ДНК содержит генетическую информацию, которая делает возможной жизнедеятельность, рост, развитие и размножение всех современных организмов. Однако, как долго в течение четырёх миллиардов лет истории жизни на Земле ДНК была главным носителем генетической информации, неизвестно. Существуют гипотезы, что РНК играла центральную роль в обмене веществ, поскольку она может и переносить генетическую информацию, и осуществлять катализ с помощью рибозимов. Кроме того, РНК - один из основных компонентов «фабрик белка» - рибосом. Древний РНК-мир, где нуклеиновая кислота была использована и для катализа, и для переноса информации, мог послужить источником современного генетического кода, состоящего из четырёх оснований. Это могло произойти в результате того, что число оснований в организме было компромиссом между небольшим числом оснований, увеличивавшим точность репликации, и большим числом оснований, увеличивающим каталитическую активность рибозимов.
К сожалению, древние генетические системы не дошли до наших дней. ДНК в окружающей среде в среднем сохраняется в течение 1 миллиона лет, а потом деградирует до коротких фрагментов. Извлечение ДНК из бактериальных спор, заключённых в кристаллах соли 250 млн лет назад, и определение последовательности генов 16S рРНК, служит темой оживлённой дискуссии в научной среде
Лекция 3. Виды генов.
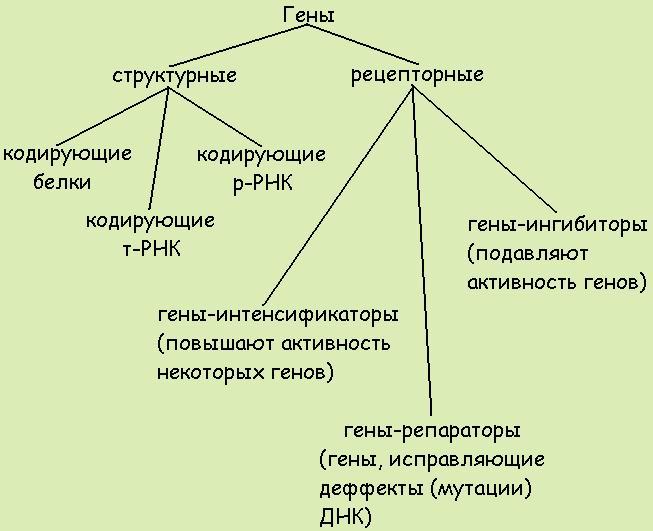
По функциям гены классифицируют на структурные и функциональные. Структурные гены содержат информацию о белках-ферментах, гистонах, о последовательности нуклеотидов в разных видах РНК.
Функциональные гены оказывают влияние на работу структурных генов. Функциональными являются гены-модуляторы и гены-регуляторы. Гены-модуляторы – это ингибиторы, интенсификаторы, модификаторы. Они усиливают, ослабляют или изменяют работу структурных генов. Регулируют работу структурных генов гены-регуляторы и гены-операторы.
Генотип всех соматических клеток организма одного вида одинаков. Но клетки разных тканей отличаются друг от друга. Вероятно, это связано с тем, что в них работают разные блоки генов. Область проявления действия данного гена называется полем его действия (например, распределение волосяного покрова на теле человека). Как правило, гены, детерминирующие определенные признаки, «работают» непостоянно (например, гены, определяющие синтез половых гормонов); их функция значительно снижается с возрастом. Период функционирования гена называется временем его действия.
По месту действия гены подразделяют на три группы
1) функционирующие во всех клетках (например, гены, кодирующие ферменты энергетического обмена);
2) функционирующие в клетках одной ткани (детерминирующие синтез белка миозина в мышечной ткани);
3) специфичные для одного типа клеток (гены гемоглобина в незрелых эритроцитах).
Гены выполняют в клетке две основные функции.
Гетеросинтетическая функция – это программирование биосинтеза белка в клетке.
Аутосинтетическая функция – репликация спирали ДНК (самоудвоение ДНК).
После открытия структуры ДНК долгое время полагали, что бактериальная хромосома представляет собой чистую ДНК в виде двойной спирали. Однако позднее выяснилось, что хромосома прокариот содержит в своей структуре примерно 20% белков. Их роль - обеспечить определенную компактизацию и прикрепление ДНК к оболочке бактерии. В настоящее время белки прокариотической хромосомы известны. Показано, что мутации в соответствующих генах не приводят к заметным фенотипическим проявлениям. По-видимому, роль этих белков вспомогательная, и они могут заменять друг друга в создании определенной структуры. Таким образом, прокариоты, в отличие от эукариот, не имеют высокоспециализированной системы организации хромосомы.
Существуют две наиболее известные модели, объясняющие механизм упаковки хроматина. Согласно одной из них, наиболее известной в зарубежной литературе, нить ДНК претерпевает пять уровней компактизации от 2 нм (ее собственный диаметр) до 1400 нм (высококонденсированная метафазная хромосома).
Низшим уровнем иерархической организации хромосом считается нуклеосомный. Нуклеосома состоит из кора (сердцевины, стержня) и намотанной на него ДНК (146 п.н„ 1,8 витка). Кор представляет собой гистоновый октамер Н2А, Н2В, НЗ, Н4 (по две молекулы каждого). Хроматин на этой стадии имеет вид «бусин» (глобул диаметром 11 нм), нанизанных на «нить» (молекулярную ДНК). Такая структура обеспечивает компактизацию примерно в 6-7 раз.
Вторая ступень компактизации - формирование хроматиновой фибриллы диаметром 30 нм. В этом процессе участвует гистон HI, который связывается с ДНК между нуклеосомными корами и сворачивает нуклеосомную фибриллу в спираль, наподобие соленоида, с шагом в 6-8 нуклеосом. Уровень компактизации на этом этапе достигает примерно 40.
Третий этап - петельно-доменный - наиболее сложный. Соленоидная фибрилла складывается, образуя петли различной длины. Общий уровень компактизации возрастает до 1000, но, очевидно, может различаться в различных районах хромосомы. Диаметр такой структуры в среднем составляет 300 нм., по-видимому, она наиболее типична для интерфазной хромосомы.
На четвертом этапе компактизации 300 нм-фибриллы дополнительно сворачиваются, образуя хроматиды диаметром примерно 600-700 нм.
Последняя, пятая, ступень компактизации (в 7000 раз) характерна для метафазной хромосомы; ее диаметр равен 1400 нм.
Известна и другая схема компактизации хроматина, предложенная Ю.С. Ченцовым. Она основана на данных световой и электронной микроскопии. Согласно этой модели, первым уровнем также является нуклеосомный. На втором этапе 8 нуклеосом образуют глобулу, называемую нуклеомером. Ряд сближенных нуклсомеров формируют 20-30-нанометровую фибриллу. Третий уровень - хромомерный. Петли фибрилл ДНП, скрепленные негистоновыми белками, образуют розетковидные структуры. На четвертом - хромонемном уровне происходит их сближение с образованием структур, состоящих из петлевых доменов. Предполагается, что на следующем, пятом, уровне компактизации, характерном для хроматид, происходит спиральная укладка хромонемных нитей.
Лекция 4. Генетический код.
Генетический код имеет много определений, например, - это система записи информации о последовательности аминокислот в белках с помощью последовательности нуклеотидов части ДНК или РНК. Более узкое определение и все таблицы генетического кода представляют его как последовательность кодонов в части молекулы мРНК. Каждый кодон состоит из трёх нуклеотидов и соответствует единственной аминокислоте. Рибосома синтезирует белок согласно инструкции, записанной в зрелой мРНК, причём соответствие между кодоном и аминокислотой определяется генетическим кодом.
Генетический код (англ. Genetic code) - совокупность правил, согласно которым в живых клетках последовательность нуклеотидов (ген и мРНК) переводится в последовательность аминокислот (белок). Собственно, перевод (трансляцию) осуществляет рибосома, которая соединяет аминокислоты в цепочку согласно инструкции, записанной в кодонах мРНК. Соответствующие аминокислоты доставляются в рибосому молекулами тРНК. Генетический код всех живых организмов Земли един (имеются лишь незначительные вариации), что свидетельствует о наличии общего предка или творца.
Правила генетического кода определяют, какой аминокислоте соответствует триплет (три подряд идущих нуклеотида) в мРНК. За редкими исключениями, каждому кодону соответствует только одна аминокислота. Конкретная аминокислота может кодироваться более чем одним кодоном, есть также кодоны, означающие начало и конец белка. Вариант генетического кода, который используется подавляющим большинством живых организмов, называют стандартным, или каноническим, генетическим кодом. Однако известно несколько десятков исключений из стандартного генетического кода, например, при трансляции в митохондриях используются несколько изменённые правила генетического кода. Простейшим представлением генетического кода может служить таблица из 64 ячеек, в которой каждая ячейка соответствует одному из 64 возможных кодонов.
Попытки понять, каким образом последовательность ДНК кодирует аминокислотную последовательность белков, начали предприниматься почти сразу же после того, как в 1953 году была установлена структура ДНК. Георгий Гамов предположил, что кодоны должны состоять из трёх нуклеотидов, чтобы кодонов хватило для всех 20 аминокислот (всего же возможно 64 различных кодона из трёх нуклеотидов). В 1961 году триплетность генетического кода удалось подтвердить экспериментально. В том же году Маршалл Ниренберг и его коллега Генрих Маттеи использовали бесклеточную систему для трансляции in vitro. В качестве матрицы был взят олигонуклеотид, состоящий из остатков урацила (UUUU…). Пептид, синтезированный с него, содержал только аминокислоту фенилаланин. Так впервые было установлено значение кодона: кодон UUU кодирует фенилаланин. Дальнейшие правила соответствия между кодонами и аминокислотами были установлены в лаборатории Северо Очоа. Было показано, что полиадениновая РНК (ААА…) транслируется в полилизиновый пептид, а на матрице полицитозиновой РНК (ССС…) синтезируется пептид, состоящий только из остатков пролина. Значение остальных кодонов было установлено при помощи разнообразных сополимеров в ходе экспериментов, проведённых в лаборатории Хара Гобинда Кораны. Вскоре после этого Роберт Холли установил структуру молекулы тРНК, которая служит посредником при трансляции. В 1968 году Ниренберг, Корана и Холли были удостоены Нобелевской премии по физиологии и медицине.
После установления правил генетического кода многие учёные занялись его искусственными преобразованиями. Так, начиная с 2001 года в генетический код были внедрены 40 аминокислот, которые в природе не входят в состав белков. Для каждой аминокислоты создавались свой кодон и соответствующая аминоацил-тРНК-синтетаза. Искусственное расширение генетического кода и создание белков с новыми аминокислотами могут помочь глубже изучить структуру белковых молекул, а также получить искусственные белки с заданными свойствами. Х. Мураками и М. Сисидо смогли превратить некоторые кодоны из трёхнуклеотидных в четырёх- и пятинуклеотидные. Стивен Бреннер получил 65-й кодон, который был функционален in vivo.
В 2015 году у бактерии Escherichia coli удалось изменить значение всех кодонов UGG с триптофана на тиенопиррол-аланин, не встречающийся в природе. В 2016 году был получен первый полусинтетический организм - бактерия, геном которой содержал два искусственных азотистых основания (X и Y), сохраняющихся при делении. В 2017 году исследователи из Южной Кореи заявили о создании мыши с расширенным генетическим кодом, способной синтезировать белки с аминокислотами, не встречающимися в природе.
Дата: 2019-03-05, просмотров: 824.