Ферменты (хеликаза, топоизомераза) и ДНК-связывающие белки расплетают ДНК, удерживают матрицу в разведённом состоянии и вращают молекулу ДНК. Правильность репликации обеспечивается точным соответствием комплементарных пар оснований и активностью ДНК-полимеразы, способной распознать и исправить ошибку. Репликация у прокариот осуществляется несколькими разными ДНК-полимеразами. ДНК-полимераза I действует на запаздывающей цепи для удаления РНК-праймеров и дорепликации очищенных мест ДНК. ДНК полимераза III - основной фермент репликации ДНК, осуществляющий синтез ведущей цепи ДНК и фрагментов Оказаки при синтезе запаздывающей цепи. Далее происходит закручивание синтезированных молекул по принципу суперспирализации и дальнейшей компактизации ДНК. Синтез энергозатратный.
Цепи молекулы ДНК расходятся, образуют репликационную вилку, и каждая из них становится матрицей, на которой синтезируется новая комплементарная цепь. В результате образуются две новые двуспиральные молекулы ДНК, идентичные родительской молекуле.
Характеристики процесса репликации
матричный - последовательность синтезируемой цепи ДНК однозначно определяется последовательностью материнской цепи в соответствии с принципом комплементарности;
полуконсервативный - одна цепь молекулы ДНК, образовавшейся в результате репликации, является вновь синтезированной, а вторая - материнской;
идёт в направлении от 5’-конца новой молекулы к 3’-концу;
полунепрерывный - одна из цепей ДНК синтезируется непрерывно, а вторая - в виде набора отдельных коротких фрагментов (фрагментов Оказаки);
начинается с определённых участков ДНК, которые называются сайтами инициации репликации.
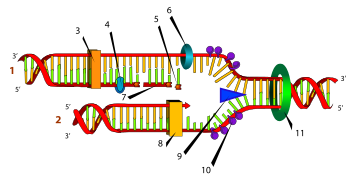
Схематическое изображение процесса репликации, цифрами отмечены: (1) запаздывающая нить, (2) лидирующая нить, (3) ДНК-полимераза (Pol α), (4) ДНК-лигаза, (5) РНК-праймер, (6) праймаза, (7) фрагмент Оказаки, (8) ДНК-полимераза (Pol δ), (9) хеликаза, (10) белки, связывающие одноцепочечную ДНК, (11) топоизомераза.
Механизмы репликации ДНК прокариот и эукариот отличаются тем, что синтез ведущей и отстающей цепей ДНК осуществляют разные ДНК-полимеразы (альфа и дельта соответственно). У E. coli обе цепи ДНК синтезируются димером ДНК-полимеразы III. У эукариот ДНК-полимераза альфа проводит инициацию синтеза ведущей цепи в точках начала репликации, а ДНК-полимераза дельта осуществляет циклические реинициации синтеза фрагментов Оказаки.
Созревание фрагментов Оказаки у эукариот требует удаления РНК-затравок с помощью 5'->3'-экзонуклеазы (белковые факторы FEN-1 или MF-1) и РНКазы H1, а также ковалентного соединения фрагментов друг с другом под действием ДНК-лигазы I.
Положение участков начала репликации в генах может иметь важное биологическое значение. Тот факт, что у ряда вирусов животных репликация начинается в определенных участках генома, позволяет предположить, что места начала репликации представляют собой специализированные последовательности в хромосомной ДНК. Среднее расстояние между местами начала репликации сравнимо со средним расстоянием между соседними петлями хроматина. Таким образом, возможно, что в каждой петле имеется лишь один участок начала репликации.
При расхождении двух репликационных вилок от одной точки начала репликации по разные стороны от этой точки родительские нуклеосомы будут попадать в разные дочерние спирали ДНК. В этом случае от точного расположения места начала репликации в транскрипционной единице будет зависеть распределение предсуществующих родительских гистонов между двумя дочерними генами. Не все нуклеосомы абсолютно одинаковы - в разных областях генетического материала структура хроматина различна. Точное положение места начала репликации в гене могло бы поэтому иметь важное биологическое значение, так как определяло бы структуру хроматина этого гена в следующем поколении клеток.
Пусковой механизм репликации ДНК явно работает по принципу «все или ничего», поскольку начавшаяся в S фазе репликация ДНК продолжается до полного завершения этого процесса. Контроль процесса репликации по принципу «все или ничего» может осуществляться по меньшей мере двумя различными способами:
1) некая общая система может специфически узнавать каждую хромосомную полосу, деконденсировть ее и тем самым делать все точки начала репликации одновременно доступными для белков, ответственных за образование репликационых пузырей;
2) репликативные белки могут узнавать лишь несколько точек начала репликации из данного набора, после чего начавшаяся локальная репликация будет изменять структуру остального хроматина репликативной единицы таким образом, что станет возможной репликация во всех других начальных точках.
Возможно, что критическим моментом в цепи событий, инициирующих репликацию ДНК, является достижение определенной стадии в процессе удвоения центриоли. По-видимому, центриоль удваивается путем матричного процесса один раз за клеточный цикл.
Пока не известно также, чем определяется фиксированная последовательность репликации хромосомных полос. Для объяснения такой последовательности было предложено две гипотезы. Согласно одной из них, различные репликативные белки, каждый из которых специфичен в отношении хромосомных полос определённого типа, синтезируются в фазе S (синтетический период интерфазы) в разное время. Согласно другой гипотезе, которая сейчас кажется более правдоподобной, репликативные белки просто действуют на те участки ДНК, которые для них более доступны; например, в течение фазы S может происходить непрерывная деконденсация хромосом, и хромосомные полосы одна за другой становятся доступными для репликативных белков.
Лекция 6. Репарация ДНК.
Репарация (от лат. reparatio - восстановление) - особая функция клеток, заключающаяся в способности исправлять химические повреждения и разрывы в молекулах ДНК, повреждённых при нормальном биосинтезе ДНК в клетке или в результате воздействия физических или химических реагентов. Осуществляется специальными ферментными системами клетки. Ряд наследственных болезней (напр., пигментная ксеродерма) связан с нарушениями систем репарации.
Начало изучению репарации было положено работами Альберта Кельнера (США), который в 1948 году обнаружил явление фотореактивации - уменьшение повреждения биологических объектов, вызываемого ультрафиолетовыми (УФ) лучами, при последующем воздействии ярким видимым светом (световая репарация).
Р. Сетлоу, К. Руперт (США) и другие вскоре установили, что фотореактивация - фотохимический процесс, протекающий с участием специального фермента и приводящий к расщеплению димеров тимина, образовавшихся в ДНК при поглощении УФ-кванта.
Позднее при изучении генетического контроля чувствительности бактерий к УФ-свету и ионизирующим излучениям была обнаружена темновая репарация - свойство клеток ликвидировать повреждения в ДНК без участия видимого света. Механизм темновой репарации облучённых УФ-светом бактериальных клеток был предсказан А. П. Говард-Фландерсом и экспериментально подтверждён в 1964 году Ф. Ханавальтом и Д. Петиджоном (США). Было показано, что у бактерий после облучения происходит вырезание повреждённых участков ДНК с изменёнными нуклеотидами и ресинтез ДНК в образовавшихся пробелах.
Системы репарации существуют не только у микроорганизмов, но также в клетках животных и человека, у которых они изучаются на культурах тканей. Известен наследственный недуг человека - пигментная ксеродерма, при котором нарушена репарация.
Источники повреждения ДНК :
Ультрафиолетовое излучение.
Радиация.
Химические вещества.
Ошибки репликации ДНК.
Апуринизация - отщепление азотистых оснований от сахарофосфатного остова
Дезаминирование - отщепление аминогруппы от азотистого основания
Дата: 2019-03-05, просмотров: 225.