Живые системы на всех уровнях организации – открытые системы. Поэтому транспорт веществ через биологические мембраны - необходимое условие жизни. С переносом веществ через мембраны связаны процессы метаболизма клетки, биоэнергетические процессы, образование биопотенциалов, генерация нервного импульса и др. Нарушение транспорта веществ через биомембраны приводит к различным патологиям. Лечение часто связано с проникновением лекарств через клеточные мембраны. Эффективность лекарственного препарата в значительной степени зависит от проницаемости для него мембраны. Проницаемость мембраны - это способность мембраны пропускать через себя атомы, ионы, молекулы веществ. В зависимости от характера связи транспорта данного иона металла или молекулы конкретного вещества от переноса других ионов или молекул веществ выделяют: унипорт - транспорт ионов или молекул через мембрану независимо от транспорта других соединений, например молекул газов, воды; симпорт - одновременный и однонаправленный перенос ионов или молекул двух различных веществ, например перенос ионов натрия и глюкозы через мембрану клеток эпителия тонкой кишки; антипорт - одновременный транспорт ионов или молекул вещества через мембрану в противоположных направлениях.
Большое значение для описания транспорта веществ имеет понятие электрохимического потенциала. Химическим потенциалом (μ) данного вещества называется величина, численно равная энергии Гиббса, приходящаяся на один моль этого вещества. Для разбавленного раствора вещества с концентрацией С:
μ = μ0 + RTlnC (3.2.1),
где μ0 - стандартный химический потенциал, численно равный химическому потенциалу данного вещества при его концентрации 1 моль/л в растворе;
R – газовая постоянная,
Т – температура.
Электрохимический потенциал μ′ - величина, численно равная энергии Гиббса G на один моль данного вещества, помещенного в электрическое поле. Для разбавленных растворов:
μ′ = μ0 + RTlnC + zFЕ (3.2.2),
где F - число Фарадея,
z - заряд иона электролита (в элементарных единицах заряда),
Е - потенциал электрического поля.
Транспорт веществ через биологические мембраны можно разделить на два основных типа: пассивный и активный.
Пассивный перенос веществ через мембрану
Пассивный транспорт - это перенос вещества из мест с большим значением электрохимического потенциала к местам с его меньшим значением. Пассивный транспорт идет с уменьшением энергии Гиббса, и поэтому этот процесс может идти самопроизвольно без затраты энергии. Плотность потока вещества jm при пассивном транспорте подчиняется уравнению Теорелла:
jm = - UC(dμ′/dx) (3.2.3),
где U - подвижность частиц,
С – концентрация вещества.
Знак минус показывает, что перенос происходит в сторону убывания μ′.
Плотность потока вещества - это величина, численно равная количеству вещества, перенесенного за единицу времени через единицу площади поверхности, перпендикулярной направлению переноса:
jm = m/(S∙t) (3.2.4),
Подставив в (3.2.3) выражение для электрохимического потенциала (3.2.2), получим для разбавленных растворов при μ0 = const уравнение Нернста—Планка:
jm = - URT(dC/dx) - UCzF(dЕ /dx) (3.2.5)
Итак, могут быть две причины переноса вещества при пассивном транспорте: градиент концентрации (dC/dx) и градиент электрического потенциала (dЕ/dx). Знаки минусов перед градиентами показывают, что градиент концентрации вызывает перенос вещества от мест с большей концентрацией к местам с его меньшей концентрацией; а градиент электрического потенциала вызывает перенос положительных зарядов от мест с большим к местам с меньшим потенциалом. В случае незаряженных веществ (z = 0) или отсутствия электрического поля (dЕ/dx) уравнение Теорелла переходит в уравнение:
jm = - URT(dC/dx) (3.2.6)
Согласно соотношению Эйнштейна, коэффициент диффузии D=URT. В результате получаем уравнение, описывающее простую диффузию - закон Фика:
jm = - D(dC/dx) (3.2.7)
На рис. 3.2.1 представлена классификация основных видов пассивного транспорта через мембрану.
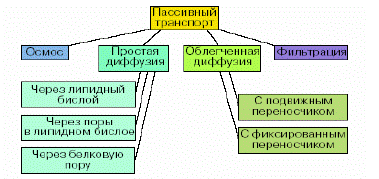
Рис. 3.2.1. Классификация видов пассивного транспорта.
Вопросы и задания для самоподготовки
1. Что такое пассивный и активный транспорт?
2. Что собой представляет унипорт и котранспорт? Приведите примеры
3. Какие вы знаете методы изучения переноса веществ? 4. Напишите уравнения для пассивного транспорта: (уравнения Теорелла, Нернста-Планка, Фика).
5. Охарактеризуйте виды пассивного транспорта (простая и облегченная диффузия, осмос, фильтрация). Приведите примеры.
6. Чем отличается облегченная диффузия от простой?
7. Что собой представляют ионные каналы? Назовите свойства ионных каналов.
8. Что такое ионофоры?
Рекомендуемая литература
1. Ремизов А.Н. Медицинская и биологическая физика : учеб. для вузов / А.Н. Ремизов, А.Г. Максина, А.Я. Потапенко. – М., 2003. – С. 191- 200.
2. Биофизика : учеб. для вузов / В.Ф. Антонов [и др.]. – М., 1999. – С. 32-42.
3. Артюхов В.Г. Биофизика : учеб. пособие / В.Г. Артюхов, Т.А. Ковалева, В.П. Шмелев. - Воронеж, 1994. – С. 198-218.
4. Биофизика : учеб. для вузов / Ю.А. Владимиров [и др.]. – М., 1983. – С. 121-129.
3.3. Активный транспорт. Ионные насосы, молекулярный механизм их работы. Сопряженный транспорт
Активный транспорт – это перенос вещества из мест с меньшим значением электрохимического потенциала в места с его большим значением. Активный транспорт в мембране сопровождается ростом энергии Гиббса, он не может идти самопроизвольно, а только за счет затраты энергии, запасенной в макроэргических связях АТФ. Активный транспорт веществ через биологические мембраны имеет огромное значение. За счет активного транспорта в организме создаются градиенты концентраций, градиенты электрических потенциалов, градиенты давления и т.д., поддерживающие жизненные процессы. Существование активного транспорта веществ через биологические мембраны впервые было доказано в опытах Уссинга (1949 г.) на примере переноса ионов натрия через кожу лягушки.
Согласно современным представлениям, в биологических мембранах имеются ионные насосы, работающие за счет энергии гидролиза АТФ, — специальные системы интегральных мембранных белков (транспортные АТФазы). В настоящее время известны три основных типа электрогенных ионных насосов, осуществляющих активный перенос ионов через мембрану: 1) при работе К+ -Nа+ -АТФазы за счет энергии, освобождающейся при гидролизе одной молекулы АТФ, в клетку переносится два иона калия и одновременно из клетки выкачиваются три иона натрия. Таким образом, создается повышенная по сравнению с межклеточной средой концентрация в клетке ионов калия и пониженная натрия, что имеет огромное физиологическое значение; 2) при работе Са2+ - АТФазы за счет энергии гидролиза АТФ переносятся два иона кальция (из клетки); 3) в Н+ -АТФазе (Н+ - помпе) происходит перенос двух протонов.
Принцип работы АТФаз-насосов основан на конформационных перестройках белковой макромолекулы при взаимодействии с транспортируемым ионом. Кальциевый насос представлен АТФазой - полипептидной цепью с молекулярной массой около 100 000 Да. Выделяют четыре этапа процесса переноса кальция (рис. 3.3.1).
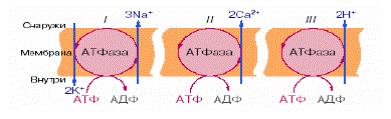
Рис. 3.3.1. Схема работы Са2+ -АТФазы
Первый этап работы Са2+ -АТФазы - связывание компонентов: иона Са2+ с комплексом Mg-АТФ. Присоединение ионов Mg2+ и Са2+ идет на наружной поверхности саркоплазматичсской мембраны к разным участкам переносчика. При связывании иона кальция освобождается энергия в 37,4 кДж/моль. Второй этап работы насоса - гидролиз АТФ с образованием фермент-фосфатного комплекса (Е~Ф). Переход комплекса Е-АТФ в Е-АДФ происходит с малым перепадом энергии. Свободная энергия гидролиза АТФ израсходовалась на синтез Е~Ф. Третий этап работы фермента – транслокация: переход центра связывания кальция на противоположную сторону мембраны. Для отрыва прочно связанных ионов кальция необходима энергия, которая доставляется за счет гидролиза АТФ. При гидролизе одного моля АТФ освобождается около 40 кДж, что достаточно для переноса двух молей ионов Са2+ через мембрану из цитоплазмы. Четвертый этап функционирования кальциевого насоса состоит в конформационных перестройках, приводящих к транслокации кальцийсвязывающих центров в исходное положение.
Вопросы и задания для самоподготовки
1. Какие вы знаете ионные насосы?
2. Опишите механизм работы кальциевой АТФазы.
3. Какие виды сопряженного транспорта вы знаете?
Рекомендуемая литература
1. Ремизов А.Н. Медицинская и биологическая физика : учеб. для вузов / А.Н. Ремизов, А.Г. Максина, А.Я. Потапенко. – М., 2003. – С. 200- 202.
2. Биофизика : учеб. для вузов / В.Ф. Антонов [и др.]. – М., 1999. – С. 42-48.
3. Артюхов В.Г. Биофизика : учеб. пособие / В.Г. Артюхов, Т.А. Ковалева, В.П. Шмелев. - Воронеж, 1994. – С. 218-223.
4. Биофизика : учеб. для вузов / Ю.А. Владимиров [и др.]. – М., 1983. – С. 129-138; 141-143.
3.4. Биоэлектрические потенциалы. Механизм формирования потенциала покоя
Одна из важнейших функций биологической мембраны – генерация и передача биопотенциалов. Это явление лежит в основе возбудимости клеток, регуляции внутриклеточных процессов, работы нервной системы, регуляции мышечного сокращения, рецепции. В медицине на исследовании электрических полей, созданных биопотенциалами органов и тканей, основаны диагностические методы: электрокардиография, электроэнцефалография, электромиография и другие. Практикуется и лечебное воздействие на ткани и органы внешними электрическими импульсами при электростимуляции. Нарушение электрических характеристик отдельных клеток, нервных волокон и целых тканей, например, сердечной ткани приводит к ряду серьезных заболеваний. Использование результатов электрофизиологических исследований в сочетании с физическим и математическим моделированием мембранных транспортных процессов лежит в основе современных теорий электрогенеза в клетках.
Различают следующие основные виды мембранных биопотенциалов: 1. Потенциал покоя – разность электрических потенциалов между внутренней и наружной поверхностью мембраны нормально функционирующей клетки в невозбужденном состоянии. 2. Потенциал действия (возбуждения) – разность потенциалов на мембране, регистрируемая в момент возбуждения между возбужденными и невозбужденными участками мембраны. 3. Потенциал повреждения – регистрируется между поврежденными и неповрежденными участками клетки, ткани, органа.
К немембранным биопотенциалам относятся метаболические потенциалы, которые регистрируются между участками с различной скоростью метаболизма внутри клетки, ткани или органа. Возникновение мембранных потенциалов связано с неравенством концентрации ионов внутри клетки и в окружающей среде и неодинаковой проницаемостью клеточной мембраны для разных ионов. Изучение механизма возникновения клеточных биопотенциалов стало возможным благодаря развитию методов клеточной электрофизиологии. В их развитии важную роль сыграли: – микроэлектродная техника; – создание усилителей биопотенциалов, обладающих высоким входным сопротивлением и высокой чувствительностью (токи до 10-12 А); – выбор удачных объектов исследования, начиная от гигантского аксона кальмара и гигантских нейронов пресноводных моллюсков и заканчивая различными модельными мембранами. Если концентрация какого-либо иона внутри клетки Сin отличается от концентрации этого иона снаружи Сout, и мембрана проницаема для этого иона, возникает поток заряженных частиц через мембрану, вследствие чего нарушается электрическая нейтральность системы, образуется разность потенциалов внутри и снаружи клетки
Ем = Еin - Еout
которая будет препятствовать дальнейшему перемещению ионов через мембрану.
Равновесие – это такое состояние системы, при котором каждая частица может переходить из некоторого состояния 1 в некоторое состояние 2 и обратно, но в целом доля состояний 1 и 2 в системе не изменяется. При установлении равновесия выравниваются значения электрохимических потенциалов по разные стороны мембраны (μ′in = μ′out). Так как μ′ = μ0 + RTlnC + zFЕ, то RTlnCin +zFЕin =RTlnCout+ zFЕout. Отсюда легко получить формулу Нернста для равновесного мембранного потенциала:
Ем = Е in - Е out = -( RT/ zF )( lnCin / Cout ) (3.4.1)
Если в формуле Нернста перейти от натурального логарифма к десятичному, то для положительного одновалентного иона (z = +1):
Ем = -2,3(RT/F)(lgCin/Cout) (3.4.2)
где Ем – равновесный потенциал, определяемый как разность потенциалов по обе стороны мембраны;
R – универсальная газовая постоянная;
T – абсолютная температура (К);
z – заряд иона;
F – постоянная Фарадея;
Cin и Cout – концентрации потенциалопределяющих ионов по обе стороны мембраны.
В 1902 г. Бернштейн выдвинул гипотезу, согласно которой потенциал покоя обусловлен тем, что цитоплазматическая мембрана проницаема для ионов К+, и на ней создается потенциал, описываемый уравнением Нернста (равновесный потенциал). Диффузия ионов через мембрану, при которой вещества транспортируются по концентрационному градиенту, создает разность электрических потенциалов. Движение ионов по концентрационному градиенту обусловлено силой, имеющей химическую природу. По мере диффузии ионов К+ из клетки ионы хлора проникают в клетку по своему концентрационному градиенту. Противоположные заряды притягиваются, поэтому электрическая сила заставляет ионы калия стремится в клетку за ионами хлора. Для более точного вычисления величины мембранного потенциала необходимо учитывать диффузию ионов К+ , Na+ и Cl-. В связи с этим для определения мембранного потенциала используют уравнение Гольдмана.
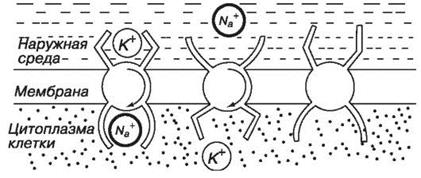
Уравнения Нернста и Гольдмана не учитывают активного транспорта ионов через мембрану, наличие в мембране ионных насосов. В цитоплазматической мембране функционируют молекулы Na+ ,К+ - АТФазы, перекачивающие калий внутрь клетки, а натрий из клетки (т.е. против концентрационного градиента). С учетом работы электрогенных ионных насосов для мембранных потенциалов используют уравнение Томаса.
Вопросы и задания для самоподготовки
1. Что такое электрический потенциал, разность потенциалов?
2. Что является причиной переноса ионов К+ , Na+ и С1- через биологическую мембрану, какими уравнениями описываются эти процессы?
3. Что такое мембранный потенциал?
4. Классификация электрических потенциалов биосистем.
5. Опишите механизм формирования потенциала покоя.
6. Какова роль ионных каналов и насосов в формировании потенциала покоя?
7. Напишите уравнения Нернста, Гольдмана, Томаса. В чем особенности этих уравнений?
Рекомендуемая литература
1. Ремизов А.Н. Медицинская и биологическая физика : учеб. для вузов / А.Н. Ремизов, А.Г. Максина, А.Я. Потапенко. – М., 2003. – С. 203- 207.
2. Биофизика : учеб. для вузов / В.Ф. Антонов [и др.]. – М., 1999. – С. 67-77.
3. Биофизика : учеб. для вузов / Ю.А. Владимиров [и др.]. – М., 1983. – С. 150-156.
4. Биофизика : практикум для студентов / В.Г. Артюхов, О.В. Башарина. – Воронеж, 2003. – С.42-48.
3.5. Потенциал действия, его свойства. Биофизика нервного импульса
Потенциалом действия (ПД) называется электрический импульс, обусловленный изменением ионной проницаемости мембраны и связанный с распространением по нервам и мышцам волны возбуждения. Опыты по исследованию потенциала действия проведены (в основном Ходжкиным и его сотрудниками) на гигантских аксонах кальмара методом микроэлектродов с использованием высокоомных измерителей напряжения, а также методом меченых атомов. Возбуждающий импульс вызывает лишь на короткое время смещение мембранного потенциала, который быстро пропадает и восстанавливается потенциал покоя. В том случае, когда возбуждающий импульс смещается еще дальше в отрицательную сторону, он сопровождается гиперполяризацией мембраны. Потенциал действия также не формируется, когда возбуждающий импульс положительный (деполяризующий), но его амплитуда меньше порогового значения. Однако если амплитуда положительного деполяризующего импульса окажется больше поргового значения в мембране развивается процесс, в результате которого происходит резкое повышение мембранного потенциала и мембранный потенциал φм меняет свой знак — становится положительным. Достигнув некоторого положительного значения потенциала реверсии, мембранный потенциал возвращается к значению потенциала покоя, совершив нечто вроде затухающего колебания. В нервных волокнах и скелетных мышцах длительность потенциала действия около 1 мс (а в сердечной мышце около 300 мс). После снятия возбуждения еще в течение 1 - 3 мс в мембране наблюдаются некоторые остаточные явления, во время которых мембрана рефрактерна (невозбудима). Новый деполяризующий потенциал может вызвать образование нового потенциала действия только после полного возвращения мембраны в состояние покоя. Причем амплитуда потенциала действия не зависит от амплитуды деполяризующего потенциала. Характерные свойства потенциала действия: 1) наличие порогового значения деполяризующего потенциала; 2) закон "все или ничего", то есть, если деполяризующий потенциал больше порогового, развивается потенциал действия, амплитуда которого не зависит от амплитуды возбуждающего импульса и нет потенциала действия, если амплитуда деполяризующего потенциала меньше пороговой; 3) есть период рефрактерности, невозбудимости мембраны во время развития потенциала действия и остаточных явлений после снятия возбуждения; 4) в момент возбуждения резко уменьшается сопротивление мембраны.
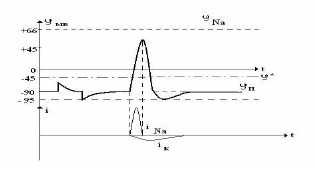
При развитии потенциала действия наряду с изменением проницаемости происходит кратковременное увеличение электропроводности мембраны. В первой фазе ПД - фазе деполяризации - усиленный поток ионов Nа+ , направленный внутрь клетки, уравновешивает концентрационный градиент, и поступление в клетку натрия прекращается. Внутренняя поверхность мембраны заряжается положительно по отношению к наружной. В это время проницаемость мембраны для Na+ увеличивается в 500 раз за 0,5 - 1 мс. Затем возрастает проницаемость мембраны для ионов К+, и усиливается диффузия этих ионов из клетки. В результате происходит уменьшение мембранного потенциала, что, в свою очередь, снижает проницаемость мембраны для Nа+. Это продолжается до тех пор, пока потенциал покоя не восстановится. После этого проницаемость для ионов К+ падает до исходного уровня. Фаза, в течение которой мембранный потенциал возвращается к уровню потенциала покоя, называется фазой реполяризации. Она осуществляется не в результате обратного перемещения ионов Nа+, а вследствие выхода из клетки эквивалентного количества ионов К+. Фаза реполяризации всегда продолжительнее фазы деполяризации. Следовательно, формирование ПД обусловлено двумя ионными потоками через биомембрану, которые приблизительно равны по величине, но сдвинуты во времени. Возбуждение мембраны описывается уравнением Ходжкина-Хаксли:
Iм = Cм(dφм/dt) + ∑Ii (3.5.1)
где Iм - ток через мембрану,
См - емкость мембраны,
∑Ii - сумма ионных токов через мембрану.
Электрический ток через мембрану складывается из ионных токов: ионов калия - IК, натрия - INa и других ионов, в том числе С1- , так называемого тока утечки Iут , а также емкостного тока. Емкостной ток обусловлен перезарядкой конденсатора, который представляет собой мембрана, перетеканием зарядов с одной ее поверхности на другую.
Возбуждение по миелинизированному волокну распространяется скачкообразно от одного перехвата Ранвье (участка, свободного от миелиновой оболочки) до другого. Нервные импульсы проводятся по аксонам в какой-то степени аналогично тому, как передаются электрические сигналы по кабельно-релейной линии. Электрический импульс передается без затухания за счет его усиления на промежуточных релейных станциях, роль которых в аксонах выполняют участки возбудимой мембраны, в которых генерируются потенциалы действия.
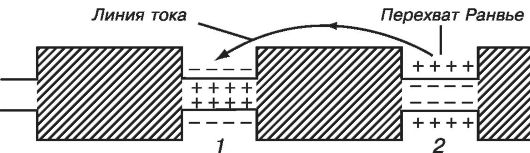
Вопросы и задания для самоподготовки
1. Что такое потенциал действия?
2. Какие фазы выделяют в потенциале действия?
3. Как изменяется проницаемость мембраны для ионов в этих фазах?
4. Опишите схему работы натриевого канала.
5. Напишите уравнение Ходжкина-Хаксли.
6. Как происходит распространение возбуждения вдоль нервного и мышечного волокна?
Рекомендуемая литература
1. Ремизов А.Н. Медицинская и биологическая физика : учеб. для вузов / А.Н. Ремизов, А.Г. Максина, А.Я. Потапенко. – М., 2003. – С. 207- 213.
2. Биофизика : учеб. для вузов / В.Ф. Антонов [и др.]. – М., 1999. – С. 77-111.
3. Биофизика : учеб. для вузов / Ю.А. Владимиров [и др.]. – М., 1983. – С. 156-161; 167-172.
Дата: 2019-03-05, просмотров: 378.