головастика, когда в ходе метаморфоза тот превращается в лягушонка. А один из классов гормонов стресса уничтожает клетки иммунной системы (что объясняет, почему стресс увеличивает восприимчивость к простуде). Гормоны исключительно практичны.
Большинство гормонов является частью т. н. нейроэндокринных осей. Из главы 2 мы помним, что все дороги лимбической системы ведут в гипоталамус, который играет ключевую роль в регуляции гормональных и автономной нервной систем. Но есть еще и другая правда. Нейроны гипоталамуса выделяют специальный гормон, путешествующий по коротеньким местным путям, связывающим гипоталамус с гипофизом в самой нижней части мозга. И этот гормон, стимулируя гипофиз, заставляет его выделять свой гипофизарный гормон, который отправляется в общий кровоток и в свою очередь активирует клетки тех или иных периферийных желез плюс, соответственно, производство третьего гормона. Вот мой любимый пример с тремя гормонами. Во время стресса нейроны гипоталамуса выделяют КРГ (кортиколиберин, или кортикотропин-рилизинг-гормон), который заставляет клетки гипофиза производить АКТГ (адренокортикотропный гормон). Попав в кровь и достигнув надпочечников, АКТГ вызывает секрецию стероидных гормонов – глюкокортикоидов (у человека это кортизол или, что то же самое, гидрокортизон). Другие гормоны (эстроген, прогестерон, тестостерон, тироидный гормон) выделяются периферическими железами на конечном отрезке их собственной нейроэндокринной оси: гипоталамус – гипофиз – (какая-то периферическая) железа[1545]. А чтобы еще чудеснее усложнить эту систему, добавлю, что секрецию каждого гипофизарного гормона контролирует не только один гормон гипоталамуса. На эту работу отправляются гормоны нескольких типов, другие же при этом могут ингибировать их секрецию. Например, выделение АКТГ регулируется, помимо КРГ, еще целым рядом гормонов гипоталамуса, причем разные факторы стресса индуцируют производство разных наборов гипоталамических гормонов.
Не все гормоны регулируются по принципу оси мозг – гипофиз – эндокринная железа. В некоторых случаях срабатывает двухступенчатая последовательность мозг – гипофиз, когда гормон гипофиза напрямую влияет на все клетки тела. Таким двухступенчатым манером, например, работает гормон роста. В других случаях мозг посылает нейроны в спинной мозг и оттуда к той или иной железе внутренней секреции, участвуя в регуляции выделения ее гормона. В качестве примера можно привести поджелудочную железу и, соответственно, ее гормон инсулин (здесь главным регулятором будет уровень глюкозы в крови). А еще есть совсем странные гормоны, которые выделяются в нехарактерных местах, в сердце, скажем, или в кишечнике. В этих случаях мозг участвует в их регуляции лишь опосредованно.
Производство гормонов типа нейромедиаторов «стоит» дешево, они так специально устроены. Ведь для их синтеза полно предшественников – будь то простые белки либо холестерол[1546], – да и требуется всего несколько биохимических шагов. При этом в теле синтезируются несколько разных типов гормонов из одного предшественника. К примеру, многие стероидные гормоны построены на основе холестерола.
Теперь рассмотрим вкратце, какие бывают рецепторы гормонов. Они выполняют примерно ту же работу, что и рецепторы нейромедиаторов. Для каждого типа гормонов имеется свой рецептор[1547] со специфически вогнутым местом (сайтом) связывания, комплементарным форме молекулы гормона. Извлечем из памяти уже знакомую по нейромедиаторам метафору ключа и замка и приложим ее теперь к гормонам, она и здесь будет кстати. Как и рецепторы нейромедиаторов, рецепторы гормонов тоже не бесплатны. Ведь различные стероидные гормоны композиционно схожи. Поэтому, чтобы реагировать адекватно на их небольшие структурные различия, рецепторы должны быть подогнаны очень точно и тонко, мы ведь не хотим , чтобы они то и дело путали эстроген и тестостерон.
Вот еще одно сходство гормонов и нейромедиаторов. Подобно рецепторам нейромедиаторов, сродство рецепторов к гормонам может варьировать. Это означает, что форма сайтов связывания чуть-чуть изменяется, из-за этого гормон встраивается в сайт связывания более или менее удобно, тем самым усиливая или ослабляя эффект гормонального действия. В каждой клетке-мишени того или иного гормона число рецепторов к нему тоже не постоянно; сдвигая его в ту или другую сторону, можно повысить или понизить чувствительность клетки к гормону. Число рецепторов даже не менее важно в этом смысле, чем уровень самого гормона: есть эндокринные болезни, когда уровень гормона в норме, а рецепторы к нему мутантные, потому-то гормональный сигнал не достигает цели. Уровень гормона можно сравнить с громкостью голоса говорящего, а количество и качество рецепторов – с остротой и чуткостью слуха у собеседника.
И наконец, гормональные рецепторы, как правило, имеются только в определенных группах клеток и тканей тела, и только эти группы восприимчивы к гормону. Например, у головастиков лишь в клетках хвоста есть рецепторы к тироидному гормону, запускающему процесс клеточного отмирания. Далее, при некоторых типах рака молочной железы появляются клетки, имеющие рецепторы эстрогена, в результате они начинают расти и делиться в ответ на действие этого гормона.
Итак, мы ознакомились с тем, как гормоны влияют на функционирование клеток-мишеней; временной масштаб их влияния – это часы и дни. Особенно сильно влияние гормонов сказывается в периоды внутриутробного развития и раннего детства, что мы предметно обсуждали в главе 7. Гормоны, в частности, играют «организующую» роль при формировании мозга, т. е. определяют, как мозг «собран». А в роли «активаторов» их влияние длится не столь долго – от нескольких часов до нескольких дней. Две эти роли взаимосвязаны: результаты действия гормонов-организаторов на эмбриональный мозг создают базу для последующей работы гормонов-активаторов уже во взрослом возрасте.
Вот теперь можно вернуться обратно к основному тексту, где разобрано действие конкретных гормонов.
Приложение 3
Основные сведения о белках
Белки – это самый многочисленный класс органических молекул живых организмов. Белки исключительно важны, потому что из них построено великое множество гормонов, нейромедиаторов, молекул-посредников иммунной системы. Из белков состоят и рецепторы, отвечающие на все эти сигналы, и ферменты, необходимые для их же синтеза и разложения[1548], и специальные каркасы, служащие для придания клетке определенной формы, и т. д.
Главное свойство белка – это форма, потому что именно она предопределяет его функцию. Белки, которые строят клеточный каркас, имеют вид перекрещивающихся балок и расположены в точках клеточной сборки (ну, примерно так). Если два белковых гормона выполняют разные функции, то они должны различаться по форме. А белковый рецептор в свою очередь должен иметь вид, комплементарный молекулам своего гормона или нейромедиатора (вспомним введенную в приложении 1 знаменитую формулу «ключ к замку», т. е. посредник, например гормон, должен подходить к рецептору именно таким образом).
Некоторые белки меняют свою форму, обычно переключаясь с одной конформации на другую и обратно. Предположим, есть фермент (опять же белок), который синтезирует молекулу сахарозы, связывая вместе молекулы глюкозы и фруктозы. У этого фермента должна быть конформация, напоминающая букву V ; один ее конец связывается с глюкозой под определенным углом, а другой – с фруктозой. Когда обе молекулы оказываются в «двух руках» фермента, то он переходит в другую конформацию, в которой обе руки V сдвигаются; и тогда молекулы глюкозы и фруктозы связываются вместе. Получившаяся сахароза открепляется от фермента, и он возвращается к исходной конформации.
Что определяет форму и функции белка? Каждый белок состоит из цепочки аминокислот. В построении белков участвуют около 20 различных аминокислот, сюда входят и некоторые уже известные нам, например триптофан и глутамат. Каждая цепочка аминокислот одного белка отличается от аминокислотной последовательности любого другого белка, примерно как слова различаются последовательностями букв. В типичном белке таких аминокислотных «букв» около 300, и если учесть, что «алфавит» составлен 20 аминокислотами, то получится число возможных белков порядка 10400 (т. е. число с 400 нулями), это больше, чем атомов во Вселенной[1549]. От того, какова последовательность аминокислот, зависит, в какую форму/формы свернется белок. Принято считать, что эта последовательность определяет специфическую форму/формы белка. Однако выясняется, что на форму влияют и такие факторы, как температура и кислотность, иначе говоря, различные факторы среды.
А чем определяется последовательность аминокислот? Генами.
Дата: 2019-07-24, просмотров: 413.