Коллоидные ПАВ обладают невысокой истинной растворимостью. Растворы, в которых они молекулярно диспергированы, могут иметь максимальную концентрацию от 10–6 до 10–3 моль/л. Это следствие наличия у молекул ПАВ длинных углеводородных радикалов. 83
Малая растворимость ПАВ влечет за собой ассоциацию их молекул, с ростом концентрации переходящую в мицеллообразование. Концентрация раствора ПАВ, при которой начинают образовываться мицеллы, называется критической концентрацией мицеллообразования (ККМ).
Изотермы поверхностного натяжения коллидных ПАВ отличаются от изотерм истинно растворимых ПАВ более резким понижением поверхностного натяжения с увеличением концентрации и наличием излома на изотерме (рис. 7.1). Концентрация в точке из лома соответствует критической концентрации мицеллообразования, выше которой в растворе самопроизвольно протекает процесс образования мицелл и истинный раствор переходит в ультрамикрогетерогенную систему — золь.
Механизм термодинамики мицелообразования
С ростом концентрации ионы или молекулы ПАВ выходят в поверхностный слой на границе раздела фаз и уменьшают поверхностно е натяжение раствора. Когда поверхностный слой полностью заполнен (это состояние соответствует достижению ККМ), в системе начинается ассоциация углеводородных радикалов молекул ПАВ. В результате ассоциации образуются мицеллы, внутренняя часть которых, так называемо е ядро, состоит из плотно упакованных, объединившихся радикалов, практически это жидкий углеводород. Поляр ные группы молекул ПАВ ориентированы в воду. Они образуют гидрофильную оболочку, которая изолирует ядро мицеллы от воды (рис. 7.2). Гидрофильность оболочек мицелл придает образовавшейся гетерогенной системе лиофильность, а, значит, и термодинамическую устойчивость.
При дальнейшем росте концентраций ПАВ (выше ККМ) количество необъединенных в мицеллы мо лекул или ионов ПАВ в поверхностном слое остается постоянным, но увеличивается количество мицелл.
Движущей силой образования мицелл являются гидрофобные взаимодействия. Энтальпия взаимодействий углеводородных радикалов ПАВ друг с другом меньше энтальпии взаимодействия их с водой. Для системы термодинамически выгоднее состояние с минимумом энтальпии, поэтому радикалы выталкиваются из водной среды в ядра мицелл, чтобы избежать, насколько это возможно, контакта с водой. В результате этого энтальпия уменьшается.
Когда молекулы или ионы ПАВ находятся в неассоциированном состоянии, вокруг их углеводородных радикалов из молекул воды образуются льдоподобные упорядоченные структуры. Переход радикалов ПАВ из воды в мицеллы разупорядочивает структуру воды, вследствие чего повышается энтропия системы.
Уменьшение энтальпии и увеличение энтропии приводит к снижению энергии Гиббса системы:
G = H – TS < 0.
Для процесса мицеллообразования уменьшение энергии Гиббса вс ледствие указанных изменений термодинамических параметров составляет пример - но 2600 Дж/мо ль на каждую –CH2– группу углеводородных радикалов. Поэтому процесс образования мицелл термодинамически выгоден и идет самопроизвольно.
Термодинамическая выгодность мицеллообразования в растворах коллоидных ПАВ обусловлена также еще и тем, что гидрофильность образующейся оболочки обеспечивает минимальное межфазное натяжение на границе мицелла—вода. Сравнительно небольшая поверхностная энергия системы компенсируется энтропийным фактором, вызывающим равномерное распределение мицелл в дисперсионной среде.
Процесс мицеллообразования обратим: при разбавлении до концентраций меньших ККМ мицеллы распадаются на ионы или молекулы (их назыв ют мономерами в отличие от мицелл) и дисперсная ультрамикрогетерогенная система переходит в молекулярный истинный раствор. Это говорит о том, что мицеллы находятся в термодинамическом равновесии с мономерами ПАВ.
Таким образом, мицеллярные системы в раствор ах коллидных ПАВ являются достаточно редким примером самопроизвольно образующихся термодинамически устойчивых лиофильных гетерогенных систем — равновесных систем с минимумом энергии Гиббса, несмотря на имеющуюся у них огромную межфазную поверхность.
Методы определения критической концентрации мицеллообразования
Для всех мицеллообразующих ПАВ критическая концентрация мицеллообра-зования лежит в области 10–6 – 10–3 моль/л. Значения ККМ зависят от разных факторов: от длины углеводородных радикалов, от наличия в растворе электролитов или органических растворителей, от характера полярной группы.
Значение ККМ уменьшается с ростом длины углеводородного радикала ПАВ, так как при этом уменьшается истинная растворимость ПАВ. Введение электролитов снижает ККМ ионогенных ПАВ из-за уменьшения гидратируемости (а, следовательно, и ухудшения растворимости) полярных частей молекул ПАВ и слабо влияет на ККМ неионогенных ПАВ. Добавление в раствор органических полярных растворителей увеличивает ККМ вследствие усиления растворяющей способности среды.
Все эти закономерности установлены на основании экспериментального определения ККМ коллоидных ПАВ.
Возникновение мицелл в растворе ПАВ сопровождается резким экспериментально фиксируемым изменением физико-химических свойств системы, которое в большинстве случаев выражается в появлении характерных изломов на кривых зависимости физико-химических параметров от концентрации ПАВ (рис. 7.7).
Более четко точка излома выявляется на кривых зависимости свойств ПАВ от концентрации в координатах "свойство – lg c".
Наиболее распространенными методами определения ККМ являются кондуктометрический и сталагмометрический.*
Кондуктометрический метод определения ККМ применяется для ионогенных ПАВ. Он основан на концентрационной зависимости эквивалентной электропроводности растворов этих ПАВ. При концентрациях, соответствующих ККМ, на графике зависимости эквивалентной электропроводности наблюдается излом, так как при этих концентрациях начинают формироваться ионные мицеллы, окруженные диффузным слоем противоионов. Подвижность ионов при этом снижается и электропроводность с увеличением концентрации ПАВ уменьшается резче, чем до критической концентрации мицеллообразования (рис. 7.7, ).
В сталагмометрическом методе определения ККМ измеряется поверхностное натяжение водных растворов ПАВ, которое резко уменьшается с ростом концентрации вплоть до ККМ, а затем остается практически постоянным (рис. 7.7., ). Этот метод применим для ионогенных и для неиногенных коллоидных ПАВ

63. Высокомолекулярные вещества (ВМВ). Классификация. Структура. Форма
макромолекул и типы связей между ними. Гибкость макромолекул.


Высокомолекулярные соединения (ВМС) получили свое название из-за очень большой относительной молекулярной массы (не менее 10000–15000). Если относительная молекулярная масса обычных низкомолекулярных веществ редко достигает нескольких сотен, например, воды – 18; диоксида углерода – 44; глюкозы – 180, то молекулярная масса натурального каучука (ВМС) находится в пределах от 7 104 до 2,5 106. Молекулы ВМС из-за их большой молекулярной массы называют обычно макромолекулами.
Высокомолекулярные соединения из-за их большой молекулярной массы нелетучи и не способны перегоняться, весьма чувствительны к воздействию внешних факторов. Макромолекулы легко распадаются под воздействием самых незначительных количеств кислорода и других деструктирующих агентов. Большинство ВМС при повышении температуры размягчаются постепенно и не имеют определенной температуры плавления. Температура разложения этих веществ ниже температуры кипения, т. е. ВМС могут находиться только в конденсированном состоянии.
Свойства ВМС зависят не только от размера, но и от формы молекул. Так, ВМС с изодиаметрическими молекулами (гемоглобин, гликоген) – порошкообразные вещества, при растворении почти не набухают, а растворы их не обладают высокой вязкостью. ВМС с сильно асимметричными вытянутыми молекулами (желатин, целлюлоза, каучук) при растворении очень сильно набухают и образуют высоковязкие растворы.
ВМС с линейными молекулами обладают волокнистой структурой, обусловливающей анизотропию свойств и высокую механическую прочность, поэтому они образуют волокна и пленки. Некоторые полимеры высокоэластичны.
Природные и синтетические ВМС. К наиболее важным природным ВМС относятся белки, являющиеся главной составной частью всех веществ животного и растительного происхождения. Молекулы белков построены из различных аминокислот, содержащих ионогенные группы (СООН, NH3OH) и соединенных пептидными связями. Молекулярная масса различных белков колеблется в широких пределах и достигает нескольких миллионов. Белки используются в основном для пищевых целей. Однако ряд содержащих белки веществ (кожа, шерсть, шелк) находят и техническое применение.
Природными ВМС являются и полисахариды – крахмал, целлюлоза. Целлюлоза – составная часть тканей растений. Наибольшее содержание целлюлозы (96–98 %) в хлопке. Молекулы целлюлозы имеют линейное строение и составлены из остатков глюкозы. Молекулярная масса колеблется от 500 тыс. до миллионов. Крахмал – составная часть всех злаков и картофеля. Макромолекулы крахмала состоят из звеньев глюкозы, и он является не индивидуальным веществом, а смесью полисахаридов, отличающихся размерами молекул и строением. Путем гидролиза из крахмала получают декстрин, патоку и глюкозу.
К природным ВМС относится и натуральный каучук – материал, обладающий высокой эластичностью. Его добывают из млечного сока (латекса) некоторых растений – каучуконосов. Это углеводород, состоящий из изопентеновых (изопреновых) остатков. При нагревании с серой каучук подвергается вулканизации, образуя резину, эбонит.
Помимо природных ВМС в технике и быту применяют синтетические ВМС. К ним относятся синтетические каучуки и различные синтетические полимеры. Эти продукты чрезвычайно разнообразны по химическому составу и свойствам, они являются заменителями природных ВМС, их используют для получения пластмасс, синтетических тканей и др. Производство синтетических ВМС значительно превысило производство конструкционных материалов (алюминиевых и магниевых сплавов и др.). К достоинствам синтетических ВМС относится то, что их можно получить с заранее заданными свойствами, подбирая необходимые исходные материалы и регулируя технологический процесс.
Строение макромолекул и структура ВМС. Молекулы ВМС могут быть линейными и разветвленными. Именно линейной формой макромолекул определяются типичные свойства полимеров: эластичность, способность образовывать нити и пленки высокой прочности, набухать, давать при растворении вязкие растворы и т. д. Эти типичные для полимеров свойства определяются гибкостью их линейных молекул, способностью к колебательно-вращательному движению отдельных звеньев макромолекул вокруг соединяющих их отдельных связей. Благодаря вращению отдельных звеньев, макромолекула изгибается и может принимать различные конформации. Конформации – формы молекул, переходящие друг в друга без разрыва химической связи за счет свободного поворота звеньев.
Гибкость макромолекул – одна из важнейших характеристик полимера, определяющая его основные макроскопические свойства. Гибкость макромолекул − это способность полимерных цепей изменять свою конформацию в результате внутримолекулярного теплового движения звеньев (термодинамическая гибкость) или же под действием внешних механических сил (кинетическая гибкость). Гибкость макромолекул обусловлена тем, что мономерные звенья цепи при тепловом движении или при внешних силовых воздействиях вращаются вокруг одинарных (s-) связей.
64.Свойства растворов ВМС. Особенности растворения ВМС как следствие их структуры. Форма макромолекул. Механизм набухания и растворения ВМС. Зависимости величины набухания от различных факторов.



65.Аномальная вязкость растворов ВМС. Уравнение Штаудингера. Вязкость крови и других биологических жидкостей.




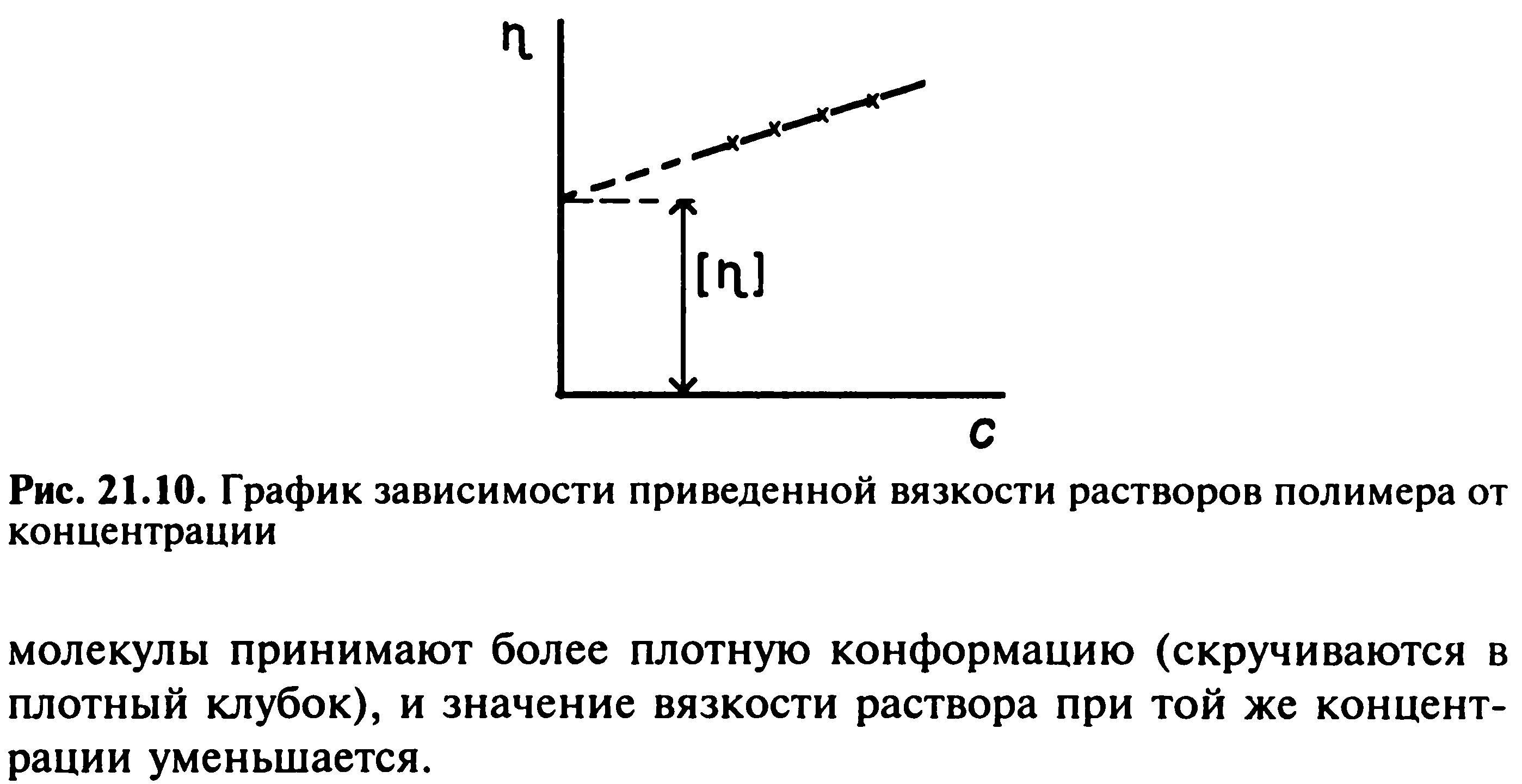

66. Полиэлектролиты. ИЭТ, методы ее определения. Осмотическое давление р-ров биополимеров. Уравнение Галлера


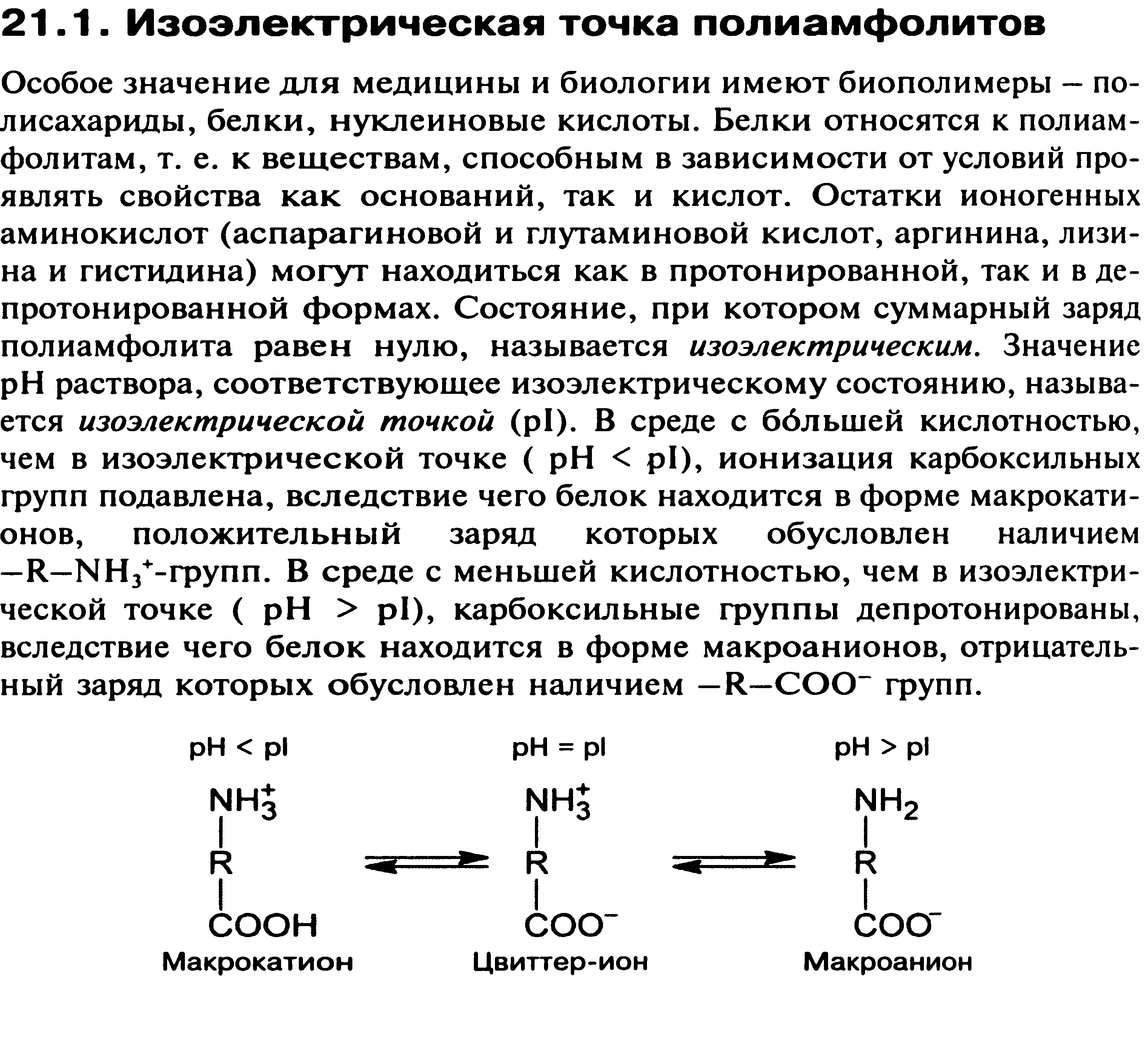
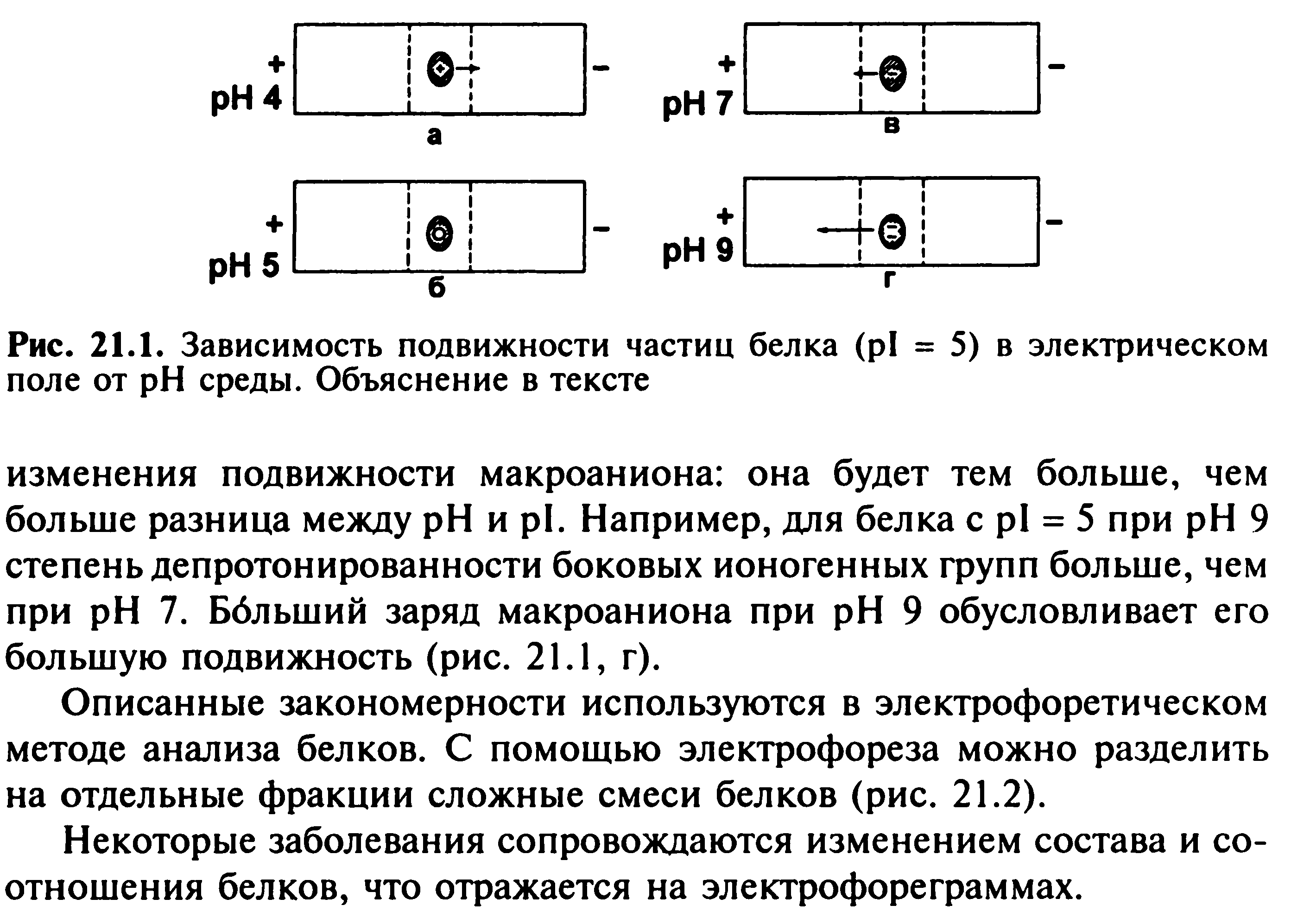

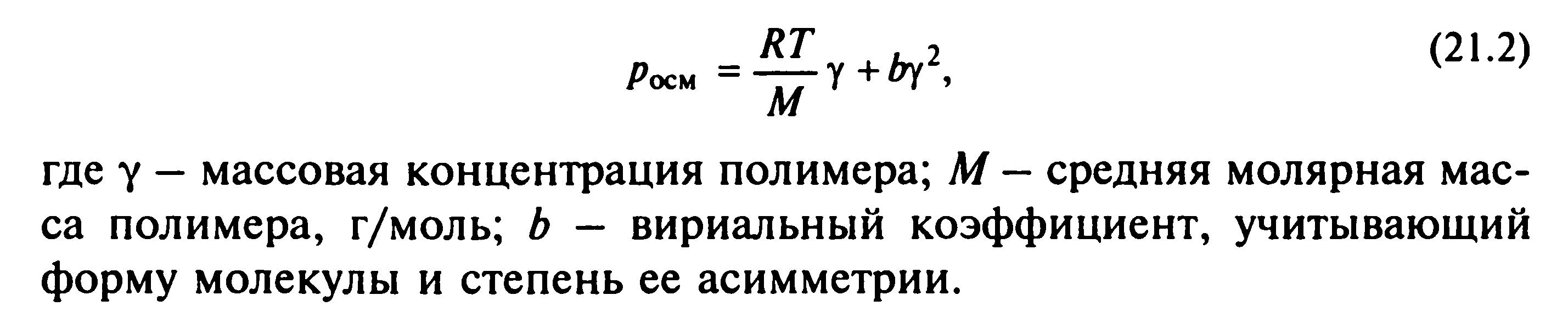
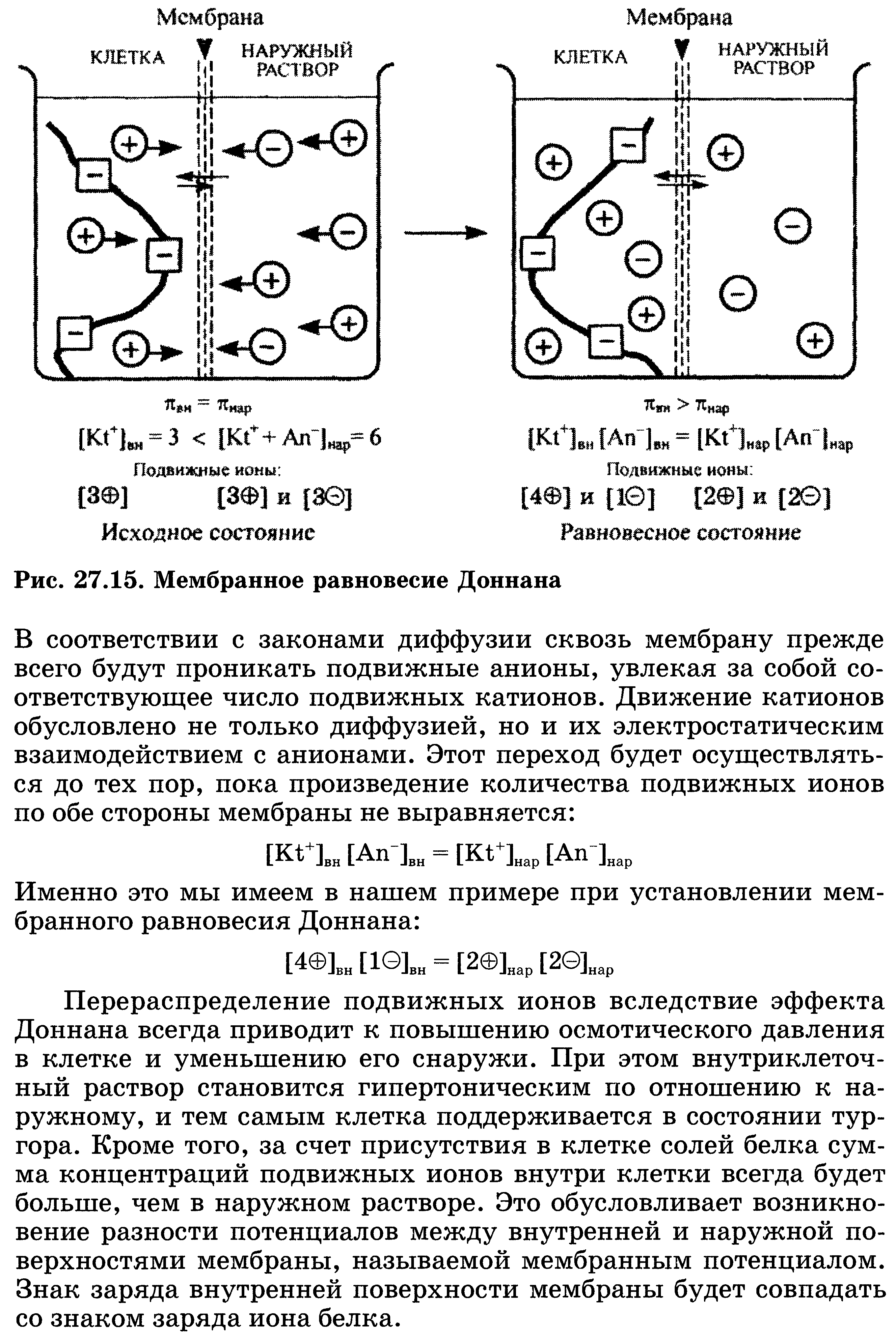
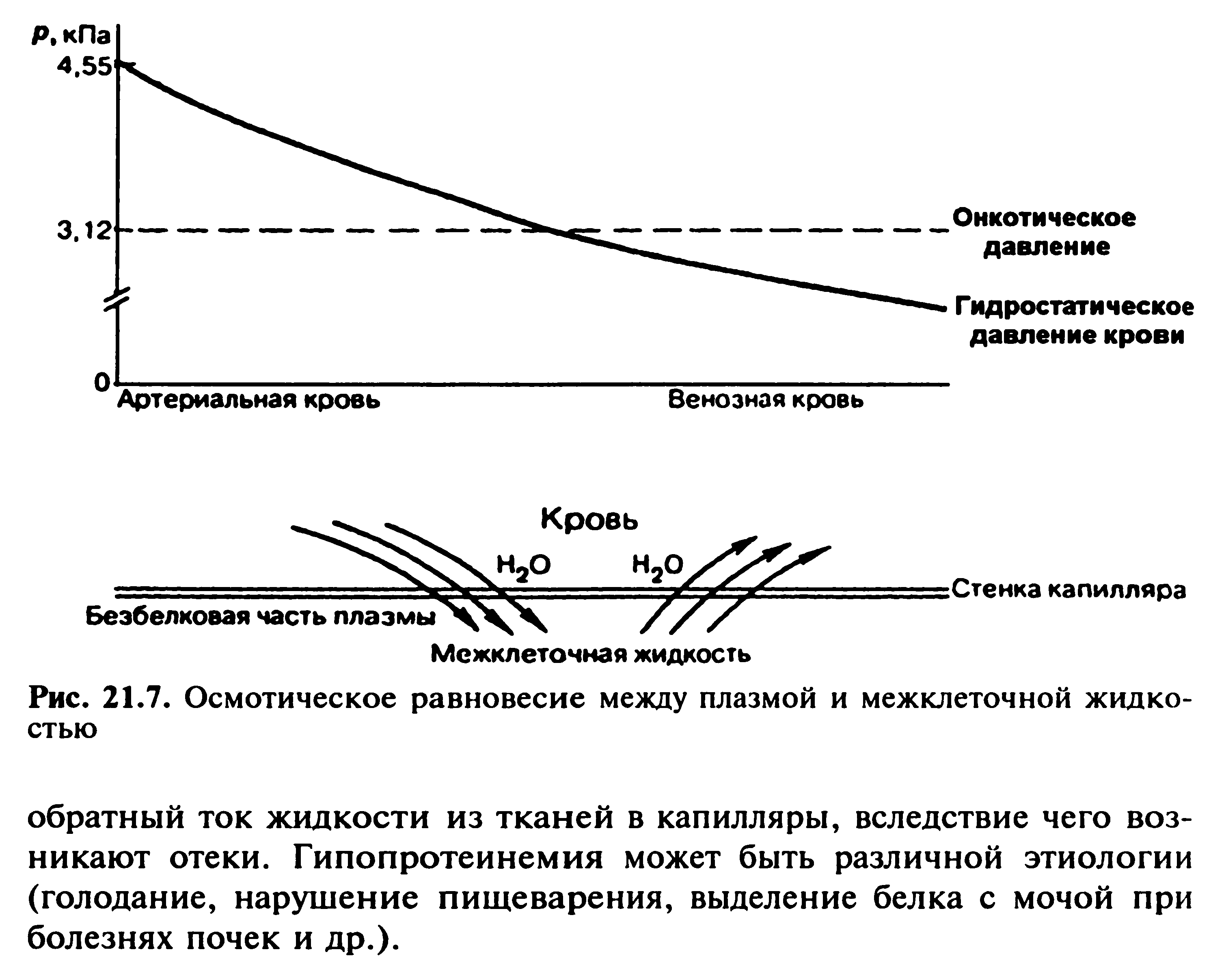
67. Мембранное равновесие Доннана. Онкотическое давление плазмы и сыворотки крови, его биологическая роль.



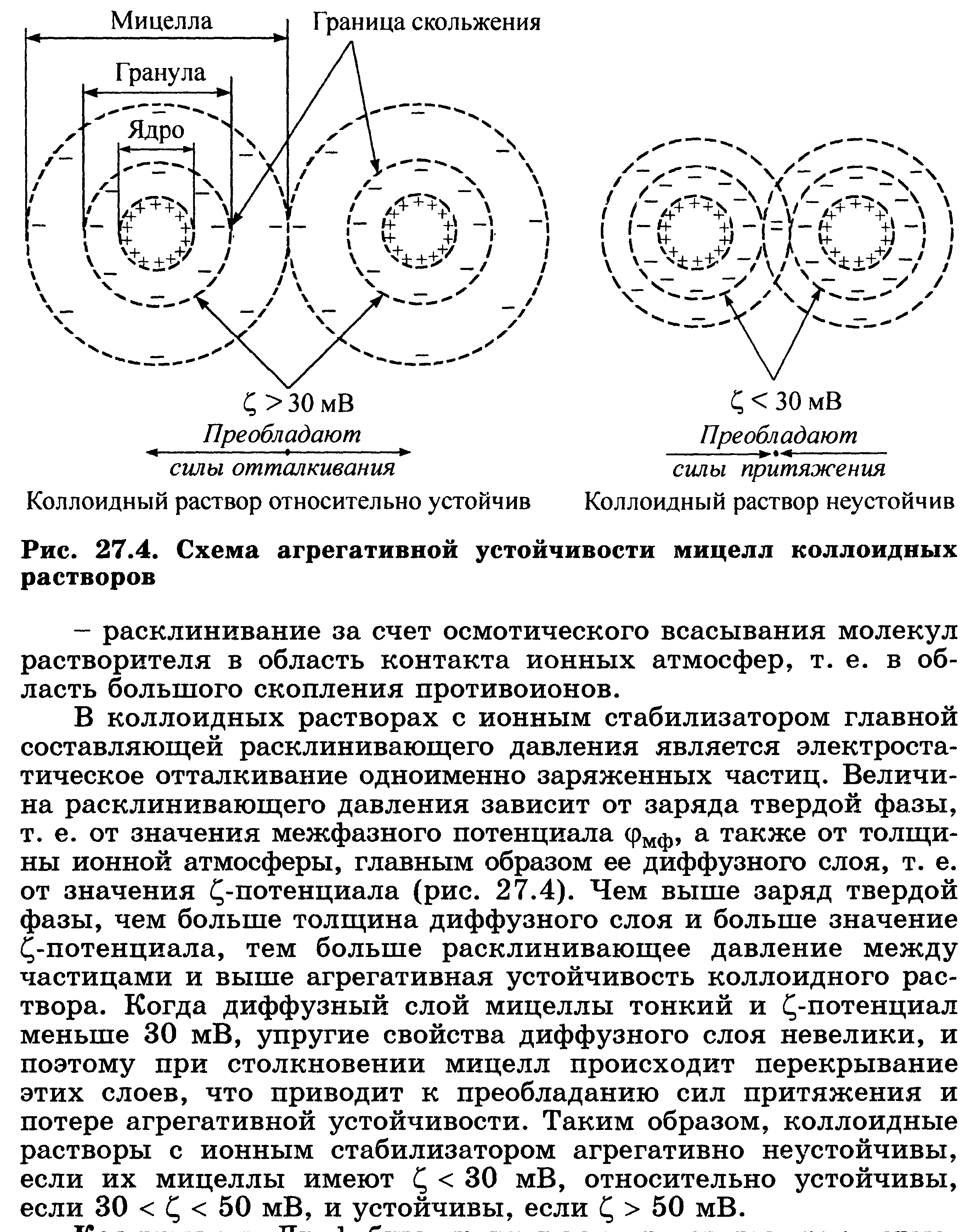
68. Устойчивость р-ров биополимеров. Высаливание биополимеров из р-ра. Коацервация, ее роль. Застуневание. Св-ва студней: синерезис, тиксотропия. Примеры.






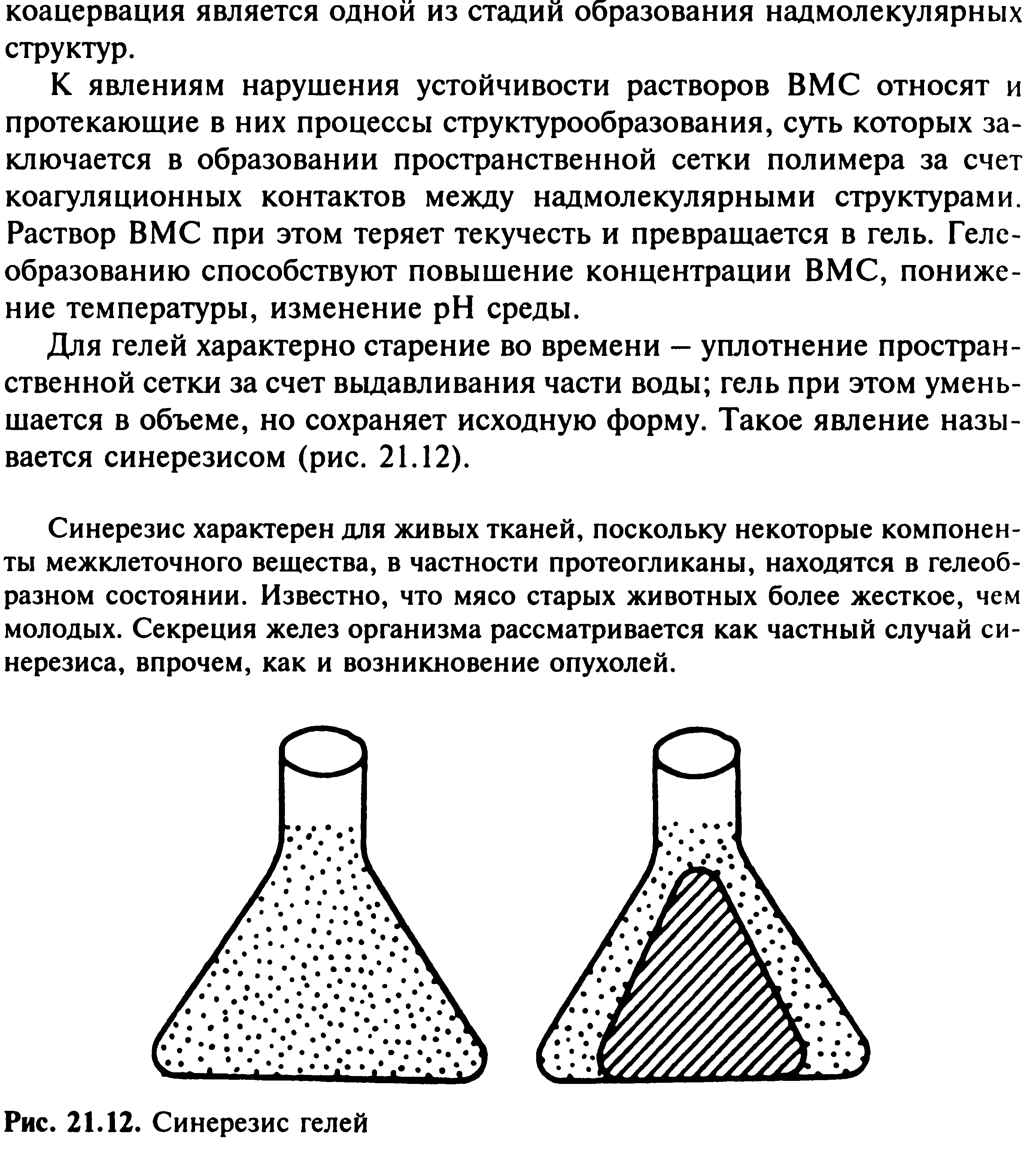

Дата: 2019-02-19, просмотров: 505.