Министерство сельского хозяйства Российской Федерации
федеральное государственное бюджетное образовательное
учреждение высшего образования
«Уральский государственный аграрный университет»
УЧЕБНО-МЕТОДИЧЕСКОЕ ПОСОБИЕ ДЛЯ САМОСТОЯТЕЛЬНОЙ ПОДГОТОВКИ СТУДЕНТОВ К КУРСУ «БИОФИЗИКА»
Составитель: Кузнецова Е.И.
Екатеринбург 2017
ВВЕДЕНИЕ
Проблема совершенствования самостоятельной работы студентов имеет исключительно большое значение в подготовке высококвалифицированных, творчески активных специалистов. Под самостоятельной работой следует понимать все виды активной познавательной и научной деятельности, требующие поисков ответов на вопросы, возникающие в ходе изучения учебного материала или проведения научных исследований.
Самостоятельная работа студентов преследует следующие цели: 1) закрепление знаний, умений и навыков студентов, их расширение и углубление; 2) формирование умения применять полученные знания в конкретных жизненных условиях, практических ситуациях; 3) выработка собственных способов, приемов, методов самостоятельного мышления, творческой активности, умения вести научный поиск.
В ходе приобщения студентов к самостоятельной работе возрастает значение внеаудиторных форм работы при непосредственном руководстве преподавателя. Специфика внеаудиторной работы студентов предусматривает обязательную работу с учебной литературой, углубление знаний в процессе выполнения заданий и самостоятельного поиска, творческий отбор материала, задач, их моделирование, овладение научной терминологией и культурой речи. Самостоятельная работа относится к наиболее эффективным средствам развития познавательной активности студентов и способствует формированию у них самостоятельности в процессе обучения в вузе.
На ветеринарном факультете в 1 семестре учебный план предусматривает организацию лекций (20 часов) и лабораторных занятий (36 часов) по биофизике. При проведении практикума по биофизике оценка знаний студентов осуществляется при сдаче работ и решении задач на данную тему. Ниже приведены разделы биофизики, изучение которых входит в учебный план. Для облегчения подготовки студента к данным темам приводятся вопросы, на которые студент должен ответить в ходе изучения материала, и список литературы, где данный материал излагается. При подготовке необходимо использовать также конспекты лекций.
Тема 1. Биофизика как наука
Рис. 3.1.1. Жидкостно - мозаичная модель строения биологических мембран.
Различают поверхностные (или периферические), полупогруженные и интегральные белки. Липиды находятся при физиологических условиях в жидком агрегатном состоянии. Это позволяет сравнить мембрану с фосфолипидным морем, по которому плавают белковые "айсберги". Полярные головы молекул фосфолипидов - гидрофильны, а их неполярные хвосты - гидрофобны. В смеси фосфолипидов с водой термодинамически выгодно, чтобы полярные головы были погружены в состоящую из полярных молекул воду, а их неполярные хвосты были бы расположены подальше от воды. Такое расположение амфифильных (имеющих и гидрофильную, и гидрофобную части) молекул соответствует наименьшему значению энергии по сравнению с другими возможными расположениями молекул. Кроме фосфолипидов и белков, в биологических мембранах содержатся и другие химические соединения. В мембранах животных клеток много холестерина. Есть в мембранах и другие вещества, например гликолипиды, гликопротеиды.
Мембрана – динамическое образование, так как и белки, и липиды способны перемещаться в составе мембраны. Различают латеральную диффузию (перемещение в одном слое) и трансбислойный (флип-флоп) переход (перескок молекулы из одного слоя мембраны в другой).
Функционирование мембраны сильно зависит от микровязкости липидного бислоя и подвижности фосфолипидных молекул в мембране, фазового состояния мембранных липидов. Отклонения биофизических характеристик липидного бислоя от нормы связано с разного рода патологиями. Так, например, при воспалении в результате пероксидного окисления ненасыщенных жирных кислот в составе фосфолипидов вязкость мембраны увеличивается, а это приводит к нарушению функционирования клетки в целом. В основе многих патологических состояний организма человека лежат изменения структурно-функциональных свойств молекулярных компонентов биомембран, которые происходят при воздействии внешних факторов среды (фармакологические агенты, яды, токсины, аллергены, ионизирующее и УФ-излучение и др.) или при внутренних функциональных расстройствах. К заболеваниям подобного рода следует отнести гипертонию, атеросклероз, ишемию, бронхолегочные заболевания, различные воспаления, злокачественный рост клеток. В связи с этим всесторонние исследования механизмов функционирования биомембран в норме и при патологии необходимы как для разработки методов лечения и профилактики вышеназванных заболеваний, так и для создания высокоэффективных лекарственных препаратов.
Модельные липидные мембраны
Липосомы, или фосфолипидные везикулы (пузырьки), получают обычно при набухании сухих фосфолипидов в воде или при впрыскивании раствора липидов в воду. При этом происходит самосборка бимолекулярной липидной мембраны. Минимуму энергии Гиббса отвечает замкнутая сферическая одноламеллярная форма мембраны. При этом все неполярные гидрофобные хвосты находятся внутри мембраны и ни один из них не соприкасается с полярными молекулами воды (рис. 3.1.2.).
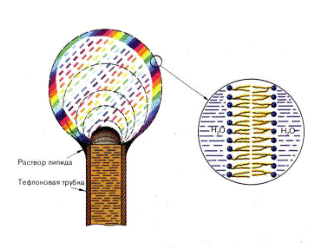
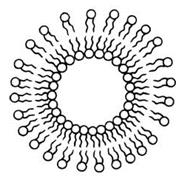
Рис. 3.1.2. Схема строения однослойной липосомы.
Однако чаще получаются несферические многоламеллярные липосомы, состоящие из нескольких бимолекулярных слоев, - многослойные липосомы. Отдельные бимолекулярные слои многослойной липосомы отделены водной средой. Диаметр многослойных липосом колеблется в пределах от 60 до 400 нм и более. Однослойные липосомы можно получить различными методами, например из суспензии многослойных липосом, если обработать их ультразвуком. Диаметр однослойных липосом, полученных этим методом, составляет 25 - 30 нм. Липосомы служат моделью для исследований различных свойств клеточных мембран. Липосомы нашли непосредственное применение в изготовлении лекарств. Например, можно заключить внутрь липосом лекарственный препарат и использовать как фосфолипидную микрокапсулу для доставки лекарства в определенные органы и ткани. Липосомы не токсичны (при правильном подборе липидов), полностью усваиваются организмом, способны преодолевать некоторые биологические барьеры. Так, инсулин, заключенный в липосому, защищен от действия пищеварительных ферментов. В настоящее время выясняется возможность вводить этот препарат в липосомах перорально, что может избавить больных диабетом от необходимости систематических уколов. Проводятся работы по разработке методов липосомальной терапии опухолей, ферментативной недостаточности, атеросклероза. Изучается возможность прицельной доставки лекарственного препарата, заключенного в липосомах, к больному органу или даже к больному участку (в частности, к пораженному участку сердца). Для этого к липосоме присоединяется белковая молекула - антитело к соответствующему мембранному антигену органа-мишени. Несмотря на заманчивые перспективы липосомальной терапии, еще имеется достаточно много нерешенных вопросов.
Вопросы и задания для самоподготовки
1. Что собой представляет мозаичная жидкокристаллическая модель биомембраны?
2. Мембранные белки, их структура, свойства и функции.
3. Какие вы знаете типы жидкокристаллических структур? Какой тип жидкокристаллической структуры имеет биомембрана?
4. Охарактеризуйте динамику структурных элементов биомембраны: латеральную диффузию и трансмембранные переходы («флип-флоп» переходы).
5. Какие вы знаете примеры модельных липидных мембран?
6. Что собой представляют мицеллы и липосомы (везикулы)?
Рекомендуемая литература
1. Ремизов А.Н. Медицинская и биологическая физика : учеб. для вузов / А.Н. Ремизов, А.Г. Максина, А.Я. Потапенко. – М., 2003. – С. 184- 190.
2. Биофизика : учеб. для вузов / В.Ф. Антонов [и др.]. – М., 1999. – С. 8-31.
3. Артюхов В.Г. Биологические мембраны: структурная организация, функции, модификации физико-химическими агентами : учеб. пособие / В.Г. Артюхов, М.А. Наквасина. - Воронеж, 1994. – С. 11-62.
Рис. 3.2.1. Классификация видов пассивного транспорта.
Вопросы и задания для самоподготовки
1. Что такое пассивный и активный транспорт?
2. Что собой представляет унипорт и котранспорт? Приведите примеры
3. Какие вы знаете методы изучения переноса веществ? 4. Напишите уравнения для пассивного транспорта: (уравнения Теорелла, Нернста-Планка, Фика).
5. Охарактеризуйте виды пассивного транспорта (простая и облегченная диффузия, осмос, фильтрация). Приведите примеры.
6. Чем отличается облегченная диффузия от простой?
7. Что собой представляют ионные каналы? Назовите свойства ионных каналов.
8. Что такое ионофоры?
Рекомендуемая литература
1. Ремизов А.Н. Медицинская и биологическая физика : учеб. для вузов / А.Н. Ремизов, А.Г. Максина, А.Я. Потапенко. – М., 2003. – С. 191- 200.
2. Биофизика : учеб. для вузов / В.Ф. Антонов [и др.]. – М., 1999. – С. 32-42.
3. Артюхов В.Г. Биофизика : учеб. пособие / В.Г. Артюхов, Т.А. Ковалева, В.П. Шмелев. - Воронеж, 1994. – С. 198-218.
4. Биофизика : учеб. для вузов / Ю.А. Владимиров [и др.]. – М., 1983. – С. 121-129.
3.3. Активный транспорт. Ионные насосы, молекулярный механизм их работы. Сопряженный транспорт
Активный транспорт – это перенос вещества из мест с меньшим значением электрохимического потенциала в места с его большим значением. Активный транспорт в мембране сопровождается ростом энергии Гиббса, он не может идти самопроизвольно, а только за счет затраты энергии, запасенной в макроэргических связях АТФ. Активный транспорт веществ через биологические мембраны имеет огромное значение. За счет активного транспорта в организме создаются градиенты концентраций, градиенты электрических потенциалов, градиенты давления и т.д., поддерживающие жизненные процессы. Существование активного транспорта веществ через биологические мембраны впервые было доказано в опытах Уссинга (1949 г.) на примере переноса ионов натрия через кожу лягушки.
Согласно современным представлениям, в биологических мембранах имеются ионные насосы, работающие за счет энергии гидролиза АТФ, — специальные системы интегральных мембранных белков (транспортные АТФазы). В настоящее время известны три основных типа электрогенных ионных насосов, осуществляющих активный перенос ионов через мембрану: 1) при работе К+ -Nа+ -АТФазы за счет энергии, освобождающейся при гидролизе одной молекулы АТФ, в клетку переносится два иона калия и одновременно из клетки выкачиваются три иона натрия. Таким образом, создается повышенная по сравнению с межклеточной средой концентрация в клетке ионов калия и пониженная натрия, что имеет огромное физиологическое значение; 2) при работе Са2+ - АТФазы за счет энергии гидролиза АТФ переносятся два иона кальция (из клетки); 3) в Н+ -АТФазе (Н+ - помпе) происходит перенос двух протонов.
Принцип работы АТФаз-насосов основан на конформационных перестройках белковой макромолекулы при взаимодействии с транспортируемым ионом. Кальциевый насос представлен АТФазой - полипептидной цепью с молекулярной массой около 100 000 Да. Выделяют четыре этапа процесса переноса кальция (рис. 3.3.1).
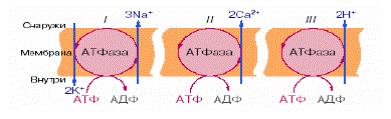
Рис. 3.3.1. Схема работы Са2+ -АТФазы
Первый этап работы Са2+ -АТФазы - связывание компонентов: иона Са2+ с комплексом Mg-АТФ. Присоединение ионов Mg2+ и Са2+ идет на наружной поверхности саркоплазматичсской мембраны к разным участкам переносчика. При связывании иона кальция освобождается энергия в 37,4 кДж/моль. Второй этап работы насоса - гидролиз АТФ с образованием фермент-фосфатного комплекса (Е~Ф). Переход комплекса Е-АТФ в Е-АДФ происходит с малым перепадом энергии. Свободная энергия гидролиза АТФ израсходовалась на синтез Е~Ф. Третий этап работы фермента – транслокация: переход центра связывания кальция на противоположную сторону мембраны. Для отрыва прочно связанных ионов кальция необходима энергия, которая доставляется за счет гидролиза АТФ. При гидролизе одного моля АТФ освобождается около 40 кДж, что достаточно для переноса двух молей ионов Са2+ через мембрану из цитоплазмы. Четвертый этап функционирования кальциевого насоса состоит в конформационных перестройках, приводящих к транслокации кальцийсвязывающих центров в исходное положение.
Вопросы и задания для самоподготовки
1. Какие вы знаете ионные насосы?
2. Опишите механизм работы кальциевой АТФазы.
3. Какие виды сопряженного транспорта вы знаете?
Рекомендуемая литература
1. Ремизов А.Н. Медицинская и биологическая физика : учеб. для вузов / А.Н. Ремизов, А.Г. Максина, А.Я. Потапенко. – М., 2003. – С. 200- 202.
2. Биофизика : учеб. для вузов / В.Ф. Антонов [и др.]. – М., 1999. – С. 42-48.
3. Артюхов В.Г. Биофизика : учеб. пособие / В.Г. Артюхов, Т.А. Ковалева, В.П. Шмелев. - Воронеж, 1994. – С. 218-223.
4. Биофизика : учеб. для вузов / Ю.А. Владимиров [и др.]. – М., 1983. – С. 129-138; 141-143.
3.4. Биоэлектрические потенциалы. Механизм формирования потенциала покоя
Одна из важнейших функций биологической мембраны – генерация и передача биопотенциалов. Это явление лежит в основе возбудимости клеток, регуляции внутриклеточных процессов, работы нервной системы, регуляции мышечного сокращения, рецепции. В медицине на исследовании электрических полей, созданных биопотенциалами органов и тканей, основаны диагностические методы: электрокардиография, электроэнцефалография, электромиография и другие. Практикуется и лечебное воздействие на ткани и органы внешними электрическими импульсами при электростимуляции. Нарушение электрических характеристик отдельных клеток, нервных волокон и целых тканей, например, сердечной ткани приводит к ряду серьезных заболеваний. Использование результатов электрофизиологических исследований в сочетании с физическим и математическим моделированием мембранных транспортных процессов лежит в основе современных теорий электрогенеза в клетках.
Различают следующие основные виды мембранных биопотенциалов: 1. Потенциал покоя – разность электрических потенциалов между внутренней и наружной поверхностью мембраны нормально функционирующей клетки в невозбужденном состоянии. 2. Потенциал действия (возбуждения) – разность потенциалов на мембране, регистрируемая в момент возбуждения между возбужденными и невозбужденными участками мембраны. 3. Потенциал повреждения – регистрируется между поврежденными и неповрежденными участками клетки, ткани, органа.
К немембранным биопотенциалам относятся метаболические потенциалы, которые регистрируются между участками с различной скоростью метаболизма внутри клетки, ткани или органа. Возникновение мембранных потенциалов связано с неравенством концентрации ионов внутри клетки и в окружающей среде и неодинаковой проницаемостью клеточной мембраны для разных ионов. Изучение механизма возникновения клеточных биопотенциалов стало возможным благодаря развитию методов клеточной электрофизиологии. В их развитии важную роль сыграли: – микроэлектродная техника; – создание усилителей биопотенциалов, обладающих высоким входным сопротивлением и высокой чувствительностью (токи до 10-12 А); – выбор удачных объектов исследования, начиная от гигантского аксона кальмара и гигантских нейронов пресноводных моллюсков и заканчивая различными модельными мембранами. Если концентрация какого-либо иона внутри клетки Сin отличается от концентрации этого иона снаружи Сout, и мембрана проницаема для этого иона, возникает поток заряженных частиц через мембрану, вследствие чего нарушается электрическая нейтральность системы, образуется разность потенциалов внутри и снаружи клетки
Ем = Еin - Еout
которая будет препятствовать дальнейшему перемещению ионов через мембрану.
Равновесие – это такое состояние системы, при котором каждая частица может переходить из некоторого состояния 1 в некоторое состояние 2 и обратно, но в целом доля состояний 1 и 2 в системе не изменяется. При установлении равновесия выравниваются значения электрохимических потенциалов по разные стороны мембраны (μ′in = μ′out). Так как μ′ = μ0 + RTlnC + zFЕ, то RTlnCin +zFЕin =RTlnCout+ zFЕout. Отсюда легко получить формулу Нернста для равновесного мембранного потенциала:
Ем = Е in - Е out = -( RT/ zF )( lnCin / Cout ) (3.4.1)
Если в формуле Нернста перейти от натурального логарифма к десятичному, то для положительного одновалентного иона (z = +1):
Ем = -2,3(RT/F)(lgCin/Cout) (3.4.2)
где Ем – равновесный потенциал, определяемый как разность потенциалов по обе стороны мембраны;
R – универсальная газовая постоянная;
T – абсолютная температура (К);
z – заряд иона;
F – постоянная Фарадея;
Cin и Cout – концентрации потенциалопределяющих ионов по обе стороны мембраны.
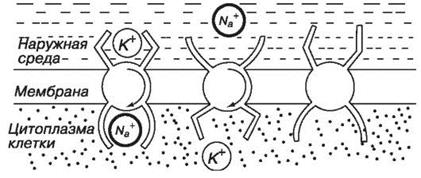
В 1902 г. Бернштейн выдвинул гипотезу, согласно которой потенциал покоя обусловлен тем, что цитоплазматическая мембрана проницаема для ионов К+, и на ней создается потенциал, описываемый уравнением Нернста (равновесный потенциал). Диффузия ионов через мембрану, при которой вещества транспортируются по концентрационному градиенту, создает разность электрических потенциалов. Движение ионов по концентрационному градиенту обусловлено силой, имеющей химическую природу. По мере диффузии ионов К+ из клетки ионы хлора проникают в клетку по своему концентрационному градиенту. Противоположные заряды притягиваются, поэтому электрическая сила заставляет ионы калия стремится в клетку за ионами хлора. Для более точного вычисления величины мембранного потенциала необходимо учитывать диффузию ионов К+ , Na+ и Cl-. В связи с этим для определения мембранного потенциала используют уравнение Гольдмана.
Уравнения Нернста и Гольдмана не учитывают активного транспорта ионов через мембрану, наличие в мембране ионных насосов. В цитоплазматической мембране функционируют молекулы Na+ ,К+ - АТФазы, перекачивающие калий внутрь клетки, а натрий из клетки (т.е. против концентрационного градиента). С учетом работы электрогенных ионных насосов для мембранных потенциалов используют уравнение Томаса.
Вопросы и задания для самоподготовки
1. Что такое электрический потенциал, разность потенциалов?
2. Что является причиной переноса ионов К+ , Na+ и С1- через биологическую мембрану, какими уравнениями описываются эти процессы?
3. Что такое мембранный потенциал?
4. Классификация электрических потенциалов биосистем.
5. Опишите механизм формирования потенциала покоя.
6. Какова роль ионных каналов и насосов в формировании потенциала покоя?
7. Напишите уравнения Нернста, Гольдмана, Томаса. В чем особенности этих уравнений?
Рекомендуемая литература
1. Ремизов А.Н. Медицинская и биологическая физика : учеб. для вузов / А.Н. Ремизов, А.Г. Максина, А.Я. Потапенко. – М., 2003. – С. 203- 207.
2. Биофизика : учеб. для вузов / В.Ф. Антонов [и др.]. – М., 1999. – С. 67-77.
3. Биофизика : учеб. для вузов / Ю.А. Владимиров [и др.]. – М., 1983. – С. 150-156.
4. Биофизика : практикум для студентов / В.Г. Артюхов, О.В. Башарина. – Воронеж, 2003. – С.42-48.
3.5. Потенциал действия, его свойства. Биофизика нервного импульса
Потенциалом действия (ПД) называется электрический импульс, обусловленный изменением ионной проницаемости мембраны и связанный с распространением по нервам и мышцам волны возбуждения. Опыты по исследованию потенциала действия проведены (в основном Ходжкиным и его сотрудниками) на гигантских аксонах кальмара методом микроэлектродов с использованием высокоомных измерителей напряжения, а также методом меченых атомов. Возбуждающий импульс вызывает лишь на короткое время смещение мембранного потенциала, который быстро пропадает и восстанавливается потенциал покоя. В том случае, когда возбуждающий импульс смещается еще дальше в отрицательную сторону, он сопровождается гиперполяризацией мембраны. Потенциал действия также не формируется, когда возбуждающий импульс положительный (деполяризующий), но его амплитуда меньше порогового значения. Однако если амплитуда положительного деполяризующего импульса окажется больше поргового значения в мембране развивается процесс, в результате которого происходит резкое повышение мембранного потенциала и мембранный потенциал φм меняет свой знак — становится положительным. Достигнув некоторого положительного значения потенциала реверсии, мембранный потенциал возвращается к значению потенциала покоя, совершив нечто вроде затухающего колебания. В нервных волокнах и скелетных мышцах длительность потенциала действия около 1 мс (а в сердечной мышце около 300 мс). После снятия возбуждения еще в течение 1 - 3 мс в мембране наблюдаются некоторые остаточные явления, во время которых мембрана рефрактерна (невозбудима). Новый деполяризующий потенциал может вызвать образование нового потенциала действия только после полного возвращения мембраны в состояние покоя. Причем амплитуда потенциала действия не зависит от амплитуды деполяризующего потенциала. Характерные свойства потенциала действия: 1) наличие порогового значения деполяризующего потенциала; 2) закон "все или ничего", то есть, если деполяризующий потенциал больше порогового, развивается потенциал действия, амплитуда которого не зависит от амплитуды возбуждающего импульса и нет потенциала действия, если амплитуда деполяризующего потенциала меньше пороговой; 3) есть период рефрактерности, невозбудимости мембраны во время развития потенциала действия и остаточных явлений после снятия возбуждения; 4) в момент возбуждения резко уменьшается сопротивление мембраны.
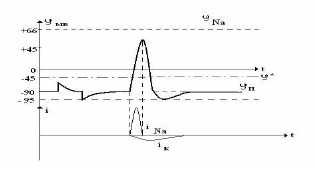
При развитии потенциала действия наряду с изменением проницаемости происходит кратковременное увеличение электропроводности мембраны. В первой фазе ПД - фазе деполяризации - усиленный поток ионов Nа+ , направленный внутрь клетки, уравновешивает концентрационный градиент, и поступление в клетку натрия прекращается. Внутренняя поверхность мембраны заряжается положительно по отношению к наружной. В это время проницаемость мембраны для Na+ увеличивается в 500 раз за 0,5 - 1 мс. Затем возрастает проницаемость мембраны для ионов К+, и усиливается диффузия этих ионов из клетки. В результате происходит уменьшение мембранного потенциала, что, в свою очередь, снижает проницаемость мембраны для Nа+. Это продолжается до тех пор, пока потенциал покоя не восстановится. После этого проницаемость для ионов К+ падает до исходного уровня. Фаза, в течение которой мембранный потенциал возвращается к уровню потенциала покоя, называется фазой реполяризации. Она осуществляется не в результате обратного перемещения ионов Nа+, а вследствие выхода из клетки эквивалентного количества ионов К+. Фаза реполяризации всегда продолжительнее фазы деполяризации. Следовательно, формирование ПД обусловлено двумя ионными потоками через биомембрану, которые приблизительно равны по величине, но сдвинуты во времени. Возбуждение мембраны описывается уравнением Ходжкина-Хаксли:
Iм = Cм(dφм/dt) + ∑Ii (3.5.1)
где Iм - ток через мембрану,
См - емкость мембраны,
∑Ii - сумма ионных токов через мембрану.
Электрический ток через мембрану складывается из ионных токов: ионов калия - IК, натрия - INa и других ионов, в том числе С1- , так называемого тока утечки Iут , а также емкостного тока. Емкостной ток обусловлен перезарядкой конденсатора, который представляет собой мембрана, перетеканием зарядов с одной ее поверхности на другую.
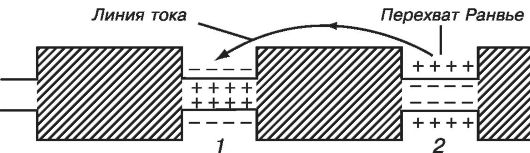
Возбуждение по миелинизированному волокну распространяется скачкообразно от одного перехвата Ранвье (участка, свободного от миелиновой оболочки) до другого. Нервные импульсы проводятся по аксонам в какой-то степени аналогично тому, как передаются электрические сигналы по кабельно-релейной линии. Электрический импульс передается без затухания за счет его усиления на промежуточных релейных станциях, роль которых в аксонах выполняют участки возбудимой мембраны, в которых генерируются потенциалы действия.
Вопросы и задания для самоподготовки
1. Что такое потенциал действия?
2. Какие фазы выделяют в потенциале действия?
3. Как изменяется проницаемость мембраны для ионов в этих фазах?
4. Опишите схему работы натриевого канала.
5. Напишите уравнение Ходжкина-Хаксли.
6. Как происходит распространение возбуждения вдоль нервного и мышечного волокна?
Рекомендуемая литература
1. Ремизов А.Н. Медицинская и биологическая физика : учеб. для вузов / А.Н. Ремизов, А.Г. Максина, А.Я. Потапенко. – М., 2003. – С. 207- 213.
2. Биофизика : учеб. для вузов / В.Ф. Антонов [и др.]. – М., 1999. – С. 77-111.
3. Биофизика : учеб. для вузов / Ю.А. Владимиров [и др.]. – М., 1983. – С. 156-161; 167-172.
Первый закон термодинамики
Это закон сохранения энергии, сформулированный в 1842-1847 г.г. Ю.Р. Майером и Г.А. Гельмгольцем: в изолированной термодинамической системе полный запас энергии есть величина постоянная и возможны только превращения одного вида энергии в другой в эквивалентных соотношениях:
U = const; ΔU = 0 (6.1.2)
Формулировка первого начала (закона) термодинамики для закрытых систем следующая: теплота, подведенная к закрытой системе, расходуется на изменение внутренней энергии системы и на совершение работы против внешних сил:
δQ = dU + δA (6.1.3)
где Q - теплота, подведенная к системе;
U - внутренняя энергия системы;
А – работа;
δ обозначает, что теплота и работа не являются функциями состояния системы и не могут быть полными дифференциалами.
В настоящее время применение закона сохранения энергии к биологическим объектам не вызывает сомнения.
Закон Гесса - основное следствие первого начала термодинамики. Он формулируется следующим образом: тепловой эффект химической реакции, развивающейся через ряд промежуточных стадий, не зависит от пути перехода, а определяется лишь разностью энтальпий конечных и исходных продуктов реакции.
Второе начало термодинамики позволяет судить о направлении протекания процесса и, таким образом, дополняет первое начало термодинамики. Формулировку второго начала термодинамики впервые дали независимо друг от друга Р. Клаузиус и У. Томпсон (1850-1851). Второе начало не имеет столь простой и общепринятой формулировки, как первое, и формулируется по-разному, в зависимости от той группы явлений, к которой оно прилагается. С. Карно: ―Коэффициент полезного действия всех обратимых тепловых машин одинаков и не зависит от рода работающего тела, а только от интервала предельных температур, в котором работает машина. Р. Клаузиус: ―Теплота не может переходить сама собой от более холодного тела к более теплому.
Важной физической величиной, используя которую можно сформулировать второе начало термодинамики в более общем виде, является еще один термодинамический параметр систем – энтропия. Энтропия - это мера неупорядоченности или вероятности состояния системы. Энтропия измеряется в тех же единицах, что и теплоемкость, - Дж/моль∙К. Энтропия является таким же свойством термодинамической системы, как температура, давление и др. В любом теле содержится определенная энтропия; как и внутренняя энергия, энтропия системы растет с ее массой и равна сумме энтропии подсистем, она увеличивается при повышении температуры за счет усиления теплового движения 70 молекул. Если атомы в молекуле упорядочены, то энтропия системы низка. Увеличение энтропии наблюдается при переходе вещества из твердого состояния в жидкое. Изменение энтропии dS системы равно отношению количества теплоты (δQ), сообщенного системе, к температуре (Т): dS = δQ/T. (6.1.5) Если ввести понятие энтропии, то второе начало термодинамики можно сформулировать так: энтропия изолированной системы возрастает в необратимом процессе и остается неизменной в обратимых термодинамических процессах: dS ≥ δQ/T. (6.1.6)
Сущность второго начала термодинамики для изолированных систем состоит в том, что все необратимые процессы протекают в направлении увеличения энтропии системы, т.е. dS > 0, dF ≠ dU. (F - свободная энергия системы). Это означает, что не вся внутренняя энергия системы переходит в работу, часть ее рассеивается в виде теплоты. При обратимых процессах dS = 0 и dF = dU, т.е. рассеивания энергии в пространстве не происходит. Энтропия - это часть общей энергии клетки, которая не может быть использована в данной системе. С увеличением энтропии энергия, вовлеченная в обмен, уменьшается, и процесс становится менее обратимым. Согласно второму началу термодинамики, энтропия изолированной системы стремится к максимуму, при котором достигается равновесие и реакция прекращается. Формулировка второго закона термодинамики для живых организмов: скорость изменения энтропии в организме равна алгебраической сумме производства энтропии внутри организма и скорости поступления энтропии из среды в организм.
Уравнение второго начала термодинамики для открытых систем следующее:
dS/dt = diS/dt + deS/dt (6.1.7)
где dS/dt - скорость изменения энтропии в организме;
diS/dt – скорость производства энтропии внутри организма;
deS/dt - скорость обмена энтропией между организмом и окружающей средой.
В отличие от первого закона термодинамики, имеющего абсолютное значение, второй закон носит статистический характер. Увеличение энтропии в необратимых процессах отражает лишь наиболее вероятное их протекание. Этот закон соблюдается особенно точно для макроскопических систем с большим количеством компонентов.
Вопросы и задания для самоподготовки
1. Применение законов термодинамики в биологии и медицине.
2. Что собой представляет термодинамичесая система? Какие типы термодинамических систем вы знаете?
5. Сформулируйте I начало термодинамики: а) для изолированной системы; б) для закрытой системы. Напишите необходимые уравнения.
6. В чем физическая сущность понятия энтропии?
7. Как связана энтропия с термодинамической вероятностью системы?
8. Сформулируйте второе начало термодинамики для изолированных и открытых систем.
Рекомендуемая литература
1. Ремизов А.Н. Медицинская и биологическая физика : учеб. для вузов / А.Н. Ремизов, А.Г. Максина, А.Я. Потапенко. – М., 2003. – С. 163- 176.
2. Артюхов В.Г. Биофизика : учеб. пособие / В.Г. Артюхов, Т.А. Ковалева, В.П. Шмелев. - Воронеж: Изд-во ВГУ, 1994. – С. 56-67.
6.2. Организм как открытая термодинамическая система. Стационарное состояние биологических систем. Уравнение Пригожина для открытой системы
Общая теория роста и развития организмов на основе представлений термодинамики была выдвинута И. Пригожиным в 1947 году. По данной теории в процессе роста и развития организмов происходит уменьшение скорости продуцирования энтропии, отнесенной к единице массы объекта:
dS = diS + deS < 0 (6.2.1)
О скорости продуцирования энтропии можно судить по теплопродукции, которая сопровождает необратимые процессы в системе и отражает уровень процессов дыхания или анаэробного гликолиза и поэтому может быть оценена по характеру поглощения О2 или образованию гликолитических продуктов. Измерения на различных объектах скорости теплопродукции dβ/dt, отнесенной к единице массы, показали, что этот параметр уменьшается, начиная с первых стадий развития организмов. Измерение удельной интенсивности дыхания имеет такую же тенденцию в те же периоды эмбрионального развития. Сходную картину можно получить и для целых организмов, и на изолированных органах и тканях. В процессе регенерации конечностей у амфибий, заживления ран у млекопитающих происходит уменьшение скорости теплопродукции или уменьшение dS/dt.
Таким образом, согласно теории И. Пригожина, в процессе развития и роста организмов скорость продуцирования энтропии непрерывно снижается и достигает минимальных значений в конечном стационарном состоянии. Благодаря более интенсивным процессам обмена веществ отток энтропии превышает ее продукцию:
diS/dt < deS/dt (6.2.2)
Жизнь - это постоянная борьба против тенденции к возрастанию энтропии. Усложнение структуры клеток в процессе эволюции и организации живых существ – мощные антиэнтропийные факторы. Термодинамическое равновесие - это состояние системы, при котором ее параметры не изменяются, и она не обменивается с окружающей средой ни веществом, ни энергией. Стационарное состояние системы характеризуется тем, что ее параметры также не изменяются во времени, но происходит обмен веществ и энергии с окружающей средой.
Сходство термодинамического равновесия и стационарного состояния системы заключается в том, что стационарное состояние, так же как и термодинамическое равновесие, сохраняет все основные параметры неизменными. Энтропия системы, находящейся в стационарном состоянии, имеет некоторую постоянную величину, не равную максимальной. Поэтому наиболее характерными свойствами стационарного состояния являются стремление системы к минимуму ежесекундного прироста энтропии и определенная внутренняя стабильность и упорядоченность.
Теорема Пригожина
И. Пригожин на основе изучения открытых систем сформулировал основное свойство стационарного состояния (1946): в стационарном состоянии при фиксированных внешних параметрах скорость продукции энтропии в открытой системе, обусловленная протеканием необратимых процессов, постоянна во времени и минимальна по величине:
diS /dt → min (6.2.3)
Таким образом, согласно теореме Пригожина стационарное состояние характеризуется минимальным рассеянием энергии. Условия стационарного процесса оказываются эквивалентными условиям минимума прироста энтропии. Это утверждение справедливо только для тех открытых систем, в которых выполняются линейные соотношения между величинами скоростей и движущих сил процессов. Кроме того, открытые системы должны находиться в частично-равновесном состоянии, при котором процессы их обмена с окружающей средой протекают равновесным образом.
Вопросы и задания для самоподготовки
1. Что собой представляет термодинамическое равновесие?
2. Сравните термодинамическое равновесие и стационарное состояние системы.
3. Как изменяется баланс энтропии при росте и старении организмов (теория Пригожина и Виам)?
4. Когда энтропия достигает своего максимального значения?
5. Охарактеризуйте устойчивое и неустойчивое стационарное состояние.
6. Как может происходить переход из одного стационарного состояния в другое? П
7. Сформулируйте теорему Пригожина.
8. Как изменяется энтропия при развитии заболевания?
Рекомендуемая литература
1. Ремизов А.Н. Медицинская и биологическая физика : учеб. для вузов / А.Н. Ремизов, А.Г. Максина, А.Я. Потапенко. – М., 2003. – С. 176- 182.74
2. Артюхов В.Г. Биофизика : учеб. пособие / В.Г. Артюхов, Т.А. Ковалева, В.П. Шмелев. - Воронеж, 1994. – С. 67-79.
6.3. Этапы моделирования, типы моделей
Моделирование - один из основных методов биофизики. При изучении сложных систем исследуемый объект может быть заменен другим, более простым, но сохраняющим основные, наиболее существенные для данного исследования свойства. Такой более простой объект исследования называется моделью. Модель — это всегда некое упрощение объекта исследования и в смысле его структуры, и по сложности внутренних и внешних связей, но обязательно отражающее те основные свойства, которые интересуют исследователя. Практически в каждой теме курса биофизики рассматриваются разнообразные модели, например, жидкостно-мозаичная модель мембраны, модель формирования потенциала действия (модель Ходжкина-Хаксли), модель скользящих нитей при описании сокращения мышцы, модель кровеносной системы (модель Франка) и др. Моделирование — это метод, при котором производится замена изучения некоторого сложного объекта (процесса, явления) исследованием его модели.
Основные виды моделей: 1) Физическая модель имеет физическую природу, часто ту же, что и исследуемый объект. Например, течение крови по сосудам моделируется движением жидкости по трубам (жестким или эластичным). При моделировании электрических процессов в сердце его рассматривают как электрический токовый диполь. Для изучения процессов проницаемости ионов через биологические мембраны реальная мембрана заменяется искусственной (например, липосомой). Физические устройства, временно заменяющие органы живого организма, также можно отнести к физическим моделям: искусственная почка – модель почки, кардиостимулятор - модель процессов в синусовом узле сердца, аппарат искусственного дыхания - модель легких; 2) Биологические модели представляют собой биологические объекты, удобные для экспериментальных исследований, на которых изучаются свойства, закономерности биофизических процессов в реальных сложных объектах. Например, закономерности возникновения и распространения потенциала действия в нервных волокнах были изучены только после нахождения такой удачной биологической модели, как гигантский аксон кальмара. Опыт Уссинга, доказывающий существование активного транспорта, был проведен на биологической модели – коже лягушки, которая моделировала свойство биологической мембраны осуществлять активный транспорт; 3) Математические модели – описание процессов в реальном объекте с помощью математических уравнений, как правило, дифференциальных. Изменяя коэффициенты или вводя новые члены в дифференциальные уравнения, можно учитывать те или иные свойства моделируемого объекта или теоретически создавать объекты с новыми свойствами, так, например, получать лекарственные препараты более эффективного действия. Можно решать сложные уравнения и прогнозировать поведение системы: течение заболевания, эффективность лечения, действие фармацевтического препарата.
Если процессы в модели имеют другую физическую природу, чем оригинал, но описываются таким же математическим аппаратом (как правило, одинаковыми дифференциальными уравнениями), то такая модель называется аналоговой. Например, аналоговой моделью сосудистой системы является электрическая цепь из сопротивлений, емкостей и индуктивностей. Основные требования, которым должна отвечать модель. 1. Адекватность - соответствие модели объекту, то есть модель должна с заданной степенью точности воспроизводить закономерности изучаемых явлений. 2. Должны быть установлены границы применимости модели, то есть четко заданы условия, при которых выбранная модель адекватна изучаемому объекту. Границы применимости определяются теми допущениями, которые делаются при составлении модели. Как правило, чем больше допущений, тем уже границы применимости. Так, например, липосома является адекватной моделью биологической мембраны, если изучается проницаемость липидного бислоя мембран для различных веществ. Если же цель исследования - электрогенез в клетках, то в этом случае липосома не адекватная модель, границы ее применимости не удовлетворяют целям исследования. Основные этапы моделирования можно свести к следующим: 1. Первичный сбор информации. Исследователь должен получить как можно больше информации о разнообразных характеристиках реального объекта: его свойствах, происходящих в нем процессах, закономерностях поведения при различных внешних условиях. 2. Постановка задачи. Формулируется цель исследования, основные его задачи, определяется, какие новые знания в результате проведенного исследования хочет получить исследователь. Этот этап часто является одним из наиболее важных и трудоемких. 3. Обоснование основных допущений. Другими словами, упрощается реальный объект, выделяются из его характеристик не существенные для целей исследования, которыми можно пренебречь. 4. Создание модели, ее исследование. 5. Проверка адекватности модели реальному объекту. Указание границ применимости модели.
Вопросы и задания для самоподготовки
1. В чем сущность метода моделирования? Какие преимущества имеет этот метод?
2. Какие типы моделей вы знаете?
Рекомендуемая литература
1. Романовский Ю.М. Математическое моделирование в биофизике. Введение в теоретическую биофизику / Ю.М. Романовский, Н.В. Степанова, Д.С. Чернавский. – М. : ИКН, 2004. – 472 с. 5.
СОДЕРЖАНИЕ
ВВЕДЕНИЕ
Тема 1. Биофизика как наука
1.1. Предмет и задачи биофизики. Проблемы современной биофизики. Значение биофизики для медицины и фармации
Тема 3. Биофизика мембран
3.1. Структура и функции биологических мембран. Динамика биомембран. Модельные липидные мембраны
3.2. Транспорт веществ через биологические мембраны. Механизмы пассивного транспорта Активный транспорт. Ионные насосы, молекулярный механизм их работы
3.3. Активный транспорт. Ионные насосы, молекулярный механизм их работы. Сопряженный транспорт
3.4. Биоэлектрические потенциалы. Механизм формирования потенциала покоя
3.5. Потенциал действия, его свойства. Биофизика нервного импульса
3.6. Биофизика рецепции. Типы клеточных рецепторов. Механизм передачи сигнала в клетку
Министерство сельского хозяйства Российской Федерации
федеральное государственное бюджетное образовательное
учреждение высшего образования
«Уральский государственный аграрный университет»
УЧЕБНО-МЕТОДИЧЕСКОЕ ПОСОБИЕ ДЛЯ САМОСТОЯТЕЛЬНОЙ ПОДГОТОВКИ СТУДЕНТОВ К КУРСУ «БИОФИЗИКА»
Составитель: Кузнецова Е.И.
Екатеринбург 2017
ВВЕДЕНИЕ
Проблема совершенствования самостоятельной работы студентов имеет исключительно большое значение в подготовке высококвалифицированных, творчески активных специалистов. Под самостоятельной работой следует понимать все виды активной познавательной и научной деятельности, требующие поисков ответов на вопросы, возникающие в ходе изучения учебного материала или проведения научных исследований.
Самостоятельная работа студентов преследует следующие цели: 1) закрепление знаний, умений и навыков студентов, их расширение и углубление; 2) формирование умения применять полученные знания в конкретных жизненных условиях, практических ситуациях; 3) выработка собственных способов, приемов, методов самостоятельного мышления, творческой активности, умения вести научный поиск.
В ходе приобщения студентов к самостоятельной работе возрастает значение внеаудиторных форм работы при непосредственном руководстве преподавателя. Специфика внеаудиторной работы студентов предусматривает обязательную работу с учебной литературой, углубление знаний в процессе выполнения заданий и самостоятельного поиска, творческий отбор материала, задач, их моделирование, овладение научной терминологией и культурой речи. Самостоятельная работа относится к наиболее эффективным средствам развития познавательной активности студентов и способствует формированию у них самостоятельности в процессе обучения в вузе.
На ветеринарном факультете в 1 семестре учебный план предусматривает организацию лекций (20 часов) и лабораторных занятий (36 часов) по биофизике. При проведении практикума по биофизике оценка знаний студентов осуществляется при сдаче работ и решении задач на данную тему. Ниже приведены разделы биофизики, изучение которых входит в учебный план. Для облегчения подготовки студента к данным темам приводятся вопросы, на которые студент должен ответить в ходе изучения материала, и список литературы, где данный материал излагается. При подготовке необходимо использовать также конспекты лекций.
Тема 1. Биофизика как наука
Предмет и задачи биофизики. Проблемы современной биофизики. Значение биофизики для медицины и фармации
Биофизика является синтетической наукой на стыке физики, химии, математики, биологии, физиологии и других наук. Существует множество определений данной науки. Одно из определений биофизики дано А.Б. Рубиным (1987): «Биофизика - наука о наиболее простых и фундаментальных взаимодействиях, лежащих в основе биологических явлений». Биофизика - интегративная наука, изучающая структуру, физические свойства и характеристики биологических объектов, фундаментальные взаимодействия молекул и молекулярных комплексов, элементарные физико-химические и физические процессы, лежащие в основе физиологических реакций и биологических явлений, а также влияние на биологические объекты различных физических факторов (света, ионизирующего излучения, температуры и др.). Биофизика как самостоятельная наука должна отвечать трем обязательным требованиям: иметь собственные цели и задачи, собственный объект (объекты) исследования и методы исследования. Основные цели биофизики: - основываясь на законах и представлениях физики и химии с широким применением математики, изучать фундаментальные элементарные процессы, протекающие в биополимерах и надмолекулярных комплексах, лежащие в основе жизнедеятельности клеток и организмов; - исследовать действие ряда физических и химических факторов на биообъекты. В задачи биофизики входит: - изучение на молекулярном уровне структуры субклеточных образований и механизмов их функционирования; - выявление общих законов (закономерностей) обмена веществ и энергии на уровне клетки и организма; - исследование молекулярных механизмов транспорта ионов, молекул через многочисленные и разнообразные мембраны поверхности разделов и фаз; - изучение молекулярных механизмов дыхания, подвижности; - исследование поглощения, размена энергии на химические превращения, влияние их на жизнедеятельность при действии энергии электромагнитных полей (видимого и ультрафиолетового излучения), проникающей радиации; - термодинамический анализ сложных систем с использованием законов классической термодинамики, а также термодинамики неравновесных процессов; - кинетический аналитический подход к изучению сложных систем и предсказание их поведения. Объектами исследования в биофизике чаще всего служат биополимеры и другие биологически важные молекулы, субклеточные комплексы, ткани, органы.
Биофизику считают относительно молодой наукой, окончательно сформировавшейся в середине XX столетия. Но это справедливо только отчасти, так как становление биофизики проходило длительное время. Условно можно выделить три этапа ее развития: I этап - с начала XVII до середины XVIII в.; II этап - с середины XVIII до середины XX в.; III этап - с середины XX в. до наших дней. I этап - накопление отдельных фактов. Делались первые попытки количественных измерений характеристик и параметров биологических объектов и систем. Первые идеи биофизики заключались в обосновании материального единства живых организмов и неорганической природы, универсальности механического движения в живой и неживой природе, первых попытках применить физические законы для объяснения ряда физиологических функций (например, кровообращения). Был создан ряд физических приборов (лупа, микроскоп и др.), позволивших установить клеточное строение живых организмов и изучать мир простейших. II этап - широкое проведение экспериментов и определение многих физико-химических параметров живых организмов. Для объяснения сложных биологических явлений привлекались законы физики и химии. III этап - формирование собственного понятийного аппарата, разработка сложных биофизических методов исследования, выделение ряда разделов в самостоятельные научные дисциплины (фотобиология, радиобиология, мембранология и др.). Для объяснения биологических процессов и явлений привлекают не только законы физики, химии, но и математики и биологии. Середина XX в. характеризуется появлением системы знаний о физико-химических фундаментальных процессах, лежащих в основе жизнедеятельности живых организмов. Изучение таких процессов и явлений, как фотосинтез, внутриклеточное дыхание, мышечное сокращение, ионные механизмы биоэлектрогенеза, механизмы мембранной проницаемости и др., потребовало разработки новых биофизических подходов и методов. Анализ полученных результатов привел к созданию системы знаний, которая отличалась от ранее существовавшей в биологии, и в частности в физиологии, переходом на субклеточный, мембранный, молекулярный уровни организации и функционирования биологических объектов. Стало очевидно, что биофизика - это не только физика живого, законов физики недостаточно для понимания процессов жизнедеятельности, необходимо привлекать биологические законы и закономерности деятельности.
В биофизике можно выделить следующие разделы: молекулярную биофизику, биофизику мембран, квантовую биофизику, термодинамику биологических процессов, прикладную биофизику. Молекулярная биофизика изучает пространственную структуру биополимеров (белков, нуклеиновых кислот, углеводов, липидов, их комплексов, надмолекулярных образований) и физические процессы, лежащие в основе их функционирования. Макромолекулы рассматривают как своеобразные машины, преобразующие энергию из одного вида в другой в пределах одной молекулы, в чем можно убедиться при анализе механизмов фотосинтеза, ферментативного катализа, фотопревращений ряда пигментов. Биофизика мембран является частью мембранологии, которая изучает структуру и функции биологических мембран. Жизнь без мембран невозможна. Разнообразие функций мембран (разграничительная, транспортная, формирования градиентов, трансформации энергии, рецепторная и др.) делает биомембраны объектом пристального внимания не только биофизиков, но и биохимиков, физиологов, иммунологов и других специалистов. Однако межмолекулярные отношения и мембранные механизмы, лежащие в основе функций живых организмов, являются предметом изучения для биофизиков. Квантовая биофизика изучает структуру электронных энергетических уровней атомов, ионов, молекул, их донорно-акцепторные свойства, электронные переходы при поглощении квантов света и пути дезактивации поглощенной энергии, химические превращения электронно- возбужденных молекул, образование фотопродуктов и молекулярные взаимодействия, лежащие в основе фотобиологических процессов и явлений. Фотобиология исследует влияние видимого и ультрафиолетового излучений на биообъекты, начиная от биополимеров и заканчивая растительными и животными организмами. В данном разделе изучаются механизмы поглощения квантов света атомами и молекулами, миграция энергии, фотохимические реакции, лежащие в основе фотобиологических процессов. Биофизика сложных систем включает в себя термодинамику и кинетику биологических процессов. Термодинамика биологических процессов анализирует функционирование биологических систем с позиций первого и второго начал термодинамики и следствий из них, используя фундаментальные физические представления. Прикладная биофизика в самостоятельный раздел отнесена весьма условно, так как в каждом разобранном ранее разделе можно выделить прикладные вопросы.
Вопросы и задания для самоподготовки
1. Что изучает биофизика?
2. Назовите основные разделы биофизики.
3. История развития биофизики. Какие этапы становления биофизики можно выделить?
Рекомендуемая литература
1. Ремизов А.Н. Медицинская и биологическая физика : учеб. для вузов / А.Н. Ремизов, А.Г. Максина, А.Я. Потапенко. – М., 2003. – С. 14-17.
2. Волькенштейн М.В. Биофизика : учеб. пособие / М.В. Волькенштейн. - М., 1988. – С. 9-22.
Дата: 2019-03-05, просмотров: 523.