По поводу увеличения размера тела или изменения длины ног, верующий дарвинист вполне может Вам сказать (апеллируя к «очевидности»):
- Но ведь это же очевидно (!) что если теперь подождать несколько тысяч (или миллионов) лет, то установленное давление естественного отбора, например, в случае конкуренции между ящерицами, благоприятствующее большому размеру тела (см. выше), в конце концов приведет к новому биологическому виду ящериц, отличающемуся от вида предка, в первую очередь, своими размерами!
Здесь Вам надо резко осечь полёт фантазии дарвиниста. Вам надо сказать ему буквально следующее:
- Точно такое же давление естественного отбора, благоприятствующее большим размерам тела, наблюдается по факту, например, в популяциях Homo sapiens (среди мужчин этого вида). Очевидно, что такое благоприятствование должно было бы привести (в ходе естественного отбора за тысячи поколений людей) примерно вот к такой картинке (Рис. 12):

Рисунок 12. Теоретически ожидаемый результат отбора (на увеличение общего размера тела) у человека (Homo sapiens) в результате внутривидовой конкуренции и полового отбора.
Или хотя бы к такой (Рис. 13):

Рисунок 1 3 . Теоретически ожидаемый результат отбора (на увеличение врожденной атлетичности) у мужчин Homo sapiens в результате внутривидовой конкуренции и полового отбора.
Однако мы почему-то не наблюдаем ни увеличения размеров тела человека (или хотя бы мужчин), ни повышения степени его атлетичности в ходе эволюции. Даже под столь очевидным давлением отбора. Вот тебе и «очевидно».
Надежно установлено, что после этого дарвинист сразу же (!) перейдёт к теоретическим рассуждениям о мощной роли стабилизирующего отбора. То есть, о влиянии разнообразных и нередко противоположных факторов окружающей среды, оказывающих разнонаправленное действие. Например, о недостатке пищи, который препятствует безудержному увеличению размеров тела у человека...
Вот после этого сразу же (!) посоветуйте дарвинисту перевести все его рассуждения (почему «эволюция ничаво не шмогла»)... обратно на «эволюционирующих ящериц». И тогда дарвинист, может быть, поймёт - то, что ему «очевидно» (в его голове), совсем не обязательно существует в реальности.
После этого следует развить эту мысль дальше, отметив, что совсем не обязательно ограничивающими факторами, делающими эволюцию признака невозможной, являются именно факторы внешней среды. Вполне можно предположить, что подобные ограничения может накладывать и некая внутренняя системность, свойственная тому или иному биологическому виду.
Предложите дарвинисту обратить внимание на самых больших людей мира (Рис. 14):

Рисунок 14. Один из самых высоких людей мира встретился с одним из самых маленьких людей мира.
И после того, как дарвинист поизучает соответствующие картинки и видеозаписи, ему станет очевидно, что мешает увеличению размеров в данном случае, совсем не «недостаток пищи», а некие внутренние ограничения (для человека, как биологического вида), природу которых мы, может быть, пока не слишком понимаем. Мы еще не знаем, как конкретно работает механизм построения организма в онтогенезе. В том числе, не знаем о возможной роли системных видовых ограничений в этом процессе. Пока мы можем ориентироваться только на внешние проявления того или иного признака. И судя по внешним проявлениям, приближение морфологического признака к границам каких-то предельных значений (характерных для этого вида), приводит к явной патологии, разрушению системности, свойственной этому биологическому виду. Например, мы видим, что двигательные возможности обсуждаемых гигантов - резко ограничены. И совсем не потому, что они недоедают, а просто потому, что они слишком большие для данного биологического вида. Между тем, у каких-нибудь медведей, тигров, или горилл, размеры которых еще больше... мы совершенно не наблюдаем столь катастрофического падения двигательных возможностей.
И падение двигательных возможностей здесь далеко не единственная патологическая черта. Так же сильно падает выносливость. И наверное, снижается скорость реакции. Больные гигантизмом жалуются на головную боль, общую слабость, онемение в руках, боли в суставах, ограничение и болезненность движений, снижение памяти, изменение внешнего вида, ухудшение зрения, сухость во рту и жажду. Нарушается функция щитовидной железы, половых желез, надпочечников, возможно развитие сахарного диабета. Увеличивается размер внутренних органов: легких, сердца, печени, селезенки, поджелудочной железы, кишечника. В результате может развиться функциональная недостаточность этих органов, например, сердечная недостаточность. Повышается вероятность образования опухолей. Почти у всех женщин нарушается менструальный цикл. Примерно у трети мужчин развивается половая слабость.
Всё озвученное справедливо не только в отношении увеличения размеров тела. Примерно то же самое (т.е. разнообразные патологии) мы наблюдаем и при излишнем уменьшении размеров (карликовость).
В общем, трудно поверить, что наблюдаемое биологическое разнообразие Земли (со столь совершенными живыми существами) образовалось за счёт таких вот отклонений от нормы. Действительно, вряд ли теоретический «естественный отбор увеличения размеров тела» мог бы проходить через людей, которым трудно не только бегать, но даже (нередко) просто ходить, причем настолько, что они вынуждены опираться на костыли.
Поэтому вопрос - можно ли вообще так изменить системность (того или иного) биологического вида, чтобы это привело к настоящей эволюции какого-нибудь морфологического признака - этот вопрос пока остаётся не просто открытым, а скорее, ответ на этот вопрос отрицательный (судя по тому, что мы реально видим). То есть, даже если естественный отбор и будет благоприятствовать, например, большим размерам тела, то отсюда еще совсем не следует (автоматически), что представители этого биологического вида в результате начнут становиться всё больше и больше (без всяких ограничений) в ходе «свободной эволюции». Наоборот, исходя из наблюдений за реальными подобными случаями, нам скорее следует сделать обратный вывод.
И это мы взяли такой признак (рост), который у человека имеет очень большие значения внутривидовой изменчивости. По другим признакам размах внутривидовой изменчивости у человека может быть гораздо меньше. Например, различия в нормальной температуре тела между разными людьми колеблются в пределах не более 5%. Любые более сильные отклонения свидетельствуют о возникновении или наличии какой-то болезни. То есть, отклонения в температуре тела более 5% уже приводят к общему падению приспособленности. И это несмотря на то, что температура тела зависит от определенного гормонального фона. Который в свою очередь (казалось бы) вполне мог бы изменяться в результате случайной эволюционной изменчивости. Но нет. Мы имеем то, что имеем - нормальная температура тела (здорового) человека колеблется в пределах не более 5%.
Такие факты никак не поддерживают идею «свободной эволюции без границ».
Поэтому если дарвинист пристаёт к Вам со своим первым символом веры (первым религиозным догматом):
- Эволюция это факт!
Скажите ему, что если он хочет кому-нибудь доказать справедливость этого догмата (а не просто в него верить), то доказывать его вот такими фразами:
- Эволюция это факт, потому что это очевидно!
Такими фразами доказывать «факт эволюции» нельзя. Если ему лично что-то кажется «очевидным», то это еще не значит, что то же самое будет очевидно и другому. И тем более не значит, что это «очевидно» будет наблюдаться в реальной природе.
Доказывать, что «эволюция - это факт» нужно, собственно, приведением соответствующих фактов (а не апелляцией к очевидности). И вот тут у нас как раз и «глухо». Во всяком случае, в отношении «наблюдаемой эволюции какого-нибудь морфологического признака».
Итак, давайте подведем предварительные итоги по поводу «эволюции» ящериц. Они пока плачевны - мы пока не нашли ни одного строго установленного примера, про который можно было бы сказать точно, что это именно «наблюдаемая эволюция». Пока в приведенных примерах мы видели только изучение давления естественного отбора (в разных условиях). А вот эволюции мы в этих примерах так и не увидели.
А именно, мы нигде не увидели, чтобы под давлением естественного отбора какой-либо морфологический признак вышел бы за границы значений, наблюдавшихся у этого вида раньше. Всё, что мы видели, это удаление из популяций выборочных значений того или иного признака (против которых работает естественный отбор), из уже имеющегося массива значений этого признака, характерных для этого вида.
Тем не менее, «наблюдаемая эволюция ящериц» - это уже, наверное, «притча во языцех» на устах многих глубоко верующих дарвинистов.
ЯЩЕРИЦЫ И ВОЛШЕБСТВО
Так что же получается? Что так сильно раскрученная «наблюдаемая эволюция ящериц», на самом деле… пшик на ровном месте?
В целом, да. Но за исключением только одного удивительного примера. Есть одно (уникальное) опубликованное исследование, где действительно был установлен «выход морфологических признаков за пределы значений, наблюдаемых у этого вида раньше». И даже не просто выход, а скорее уж, «выбегание». Такое «выбегание», что остаётся только изумленно хлопать ресницами. Лучше всего назвать результаты, опубликованные в этом исследовании, эволюцией по волшебству. Понятно, что такие вещи надо разбирать подробно. Чем мы сейчас и займемся.
В предыдущих главах, посвященных дискуссии с воображаемыми и реальными дарвинистами, я выборочно показал, что доказательная база эволюционного учения в области «наблюдаемой эволюции» - практически нулевая. А именно, почти не имеется зафиксированных (в публикациях) фактов эволюционных изменений хотя бы какого-нибудь морфологического признака (у хотя бы какого-нибудь биологического вида), которые бы несомненно вышли за пределы ранее наблюдавшихся значений, характерных для того или иного биологического вида.
Сейчас же мы сосредоточим своё внимание именно на таких установленных примерах, в которых была зафиксирована эволюция тех или иных признаков. То есть, признаки действительно приняли значения, не наблюдавшиеся у этих видов раньше. Таких примеров установлено очень мало, но они есть, и они очень интересные.
ЭВОЛЮЦИЯ ИТАЛЬЯНСКИХ ЯЩЕРИЦ
Сразу в двух опубликованных научных статьях исследователи поведали интересные подробности из жизни новой популяции итальянских стенных ящериц (Podarcis sicula). Приведу полные названия этих статей, поскольку уже сами названия этих статей весьма говорящие:
1. «Различия в морфологии, эффективности и поведении между недавно разошедшимися популяциями Podarcis sicula отражают различия в хищническом давлении» (Vervust et al., 2007).
2. «Быстрое крупномасштабное эволюционное расхождение в морфологии и эффективности, связанное с разными пищевыми ресурсами» (Herrel et al., 2008).
Вся история началась с того, что в 1971 году ученые выпустили на один из островков Адриатического моря (Pod Mrcaru) пять пар взрослых итальянских стенных ящериц Podarcis sicula (пять самок и пять самцов). Ученые взяли этих ящериц с соседнего островка Pod Kopiste.
Оба этих островка (Pod Mrcaru и Pod Kopiste) похожи между собой по размеру, высоте и микроклимату. На всякий случай, отметим, что на островке Pod Mrcaru в 1971 году обитал еще и другой вид ящериц из этого же рода - Podarcis melisellensis.
В 2004 году исследователи вернулись на остров Pod Mrcaru, чтобы посмотреть, как поживают переселенные ящерицы (Podarcis sicula). Биологи обнаружили, что второй, аборигенный вид ящериц (Podarcis melisellensis) к этому времени полностью вымер, и ящеричное население островка Pod Mrcaru представлено только особями итальянской стенной ящерицы (Podarcis sicula).
Помимо этого, биологи обнаружили целую плеяду интересных различий между предковой и новой популяцией стенных ящериц:
1. Плотность населения ящериц на островке Pod Mrcaru оказалась намного выше, чем на исходном островке Pod Kopiste (откуда были взяты первые пять пар ящериц).
Кроме того, представители новой популяции ящериц оказались:
2. В целом немного больше по размеру.
3. Их ноги оказались короче, чем у представителей предковой популяции.
4. Они слабее реагировали на угрозу хищника.
5. Они развивали меньшую скорость бега.
6. Их головы оказались пропорционально больше в длину, ширину и высоту.
7. Сила укуса оказалась выше, чем у представителей предковой популяции (Табл. 2):
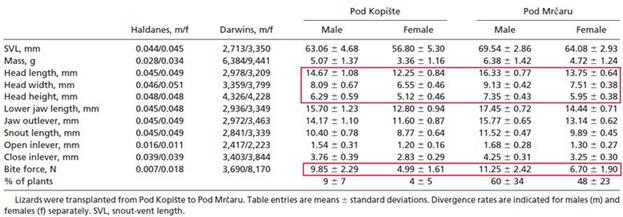
Таблица 2. Из работы ( Herrel et al ., 2008), иллюстрирующая изменения в длине, высоте, ширине головы и силе укуса (обведено красными рамками) итальянских стенных ящериц из новой популяции, размножившейся на острове Pod Mrcaru от ящериц, переселенных с острова Pod Kopiste.
8. В диете резко возросла доля растительной пищи в весенне-летний период - 34% весной и 61% летом против 7% весной и 4% летом в предковой популяции (Рис. 15):
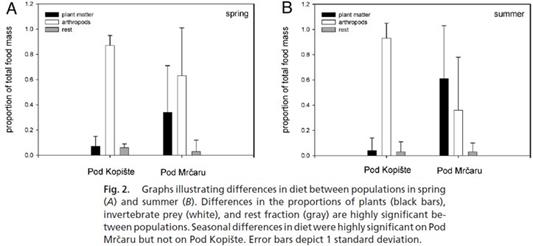
Рисунок 1 5 . Резкие изменения в диете итальянских стенных ящериц новой и старой популяций. Черными столбиками обозначена доля растительной пищи в рационе ящериц, белыми столбиками – членистоногие. Рисунок взят из работы ( Herrel et al ., 2008).
9. И наконец, самое удивительное, у всех препарированных ящериц новой популяции в пищеварительной системе обнаружилось нечто вроде нового органа (!) «cecal valve» (цекальный клапан), который имеет вид складки (перегородки) на внутренней стороне кишечника. Это образование задерживает прохождение пищи через кишечник и образует некие «ферментативные палаты», где целлюлоза преобразуется в более удобоваримые вещи. Таким образом, это новоприобретение улучшает усвоение растительной пищи.
Подобные морфологические образования имеются у некоторых растительноядных ящериц из этого семейства (Lacertidae) и из других семейств ящериц (агамовые, игуановые). Однако никогда до этого не регистрировались у обсуждаемых итальянских стенных ящериц (Podarcis sicula) или у ящериц P. melisellensis. В целом, это морфологическое образование имеется менее чем у 1% всех существующих ящериц и змей.
Таким образом, исследователи столкнулись не только с количественными морфологическими изменениями, произошедшими в новой популяции, но и с качественными, выразившимися в появлении новых морфологических структур!
10. Более того, в кишечнике ящериц новой популяции обнаружились нематоды (круглые черви). Авторы работы посчитали нематод индикаторами присутствия в пищеварительной системе ящериц симбиотических микроорганизмов, перерабатывающих целлюлозу из растительной пищи. Поскольку у ящериц исходной популяции (на островке Pod Kopiste) нематоды не были найдены, то авторы предположили, что симбиотические микроорганизмы появились в новой популяции, вероятно, тоже вместе с новым органом, и (или) в связи с резким изменением пищевого рациона в сторону растительноядности.
Как видим, список произошедших изменений весьма впечатляет. Во-первых, произошли множественные количественные морфологические изменения, а во-вторых, еще и качественные (морфологическое новообразование в пищеварительной системе). И весь этот «букет» изменений случился с новой популяцией всего за 36 лет (авторы пишут примерно о 30 поколениях).
Первые четыре пункта изменений авторы связали с разной степенью хищничества на двух островках (Vervust et al., 2007). А именно: 1) более короткие ноги ящериц в новой популяции, 2) меньшую скорость бега, 3) ослабленную реакцию на хищников, и 4) большой размер тела - авторы связали с тем, что ящерицы, видимо, «расслабились» из-за меньшего хищнического давления. Авторы установили, что на новом островке искусственные модели ящериц реже атакуются птицами, чем на исходном острове (возможно, из-за лучшего растительного покрова). Поэтому (по предположению авторов) ящерицы и «расслабились» - по механизму, который я уже описывал выше (см. раздел «ящерицы и недоразумения»). В присутствии хищников самые медлительные ящерицы больше выедаются, поэтому популяция ящериц поддерживается в «спортивной форме». А при ослабленном хищническом давлении начинает выживать большее число ящериц. В том числе, и более медлительные, коротконогие и т.п. В результате, средние показатели озвученных признаков сдвигаются в соответствующую сторону.
Таким образом, первые четыре пункта изменений в ящеричной популяции вряд ли можно отнести к разряду эволюционных изменений вообще, и уж тем более, к эволюции под действием естественного отбора, в частности. Если уж нам очень хочется заявить именно об эволюции, то это пример антидарвиновской эволюции - когда сдвиг нормы признака произошёл из-за ослабления естественного отбора. И затем (предположим) этот «эволюционный сдвиг» закрепился в популяции чисто случайным образом, в результате банального дрейфа генов. Например, более резвые и длинногие ящерицы просто сдохли по разным случайным причинам (в этой маленькой популяции), и остались только медлительные и коротконогие ящерицы. То есть, в популяции произошли некоторые наследственные изменения, но естественный отбор к этим изменениям не имел никакого отношения. А имела место чистая случайность.
Хотя (еще раз) озвученные изменения, возможно, вообще не наследственные, а являются просто результатом прекращения избирательного выедания хищниками (см. выше).
Однако другая часть новых и изменившихся признаков вряд ли появилась вследствие ослабления естественного отбора. Давайте еще раз посмотрим на эти признаки:
1. Головы ящериц в новой популяции оказались больше в длину, ширину и высоту (относительно размера тела), чем головы ящериц с исходного островка.
2. Сила укуса новых ящериц оказалась выше (у самцов на 14%, у самок на 34%).
3. В диете резко возросла доля растительной пищи в весенне-летний период - 34% весной и 61% летом против 7% весной и 4% летом в предковой популяции.
4. В пищеварительной системе возникло новое морфологическое образование – цекальный клапан, замедляющий прохождение пищи по пищеводу, и дающий возможность кишечным микроорганизмам лучше переработать целлюлозу (из растительной пищи).
5. Наконец, авторы предположили, что ящерицы новой популяции, возможно, отказались и от территориального поведения (больше не защищают собственные участки), так как плотность ящериц в новой популяции оказалась намного выше, чем в старой популяции.
Авторы работы связали все эти изменения в единый адаптивный комплекс. Например, большие размеры головы позволяют новым ящерицам развивать более сильный укус. Что в свою очередь, облегчает ящерицам процесс откусывания прочных кусочков растений (листьев, стеблей). Одновременно, новый морфологический орган в кишечнике (цекальный клапан) помогает ящерицам лучше усваивать растительную пищу. И всё это вместе позволило ящерицам резко повысить долю растительной пищи в собственном рационе. И наконец, авторы предположили, что поскольку растительный корм - это более «предсказуемый» и доступный ресурс, чем насекомые, то ящерицы, ставшие в значительной степени растительноядными, отказались и от территориального поведения.
Вопрос, возникли ли (только что перечисленные) изменения именно по механизму естественного отбора в данном исследовании:
1) Вообще не изучался.
2) Более того, подобный механизм «эволюции» здесь вообще крайне маловероятен.
Во-первых, осталось неизвестным, повысили ли произошедшие изменения общую приспособленность ящериц на новом островке? Или наоборот, понизили? Или приспособленность не изменилась? Эти вопросы повисли в воздухе, так как сравнение приспособленности «старых» и «новых» ящериц в условиях нового островка вообще не проводилось (в рамках данного исследования).
Действительно, насекомые - это более ценный пищевой ресурс, чем растения (листья и стебли). Поэтому выгода «эволюционного перехода» ящериц новой популяции в сторону растительноядности - остаётся под большим вопросом, но к сожалению, в рамках обсуждаемого исследования этот вопрос вообще не изучался.
Более того. Если уклон в сторону растительной пищи - действительно выгоден, то почему подобный переход не случился (уже давным-давно) на исходном островке, где «испокон веков» обитала исходная популяция итальянских стенных ящериц? Это действительно странно. Мы видим, что уклон к растительноядности развился в новой популяции ящериц всего за 30 (!) поколений... но в то же время, на соседнем островке, с аналогичными условиями среды, такой переход не осуществился ни за 30 поколений, ни за 300 поколений, ни даже за 3000 поколений... Если растительноядной ящерицей быть выгодней, чем насекомоядной, то почему итальянские стенные ящерицы - не стали растительноядными еще миллион лет назад?
Понятно, что вопрос о выгоде (или об отсутствии выгоды) подобного перехода является крайне актуальным. Ведь если никакой выгоды от уклона в растительноядность, на самом деле, нет, тогда мы не имеем никакого права говорить, что произошедшие в ящерицах изменения - это результат именно естественного отбора (более приспособленных). К сожалению, нам остаётся только гадать на эту тему, поскольку, как уже говорилось выше, сравнение приспособленности между ящерицами с разными пищевыми стратегиями в условиях нового островка не проводилось.
Когда мы обсуждали «эволюцию ракушек», мы видели, что во многих таких работах разница в приспособленности раковин разной морфологии прямо исследовалась авторами. Причем как в эксперименте, так и в природе. А именно, выяснялась разница в успешности преодоления раковин (разной морфологии) хищными крабами. В случае же «эволюции итальянских ящериц» - нам остаётся только гадать на эту тему.
Поэтому Вам придётся сообщить (вашему дарвинисту) эту новость примерно таким образом:
- С прискорбием вынуждены сообщить Вам, что данный пример «морфологической эволюции ящериц под действием естественного отбора» не может считаться строго установленным примером «морфологической эволюции ящериц под действием естественного отбора», потому что «действие естественного отбора» в этом примере вообще не изучалось.
На этом дискуссию по этому «строго установленному примеру» Вам следует прекратить (я имею в виду, дискуссию с верующим дарвинистом). Потому что по теме беседы (реальные примеры эволюции в ходе естественного отбора) здесь обсуждать больше нечего.
Для наседающего на Вас верующего дарвиниста этого будет достаточно. Но не для нас. Потому что сам по себе этот пример слишком интересный, чтобы бросать его рассмотрение, едва начав.
Дальше я озвучу лишь возможные (гипотетические) варианты, поскольку, как я уже только что сказал, ничего более строгого (строго установленного) в отношении возможных причин эволюционных изменений из этого исследования не вытянуть. Но попытаться разобраться с этим примером всё-таки надо. Хотя бы в рамках теоретических предположений.
В первую очередь, здесь следует подождать независимых исследований, которые, может быть, прояснят ситуацию. Например, возможно, морфологическое новообразование, обнаруженное исследователями в пищеварительной системе ящериц новой популяции, на самом деле, не является новым для этого вида, а всё-таки иногда встречается у отдельных его представителей (пусть и очень редко)? Тогда это будет не «эволюция нового органа», а просто закрепление уже имеющегося (у данного вида) редкого признака в отдельно взятой популяции. Например, за счет всё того же дрейфа генов.
Не исключены и просто какие-то ошибки. Поэтому хорошо было бы проверить это удивительное исследование на воспроизводимость результатов - путем исследования этих же ящериц независимой группой исследователей.
Кроме того, здесь можно попытаться порассуждать, оставаясь в рамках общепринятой теории эволюции, но с помощью более убедительных теоретических сценариев. Поскольку, как я уже говорил выше, польза от уклона ящериц в растительноядность находится под большим вопросом (хотя бы просто потому, что ящерицы на соседнем островке не спешат становиться растительноядными), то даже рассуждая в рамках принятой теории эволюции, здесь следует предположить более сложные сценарии развития событий, чем просто естественный отбор.
Например, можно предположить такой сценарий:
а) Сначала в результате дрейфа генов ящерицы перестали быть территориальными. То есть, ген (или гены), отвечающие за территориальное поведение - просто выпали или «сломались» у одной из ящериц, завезённых на остров. И затем в силу чисто случайных причин потомки именно этой ящерицы расплодились на острове.
б) В результате, плотность новой популяции ящериц резко возросла (авторы исследования установили такой факт - серьезное повышение плотности ящериц на новом острове).
в) Расплодившимся не территориальным ящерицам перестало хватать привычных пищевых ресурсов в виде насекомых.
г) Вот поэтому (в таких условиях) начался естественный отбор этих всеядных ящериц (но с преобладанием в рационе животной пищи) в сторону преобладания растительной пищи. А именно, любые изменения, позволяющие использовать растительную пищу более эффективно, стали полезными в новых условиях. И таким образом, в данной популяции закрепились и более крупные головы ящериц, способные развить более серьезное усилие при укусе (откусывании кусочков растительности). И пищевые предпочтения ящериц тоже изменились. И даже новое морфологическое образование (цекальный клапан) в пищеварительном тракте появилось, и быстро закрепилось в популяции за счет преимущественного выживания особей именно с таким признаком. И нужные симбионты тоже не замедлили поселиться в пищеварительной системе «новых» ящериц.
Можно предположить и другой, полностью аналогичный сценарий развития эволюционных событий, но только уже с другой стартовой причиной:
а) Сначала, из-за случайного дрейфа генов, на новом островке остались только такие ящерицы, которые предпочитали есть растительную пищу в значительно большем количестве, чем это было характерно для ящериц в старой популяции, даже несмотря на сравнительную бесполезность такого поведения.
б) В результате, начался естественный отбор в данном направлении. Любые изменения, позволяющие использовать растительную пищу более эффективно, стали полезными в новых условиях. И таким образом, в данной популяции закрепились и более крупные головы ящериц, способные развить более серьезное усилие при укусе (откусывании кусочков растений). И новое морфологическое образование в пищеварительном тракте быстро закрепилось в популяции, и нужные симбионты тоже поселились в пищеварительной системе «новых» ящериц.
в) На каком-то из 30 поколений ящерицы чисто случайным образом (снова за счет дрейфа генов) потеряли тягу и к территориальному поведению.
Наверное, можно предположить и какие-то другие гипотетические сценарии произошедших событий, задействуя тот же механизм: «дрейф генов + новое направление естественного отбора». Но к сожалению, всё это будет лишь гаданием на кофейной гуще. Как я уже говорил, мы не знаем (и никогда уже не узнаем), по какому именно сценарию развивались реальные события на этом островке.
Однако даже предложенные сценарии всё равно являются маловероятными. Маловероятными из-за чрезвычайно высокой скорости произошедших изменений (слишком высокой для механизма «случайные мутации + естественный отбор»).
Действительно, авторы исследования констатировали целый «букет» новых признаков, которые, весьма похоже, биологически согласованы друг с другом. И на появление и отбор всего этого «букета» было отпущено всего 30 поколений. Такая, воистину, сказочная скорость изменений меньше всего напоминает постулированный механизм эволюции («случайные мутации + естественный отбор»).
Если мы постулируем случайность появления соответствующих признаков, а потом еще и некоторый естественный отбор в ряду поколений, приводящий к закреплению этих признаков, то 30 поколений ящериц - неправдоподобно мало для такого механизма. Особенно если учесть, что и сам материал для эволюции, в данном случае, тоже был минимальным: 1) всего пять пар исходных ящериц в самом начале, и 2) небольшая популяция ящериц в самом конце. В связи с этим выглядит весьма странным, что из этого мизерного материала всего за 30 поколений как-будто по заказу «вырисовался» целый букет новых морфологических и поведенческих признаков: 1) более «мирные» пищевые предпочтения ящериц (в сторону растительноядности); 2) относительно крупные размеры головы (и как следствие, повышенная сила укуса); 3) появление нового морфологического образования в пищеварительной системе. Очевидно, что в рамках парадигмы «новые признаки - это случайные мутации» мы здесь имеем дело:
1. Либо с невероятным везением. То есть, исследователям крайне повезло натолкнуться на случай невероятного стечения обстоятельств.
2. Или с эффектом телепатической силы исследователей, чья вера в «теорию эволюции» так повлияла на объективную реальность, что сделала невероятное возможным. Например, сумела повлиять на опубликованные результаты.
3. Или это указывает на то, что «новые признаки», на самом деле, совсем не новые, а уже присутствовали у особей, выпущенных на остров, пусть даже и в скрытом виде (см. ниже).
4. Наконец, это может указывать на то, что «случайные мутации + естественный отбор», на самом деле, совсем не являются механизмами реальных эволюционных изменений. А эволюция происходит под действием каких-то других биологических законов. Или даже вообще не биологических законов, а каких-то особых сил, благодаря которым эволюция может происходить буквально по волшебству и в крайне сжатые сроки.
Но и это еще не всё.
Допустим, исследователи итальянских ящериц, действительно, натолкнулись именно на комплекс невероятных совпадений - в крошечной популяции как по заказу появилась целая «плеяда» новых (нужных) случайных мутаций.
Однако даже если это действительно так, то времени на естественный отбор этих «случайных мутаций» тоже катастрофически не хватает (для их закрепления во всей популяции). Например, допустим, сначала появилась «случайная мутация» в виде появления нового морфологического образования в пищеварительной системе (цекальный клапан). Насколько конкретно этот новый признак повысит приспособленность ящерицы, обладающей этим признаком, в сравнении с ящерицей, которая еще не имеет этого признака? Если сравнительная приспособленность такой ящерицы повысится, допустим, на 10% (а это довольно большая величина, учитывая еще тысячи других причин, из которых складывается общая приспособленность ящериц), тогда этот признак завоюет всю популяцию ящериц только примерно через 150 поколений. Что никак не укладывается в рамки «отпущенных нам» 30 поколений.
А если цекальный клапан появился в тот момент, когда ящерицы были еще преимущественно насекомоядными? В этом случае он вообще окажется нужен?
Другой пример - у «новых» ящериц размеры тела немного увеличились, и дополнительно еще увеличились размеры головы. В результате у самцов сила укуса возросла на 14%, а у самок на 34%. Вряд ли общая приспособленность ящериц от этого могла возрасти на столько же процентов. Ведь в природе достаточно одних только растительных ресурсов, которые вполне способна откусить и ящерица с меньшей силой укуса (и с меньшей головой). Не говоря уже о насекомых. Более того, доля растительной пищи в рационе «новых» ящериц составляет примерно половину от общей массы пищи (34% весной и 61% летом). В связи с этим, нам придётся резко уменьшить роль силы укуса на общую выживаемость ящериц. Но даже если мы сократим этот показатель всего втрое (от установленного увеличения силы укуса), то получим увеличение общей приспособленности у самцов на 5%, а у самок на 11%. Признак, увеличивающий общую приспособленность ящерицы на такую величину, будет «завоёвывать» популяцию примерно за 150 поколений для самок, и примерно за 300 поколений для самцов. Возникает вопрос, как же тогда этот признак успел завоевать популяцию всего за 30 поколений? И это еще в том случае, если данный признак появился уже среди первых поколений ящериц, а не где-нибудь на «полпути», или вообще в конце (например, за 5 лет до приезда исследователей).
Кроме того, предположение об увеличении общей выживаемости ящериц на 5% из-за увеличения силы их укуса на 14% - это предположение вряд ли правдоподобно. Реальное повышение общей приспособленности (в условиях реальной растительности) здесь может составить всего доли процента (если вообще будет заметно). Напомню, что признак, гипотетически повышающий общую приспособленность особи на 1%, будет завоевывать популяцию примерно 1000 поколений. Если вообще закрепится - в маленьких популяциях роль дрейфа генов очень серьезна и новые признаки (даже будучи полезными), едва появившись, вполне могут «вылетать» из популяций по чисто случайным причинам, пока доля носителей этих признаков в популяции еще мала.
Можно, конечно, предположить, что увеличение силы укуса на 14-34% могло улучшить питание ящериц не слабо, а наоборот, резко. Например, сделав доступными те растительные корма, которые раньше были вообще недоступны этим ящерицам. Но тогда (еще раз) почему такое, столь полезное изменение давным-давно не закрепилось в других популяциях итальянских стенных ящериц?
В общем, чисто теоретически, озвученная версия «дрейф генов + случайные мутации + естественный отбор = букет эволюционных изменений за 30 поколений в маленькой популяции» - не является принципиально невозможной. Так же как и самопроизвольное залезание газа, выпущенного из баллона, обратно в баллон - это тоже теоретически возможный процесс, просто очень маловероятный. Так же и здесь, эволюция за 30 поколений по механизму «дрейф генов + случайные мутации + естественный отбор» в обсуждаемом случае тоже, в принципе, возможна, хотя и весьма маловероятна.
Об этом Вы тоже можете сообщить тому верующему дарвинисту, с которым беседуете в текущий момент - приведенный пример с «чудо-ящерицами» не проходит критерий строго установленного по предлагаемому механизму изменений. Потому что, во-первых, эти механизмы в данном исследовании вообще не изучались, а во-вторых, скорость всех произошедших изменений не очень вписывается в постулируемый механизм «случайные мутации + естественный отбор».
Ну а теперь давайте рассмотрим более правдоподобные версии возможных причин, которые могли бы привести к установленным изменениям за столь короткий срок. Учтите, что эти гипотетические версии верующему дарвинисту озвучивать не надо. Дело в том, что у глубоко верующих дарвинистов часто происходит некая интеллектуальная редукция, механизм возникновения которой пока неясен. В результате этой редукции, дарвинист вообще перестаёт воспринимать какие-либо альтернативные сценарии изменения видов, кроме единственно верного учения о естественном отборе. Который он и проповедует аудитории, даже если альтернативные сценарии прямо-таки напрашиваются. Поэтому беседовать с верующим дарвинистом на уровне возможных гипотез - практически бесполезное занятие. Общение с верующим дарвинистом следует проводить исключительно на языке строгих эмпирических фактов, которые, как известно, печальны для дарвинизма. Попросту говоря, их нет вообще (строгих фактов в пользу этой теории).
Однако нам же самим всё-таки надо попытаться разобраться, что же там случилось с этими ящерицами на самом деле? Поэтому давайте теперь перечислим наиболее правдоподобные версии. Начнем с самой простой:
Возможно, все перечисленные выше «эволюционные изменения» - это на самом деле, просто реализация внутривидовой изменчивости, которая уже имелась у данного вида. Действительно, итальянская стенная ящерица (Podarcis sicula) - весьма полиморфный (морфологически изменчивый) биологический вид. В рамках этого вида выделены десятки подвидов.[31] Так что вполне возможно, многие из перечисленных выше «эволюционных изменений», на самом деле, не выходят за рамки наблюдавшихся прежде значений, характерных для этого вида, а встречаются и в других популяциях этих же ящериц, хотя бы у отдельных представителей. Почти наверняка среди разных популяций итальянских стенных ящериц можно найти индивидуумов столь же «головастых», как и ящерицы в обсуждаемой новой популяции. Вероятно, можно найти и особей, предпочитающих растительную пищу, и даже особей, у которых имеется (в качестве редкой морфологической особенности) пресловутый цекальный клапан в пищеварительном тракте. Авторы работы отмечают, что данный орган, до этого, не был найден у каких-либо представителей этого вида. Но может быть, теперь стоит поискать тщательней? Тем более что этот орган, можно сказать, «рассыпан» по многим «ящеричным» таксонам, встречаясь у разных видов ящериц из семейств агамовых, игуановых и настоящих ящериц. Так может быть, этот орган (в том числе) затерялся и в каких-нибудь популяциях итальянской стенной ящерицы, хотя бы у отдельных представителей этого вида?
Действительно, вряд ли следует признать экономичным предположение, что этот орган возник у итальянской стенной ящерицы совершенно заново, в ходе чисто случайной эволюции... если такой же орган уже имеется у некоторых других представителей из этого же семейства (настоящие ящерицы, Lacertidae).
Впрочем, лично я не знаком с морфологическим разнообразием итальянских стенных ящериц. И не настаиваю именно на версии внутривидовой изменчивости. Тем более что даже если все перечисленные признаки, действительно, встречаются в разных популяциях итальянских стенных ящериц, то все равно непонятно, каким образом все эти признаки вдруг очутились в новой популяции ящериц, если конкретные родоначальники этой популяции не имели данных признаков.
Хотя примерно такая же трудность встает и перед версией «случайные мутации + естественный отбор». Как я уже говорил выше, весьма неправдоподобно, что все необходимые новые признаки, как по заказу (но чисто случайно) возникли всего за 30 поколений в изолированной маленькой популяции ящериц.
Вторая версия - гораздо более интригующая, и возможно, недалека от истинной картины. Здесь необходимо вспомнить о так называемой скрытой изменчивости.
Для тех, кто мало знаком с этим термином - поясняю. Внутри рамок любого биологического вида, в природных условиях, мы практически всегда наблюдаем ту или иную внутривидовую изменчивость (индивидуальную и между разными популяциями одного вида). Условно назовем эту внутривидовую изменчивость «наблюдаемой изменчивостью». Каждый биологический вид обладает тем или иным запасом «наблюдаемой изменчивости». Причем мы наблюдаем, что у одних видов этот запас весьма впечатляющ (так называемые полиморфные виды), а у других видов запас «наблюдаемой изменчивости» существенно ниже.
Вполне логично предположить, что какая-то часть от этого (имеющегося) запаса внутривидовой изменчивости - вообще скрыта от глаз наблюдателя, то есть, вообще не проявляется в фенотипах особей (того или иного биологического вида) в известных природных условиях. Однако эта скрытая изменчивость, в принципе, может проявиться. Например, при расщеплении полиморфных генов. Или (еще интересней) в каких-нибудь новых условиях окружающей среды, которые и «включают» соответствующие «спящие» гены (о существовании которых мы ранее не подозревали), и мы начинаем наблюдать якобы «новые» признаки.
В качестве проявления скрытой изменчивости (того или иного биологического вида), вероятно, можно рассматривать и результаты искусственного отбора. Действительно, многие морфологические признаки, которые мы наблюдаем в разных породах собак, кошек, голубей, кур и т.п. - никогда не проявляются в диких фенотипах, но могут быть «обнажены» в ходе искусственного отбора.
Впрочем, плоды искусственного отбора можно и не считать проявлением скрытой изменчивости (того биологического вида, с которым проводилась селекция), а считать эти плоды результатом кропотливого труда по нахождению и отбору соответствующих новых мутаций, новых сочетаний аллелей (в том числе и мутантных) и т.п. Однако в этом случае становится не совсем понятно, почему эти «плоды искусственного отбора» столь сильно различаются по своему масштабу, в зависимости от того, с каким конкретно видом производилась селекция.
Достаточно сравнить результаты искусственного отбора уже упомянутых собак с результатами искусственного отбора, например, ослов, верблюдов, лам, яков или гусей... И мы начинаем понимать, что далеко не над всеми видами можно так эффектно «поиздеваться», как над собаками. Можно, конечно, говорить, что это просто так получилось, что селекционеры над гусями, в целом, трудились меньше, чем, например, над курами (или голубями), и поэтому такие скромные результаты. Однако большинство имеющихся свидетельств этого не подтверждают. Например, с одной стороны, известно, что многие слабо изменившиеся одомашенные животные имеют очень древнюю историю одомашнивания. Тем не менее, результаты селекции не впечатляют (ослы, верблюды, ламы, альпаки, гуси и др.). С другой стороны, породы собак очень маленького размера известны уже из захоронений 3 - 1.5 тыс. лет до нашей эры. Причем в обеих частях Света (например, в египетских и древне-индейских захоронениях). Получается, что уже древние индейцы сумели вывести маленьких собачек, но при этом в отношении лам или альпак (которые использовались и продолжают использоваться в сельском хозяйстве активно, всесторонне и уже очень давно) почти не преуспели. Но почему? Потому ли, что они просто мало трудились конкретно над ламами и альпаками, а над собаками трудились много? Или потому, что собака, как вид, имеет впечатляющий размах возможной внутривидовой изменчивости, и поэтому она его и демонстрирует (весьма щедро, причем уже с древнейших времен), а у лам или альпак - запас внутривидовой изменчивости гораздо скромнее?
В общем, анализ результатов селекции разных одомашненных видов как бы намекает на возможность существования неких границ изменчивости (в том числе, и границ скрытой изменчивости), которые уникальны для каждого вида.
Правда, некоторые генетики, на основании каких-то там своих генетических фактов, утверждают, что они не видят никаких «границ изменчивости вида» на генетическом уровне, и что внутривидовая изменчивость на генетическом уровне ничем принципиальным не отличается от изменчивости между видами (нет никаких «генетических скачков» на этом уровне). Однако морфологические факты – тоже упрямая вещь. И наверное, эти факты не менее упрямы, чем генетические?
Чтобы попытаться как-то совместить всё это, можно предположить, что в геномах биологических видов присутствует некоторая вариативная часть генома, которая может изменяться самым широким образом. Но одновременно, имеется и некая неизменная часть генома, строго хранящая именно видовые признаки, и не подверженная изменениям. В этом случае мы будем наблюдать именно такую картину с искусственным отбором, какую и наблюдаем в реальности.
Следует отметить, что именно такую идею высказывал российский генетик Юрий Петрович Алтухов (о генетическом мономорфизме некоторых видовых признаков), называя неизменную часть генома биологического вида – мономорфной частью генома (Алтухов, Рычков, 1972).
Теперь если предположить, что эта мономорфная часть, отвечающая за видовые признаки, занимает в геноме очень небольшой объем (а скорее всего, так и есть), то вполне может получиться, что генетики её действительно не видят. Таким образом, мы спекулятивно (но эффективно) снимаем противоречие между мнением генетиков (которые не видят никаких «границ изменчивости» в генотипах) и фактами из области морфологии, которые серьезно свидетельствуют в пользу существования таких границ.
Прямо сейчас я не буду пытаться решить, какие из озвученных выше предположений верны. Или же «истина где-то посередине». Но позже мы еще вернёмся к «мономорфизму биологических видов». Ниже я представлю несколько конкретных примеров такого «мономорфизма видов», которых, наверное, уже достаточно, чтобы признать принятую сегодня теорию эволюции опровергнутой. А сейчас давайте пока вернемся обратно к «эволюционирующим ящерицам».
Итак, можно предположить, что появление того же цекального клапана в пищеварительной системе ящериц новой популяции - это, на самом деле, не новообразование, а реализация скрытой изменчивости, которая закрепилась в данной популяции. То есть, гены, ответственные за появление этого морфологического признака - уже имелись в генофонде этого биологического вида, и могут время от времени проявляться у отдельных его представителей, под действием тех или иных причин, или вообще случайно. Теоретически, они могут проявиться и закрепиться в новой популяции. Данная версия является не более спекулятивной, чем случайное появление целого «букета» новых морфологических признаков в маленькой популяции, и их успешный отбор всего за 30 поколений.
СКРЫТНЫЕ МУРАВЬИ
Интересно, что мы уже располагаем и другими установленными примерами проявления скрытой изменчивости у разных биологических видов.
Например, в работе (Rajakumar et al., 2012) исследователи обнаружили, что сразу у нескольких видов муравьев из обширного рода Pheidole, потенциально имеется (в скрытом виде) особая каста «сверхсолдат». То есть:
1) У нескольких видов муравьев из этого рода - каста «сверхсолдат» присутствует постоянно.
2) У некоторых видов муравьев из этого же рода - каста «сверхсолдат» присутствует лишь в виде редких аномалий. То есть, у отдельных, редко появляющихся особей, напоминающих по виду «сверхсолдат».
3) И наконец, у некоторых видов муравьев этого рода - каста «сверхсолдат» вообще никогда не проявляется (даже в виде аномалии)... но биологам в данной работе удалось создать касту «сверхсолдат» и у этих видов тоже, обработав личинки специальным образом.
Получается, что способность к образованию касты «сверхсолдат» уже имеется в скрытом виде, по крайней мере, у некоторого числа видов муравьев рода Pheidole. То есть, это именно то, о чем мы только что говорили - та самая скрытая изменчивость.
Вот что написал по этому поводу в соответствующей научно-популярной заметке уже хорошо известный нам автор (Марков, 2012б):
...Опираясь на полученные данные, исследователи предположили, что потенциальная возможность формирования сверхсолдат была унаследована муравьями рода Pheidole от общего предка, жившего 35–60 млн лет назад. Реализовали же ее только те виды, которым это почему-то оказалось выгодно (например, из-за жизни в местах, где водятся кочевые муравьи). У других видов эта возможность сохранилась в скрытом состоянии. В таком случае следует ожидать, что из личинок тех видов, у которых нет касты сверхсолдат, можно, подобрав правильные условия, вырастить нечто на них похожее.
Известно, что выбор личинкой муравья того или иного варианта развития зависит от уровня ювенильного гормона … Поэтому логично предположить, что при помощи этого гормона можно «включить» и скрытую программу развития сверхсолдат у видов, не имеющих этой касты. Эксперименты на трех видах Pheidole, не имеющих сверхсолдат (P. spadonia, P. morrisi, P. hyatti), подтвердили эту гипотезу. Оказалось, что если взять личинку любого из этих видов, собравшуюся превратиться в обычного солдата, и намазать ей брюшко метопреном - аналогом ювенильного гормона, - то личинка ускоряет свой рост, обзаводится двумя парами крыловых дисков с высокой экспрессией гена sal и в итоге становится сверхсолдатом.
Если теперь выбросить из первого абзаца этой цитаты все лишние теоретические разглагольствования про «эволюцию», «общего предка» и «биологическую выгоду» (заменив их более строгими формулировками), то первый абзац можно переписать таким образом:
...потенциальная возможность формирования сверхсолдат [имеется] у муравьев рода Pheidole. Реали[зуют] же ее только [некоторые] виды этого рода. У других видов эта возможность [имеется] в скрытом состоянии. В таком случае следует ожидать, что из личинок тех видов, у которых нет касты сверхсолдат, можно, подобрав правильные условия, вырастить нечто на них похожее.
Точно так же можно предположить, что и обсуждаемый нами цекальный клапан в пищеварительной системе «новых» итальянских стенных ящериц - это, на самом деле, никакое не эволюционное новшество, а просто «вынырнувший» скрытый признак, уже имеющийся в геноме данного вида. Тем более что такие же клапаны уже имеются у разных представителей разных семейств ящериц. В том числе, и у некоторых представителей семейства настоящих ящериц (Lacertidae), к которому принадлежит и обсуждаемая нами стенная ящерица. То есть, аналогия с муравьями и кастой «сверхсолдат» здесь практически полная.
К этой гипотетической эволюционной схеме («эволюция» как реализация скрытой изменчивости) можно «пристегнуть» (при большом желании) еще и другие теоретические спекуляции. Например, «естественный отбор» или даже «эпигенетическую теорию эволюции». Вот как это было сделано в уже упомянутой заметке про муравьев и сверхсолдат (Марков, 2012б):
...Как уже говорилось, у некоторых видов, не имеющих касты сверхсолдат (включая P. morrisi), такие особи иногда появляются в качестве редкой аномалии развития («морфоза»). Это позволило авторам предположить, что независимое появление касты сверхсолдат в разных эволюционных линиях муравьев рода Pheidole происходило благодаря механизму, известному как «генетическая ассимиляция морфозов» (см. также: Genetic assimilation). Суть идеи в следующем. Если какой-нибудь редкий морфоз (то есть аномалия развития, иногда возникающая в ответ на какие-то внешние воздействия) вдруг окажется полезным, отбор начнет закреплять в генофонде популяции такие мутации, которые повышают вероятность реализации этого морфоза. В итоге бывшая аномалия постепенно станет нормой, то есть «впишется» (ассимилируется) в генетическую программу развития. Абсолютизация этого эволюционного механизма лежит в основе так называемой «эпигенетической теории эволюции».
Однако еще проще было бы подредактировать эту идею таким образом – «эволюционные изменения таксона» - это, на самом деле, просто реализация скрытых возможностей и признаков, уже заложенных в данном таксоне. Реализация, вызванная соответствующими требованиями среды, или даже без всяких «требований» (например, из-за чисто случайного дрейфа). Геном исходного биологического таксона (с множеством заложенных возможностей и вариантов развития признаков) просто распадается на несколько разных, более «узких» вариантов генома, которые уже имелись внутри исходного (предкового) генома. Понятно, что в этом случае правильнее будет говорить не об «эволюции», а о «псевдо-эволюции», которая не производит, а лишь имитирует появление эволюционных изменений. Такая «псевдоэволюция» может идти только по механизму распада геномов с изначально большими потенциальными возможностями, в сторону всё более «узких» и «конкретных» вариантов геномов, со всё меньшими способностями к демонстрации «новых признаков».
На всякий случай отмечу, что похожий взгляд на глобальную эволюцию животных выдвинул генетик М. Шерман в своей концепции «универсального генома» (Sherman, 2007), где вся эволюция животных представляется, скорее, распадом некоего исходного «универсального генома», чем появлением чего-нибудь принципиально нового.
Подобные взгляды на эволюцию сходны и с известной «бараминной концепцией» креационистов, где многочисленные биологические виды весьма быстро появляются (распадаются?) из неких исходно созданных «бараминов».
Кроме того, в рамках идеи «псевдоэволюции по модели распада геномов», вполне допустимо ввести еще одну теоретическую спекуляцию - что возможен не только распад «широких» (предковых) геномов на всё более «узкие», но и простой «дрейф» некоторых признаков в разных направлениях тоже возможен. Если в исходном генотипе имеется (запрограммирована) сама возможность некоторой вариабельности того или иного признака, то и в природе следует ожидать быстрого появления таких вариаций.
Допустим, в исходном генотипе (гипотетического) общего предка всех птиц-шалашников была заложена возможность вариабельности формы построения и украшения брачных шалашей. Тогда и в природе следует ожидать сравнительно быстрого «распада» птиц-шалашников по этим признакам - по разнообразию формы брачных шалашей и способов их украшения. А вот сама способность и потребность строить брачные шалаши (той или иной формы) остаётся неизменной у разных представителей этого таксона птиц. Несмотря на то, что эти сооружения не имеют никакого отношения к гнездам, а служат только брачными шалашами в прямом смысле этого слова.[32] То есть, с позиций чисто биологической целесообразности, эти брачные беседки совершенно не нужны - тысячи видов птиц прекрасно спариваются без всяких брачных беседок. Но поскольку именно такая (романтичная) особенность брачного поведения запрограммирована в генах конкретно этого таксона птиц, то птицы её и демонстрируют. Причем эта романтичная особенность включает еще и элементы самой настоящей эстетики – самцы украшают свои шалаши в соответствии с определенными эстетическими правилами.
КСТАТИ О ПТИЦАХ
Со стороны этологов имеются попытки объяснить целесообразность романтических беседок этих птиц тем, что спаривание у представителей этого биологического таксона требует долгой «настройки». Иначе спаривание чревато физическими или психическими травмами для самки.
Но такое объяснение смотрится чрезвычайно странно (если не сказать, смешно) в рамках «теории естественного отбора». Ведь в рамках этой теории, естественный отбор должен был сам «настроить» самцам и самкам такое спаривание, которое бы их не травмировало. И наверное, настроить каким-нибудь более прямым способом, чем через сооружение специальных брачных беседок, с украшением этих беседок цветами, ягодами и раковинами, да еще и с соответствующим разделением этих декоративных элементов по разным цветам?
Отметим, что у многих видов птиц, самки, наоборот, постоянно «умоляют» самцов о сексе. А у некоторых птиц самки еще и склонны к супружеской неверности, и стоит только «мужу» отвернуться… Отсюда можно заключить, что сам процесс спаривания явно не приносит самкам этих видов ни физических, ни психических страданий. Скорее уж, наоборот.
А вот конкретно у птиц-шалашников, «естественный отбор самых приспособленных самок» (т.е. самых плодовитых!) почему-то довёл этих бедных птиц до такого состояния, что они вообще не могут спариться без длительного периода деликатных ухаживаний – иначе психическая или физическая травма обеспечена. Таким образом, эти бедные птицы стали жертвой воистину беспрецедентной неисповедимости путей естественного отбора. В результате «эволюции более приспособленных», самки в конечном итоге получились такими «нежными», что обычное спаривание (необходимое для размножения) приносит самкам физические и душевные страдания! Настолько, что эти птицы вынуждены строить брачные шалаши, украшать их цветами и ракушками, и осторожно, постепенно сближаться друг с другом.
То есть, «естественный отбор более приспособленных», кажется, сделал всё, чтобы максимально затруднить этим птицам их размножение.
Давайте представим себе, что человечество объявило всемирный конкурс на самых плодовитых женщин. Я глубоко сомневаюсь, что в этом конкурсе победительницами станут такие женщины, которым каждая физическая близость с мужчиной доставляет физические и психические страдания. Мне кажется, такие женщины никогда не попадут даже в число призёров. А вот у птиц-шалашников (как уверяют нас некоторые дарвинисты), именно такие самки почему-то выиграли все призы.
Я же говорю – пути Естественного Отбора неисповедимы.
Интересно так же, что у шалашников в ходе знакомства самца и самки, самец тоже может совершать удивительные поступки. Например, самец может… прогнать самку от своей брачной беседки. То есть, самец прогоняет ту самую самку, ради привлечения которой (согласно дедушке Дарвину) эта беседка и строилась.[33]
В общем, если говорить серьезно (без иронии), то это как раз тот случай, когда применение «теории естественного отбора» к конкретным биологическим фактам приводит уже к откровенному бреду (или абсурду).
Объяснить подобные факты можно только в рамках представлений о дизайне биологических таксонов, где главным мотивом было создание максимального разнообразия, в том числе, и разнообразия брачного поведения птиц. То есть, здесь мы имеем дело с разнообразием ради самого разнообразия, без какой-либо биологической целесообразности.
ОБРАТНО К ЯЩЕРИЦАМ
Возвращаясь опять к нашим ящерицам и к предполагаемой у них скрытой изменчивости, мы сталкиваемся с некоторыми трудностями. Допустим, новое морфологическое образование (цекальный клапан) было уже скрыто в геноме данных ящериц, а потом проявилось в новой популяции. Но ведь были зарегистрированы еще и другие морфологические изменения, которые никак не вписываются в те 30 поколений, в которые мы должны втиснуть наши теоретические спекуляции.
В связи с этим, особенно интригующе выглядит возможность, что в геноме ящерицы скрыт не какой-то отдельный признак, а сразу целая группа признаков, связанная между собой в единый адаптивный комплекс, где разные признаки усиливают биологическую целесообразность друг друга. И вся эта группа признаков «включается» одновременно. Или в результате «пробуждения» какого-то единого гена-переключателя, запускающего всю остальную генную сеть. Или же через одновременное включение сразу нескольких генов, отвечающих за разные признаки этого адаптивного комплекса, но в ответ на один и тот же вызов среды. То есть, в геноме ящерицы может быть скрыт целый альтернативный морфотип (комплекс признаков), который может «просыпаться» сразу как единое целое. В этом случае, зарегистрированный факт «быстрых эволюционных изменений» из разряда «невероятного стечения обстоятельств» переходит в разряд вполне объяснимых природных явлений.
Тем более что в живой природе мы имеем огромное число аналогичных примеров - практически всем живым существам, в большей или меньшей степени, свойственна пластичность фенотипов, вызываемая спецификой конкретной среды обитания. Особенно сильно такие способности выражены (наверное) у некоторых растений. Целый ряд видов растений может демонстрировать весьма разные варианты (морфотипы) в зависимости от тех условий, в которые они попали. Иногда растения одного вида в разных условиях вырастают настолько разными, что могут стать почти неузнаваемыми для неспециалиста.
ЖУКИ ПОЛЕТЕЛИ
В связи с вышеозвученным (возможности переключения целых морфотипов), нельзя не привести еще один пример быстрой реализации скрытой изменчивости вида. Пример настолько яркий и наглядный, что лучше уже, наверное, нельзя и придумать.
Эта поучительная история случилась с американским жуком - амброзиевым полосатым листоедом, или зигограммой (Zygogramma suturalis). Этот жук, прибыв к нам из Северной Америки, очень быстро научился... летать!
То есть, до своего «приезда» в Евразию, этот полосатый листоед летать не умел... но после приезда в Евразию по-быстрому научился этому хитрому занятию. Даю слово авторам соответствующей научно-популярной публикации (Голубева, Опарина, 2011):
После первого выпуска зигограммы в 1978 г. на поля Ставрополья прошло всего 5 лет, и нелетающий жук обрел способность к полету, с соответствующими морфологическими изменениями в крыльях, их мускулатуре и ее иннервации. Все группы метаторакальных мышц полностью развиты и состоят из 10–15 толстых, эластичных пучков. За этот период в зоне «популяционной волны» «пешие» жуки полностью замещаются «летунами». Сейчас уже установлено, что все палеоарктические популяции зигограммы — на юге Европы, в Китае, на Дальнем Востоке России (тоже интродуцированные) — как-то «включают» способность летать.
Это удивительно яркий пример. Во-первых, крайне малый срок изменения - 5 лет. Во-вторых, мы видим, что способность к полету, действительно, прямо-таки «включается» у этих жуков в массовом порядке, на огромной территории Евразии. Стоило только жукам попасть в новые условия (или даже просто в новую географическую область), как «крупнейшее эволюционное изменение» - способность к полёту - не замедлило себя ждать.
На фоне столь мощного «эволюционного» изменения (жуки полетели!) обсуждаемый нами пример с несчастными эво-ящерицами (появлением в их пищеварительной системе новой «складки») как-то сразу бледнеет и начинает казаться таким незначительным, что вряд ли даже стоило бы о нем говорить, если бы верующие дарвинисты упорно не приводили в качестве «строго доказанного факта эволюции» именно его. Упорство дарвинистов здесь вполне объяснимо. В ответ на просьбу предъявить доказательства правильности теории естественного отбора, дарвинисты первого и второго уровня (школьный или институтский учебник биологии), как правило, обращаются за информацией в русскоязычную википедию. А там как раз и описаны те самые итальянские чудо-ящерицы в качестве «примера наблюдаемой эволюции». Вот дарвинисты и «выкатывают» этих ящериц раз за разом.[34]
А вот «полетевшие жуки зигограммы» в википедии не описаны. Поэтому верующие дарвинисты (первого и второго уровня) их в пример не приводят.
Дарвинисты же более высокого уровня, может быть, и знают о «полетевших жуках». Но молчат. В качестве «строго доказанного примера морфологической эволюции под действием естественного или полового отбора» - этих жуков не приводят. И это понятно.
Потому что даже самому оголтелому дарвинисту в данном случае ясно, что ни о какой «эволюции под действием естественного отбора» здесь не может быть и речи. Ведь способность к активному полёту - это очень сложная способность, которая возникает только если десятки (или даже сотни) признаков «сольются» вместе в их неразрывном единстве. Начиная с упомянутых авторами (в цитате) «морфологическими изменениями в крыльях, их мускулатуре и иннервации»... и заканчивая специфическим поведением, обеспечивающим правильную технику полета.
Поэтому даже самый невменяемый дарвинист в данном случае понимает, что появление у этих жуков способности летать - это никакое не «эволюционное ново-приобретение», а нечто, уже имеющееся в генофонде этого вида. Которое нужно было просто включить для того, чтобы жуки из нелетающих превратились в летающих.
Об этом пишут и сами авторы заметки, отмечая, что за способность к полету (или за редукцию этой способности) могут отвечать не «случайные мутации», а простые изменения в регуляции уже имеющихся (!) генов в новых условиях (Голубева, Опарина, 2011):
...Можно предположить, что редукция летательного аппарата у жуков чаще бывает вызвана изменениями регуляции экспрессии генов, а не их структурными мутациями или полным исчезновением...
И сразу ниже:
...наш герой - зигограмма - до завоза в Евразию не был замечен в наличии подобной изменчивости.
То есть, судя по описанию авторов - это действительно классическая скрытая изменчивость.
Автор оригинальной работы (Ковалев, 2001), подробно описывая исходную (североамериканскую) форму жука зигограммы, констатирует серьезную редукцию целого ряда признаков, ответственных за полёт:
Структуры груди жуков американских популяций, связанные с полетом: слабая склеротизация задней части нотума, отсутствие системы упора с третьим аксиллярным склеритом; слабое развитие базирадиале, эта структура обнаруживается как склеротизированная пленка, не связанная с радиальной жилкой; второй аксиллярный склерит слабо выражен, не развита проксимальная медиальная пластинка; крылья имеют «пергаментную» структуру.
Крыловая мускулатура третьего грудного сегмента слабо развита, мышечные волокна тонкие и рыхлые. У самцов выражена максимальная редукция групп крыловых мышц: косые спинные мышцы не обнаруживаются в жировой выстилке, тергальные продольные мышцы представлены четырьмя рыхлыми волокнами, дорсовентральные мышцы представлены тремя волоконцами на пластинке жировой ткани; мышцы, прикрепляющиеся к субалярному склериту, представлены одним волокном…
Исходя из такого числа разных признаков, ответственных за полёт, но находящихся у исходных жуков в неработоспособном состоянии, приходится сделать вывод, что способность к активному полету, которую эти жуки дружно приобрели уже через четыре (!) поколения, не могла развиться постепенно, а должна была, действительно, включиться (очень быстро), практически синхронно «приведя в порядок» все перечисленные выше признаки.
Таким образом, мы вынуждены заключить, что в лице жуков зигограмм мы имеем отличный задокументированный факт той самой скрытой изменчивости, о которой мы только что говорили, как о возможной причине видимых «эволюционных изменений». Правда, в случае скрытой изменчивости, более точным будет термин - псевдо-эволюционные изменения. То есть, такие изменения, которые выглядят как эволюционные, но на самом деле, это лишь проявление скрытых потенций, которыми этот вид уже обладает.
Но если с этими жуками листоедами - всё ясно, тогда почему дарвинисты решили, что в случае с «новообразованием в пищеварительной системе итальянских ящериц» - не имеет место точно такое же явление? Тем более что данное «новообразование», на самом деле, отнюдь не ново, а уже «рассыпано» по разным таксонам ящериц, включая, в том числе, и семейство Lacertidae, к которому обсуждаемые ящерицы как раз и принадлежат.
Вот на этой вопросительной ноте, которая превращает данный «строго установленный пример эволюции» - в пример неоднозначный, дискуссионный и «требующий дальнейших исследований»... мы и закончим с чудо-ящерицами острова Монте-Кристо... пардон, Под-Кописте (еще раз пардон, с островка Pod-Mrcaru).
С американским же полосатым жуком листоедом (зигограммой), мы тоже закончим, но только после некоторых завершающих (интересных) штрихов. Помимо того, что мы уже узнали об этом жуке, заслуживает особого внимания еще несколько обстоятельств.
В первую очередь, интересно, что давление естественного отбора здесь опять никем не измерялось. Более того, естественный отбор тут к делу вообще «пришить» не удастся. Потому что изменение способности жуков к полёту случилось всего за пять (!) лет. Причем без всяких «бутылочных горлышек» (резкого сокращения численности жуков), а наоборот, на фоне быстрого наращивания численности в идеальных (для этих жуков) условиях - изобилия кормовой базы, возможного отсутствия ряда «родных» паразитов, заболеваний и т.п. То есть, «переключение» жуков с нелетающих в летающие произошло на фоне отсутствия борьбы за существование (во всяком случае, резкого её ослабления). И это при том, что даже весьма суровый естественный отбор просто физически не способен «заменить» одних жуков на других всего за пять поколений. Да еще и осуществить подобную замену практически синхронно в разных популяциях этих жуков в Евразии (например, в европейских и дальневосточных популяциях).
Очевидно, что естественный отбор, как фактор эволюции, в данном случае вообще не при чём (опять).
Еще интересно, что прибыв в Евразию, наши замечательные жуки зигограммы, видимо, решили – «гулять так гулять»... и помимо мгновенного освоения полёта, начали демонстрировать впечатляющую изменчивость еще и по окраске (Рис. 16). Опять даю слово авторам исходной заметки (Голубева, Опарина, 2011):
В этих специфических условиях зигограмма сильно изменилась. Появились разнообразные варианты окраски покровов, которые не были известны ни в американских популяциях, ни у других видов рода. Более того, коллекционные данные из 11 штатов США говорят об исключительной стабильности окраски жука. В США известны два подвида зигограммы, но они отличаются незначительно и по иным признакам. Большинство появившихся форм были по сравнению с исходной меланистическими, т. е. темноокрашенными, вплоть до полностью черных покровов. Ковалёв пишет, что полная меланизация настолько необычна у видов этого рода, что систематик немедленно выделил бы новый таксон, если бы не наблюдал быстрое появление таких вариаций своими глазами. Всё это необычное разнообразие можно было наблюдать внутри «одиночной волны» на нескольких тысячах гектар на Северном Кавказе.
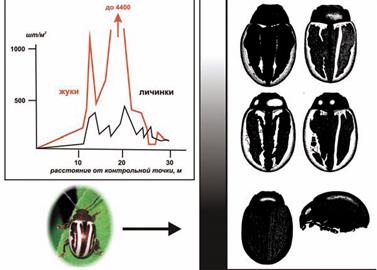
Рисунок 16. Вариации окраски зигограммы — исходный тип и меланисты (из Голубева, Опарина, 2011). Рисунок адаптирован: слева внизу - фотография северо-американской зигограммы с сайта www.texasento.net, Harvard, Worcester Co., Massachusetts, July 18, 2004 (Tom Murray). Структура «уединенной популяционной волны» и фенотипические вариации окраски зигограммы (справа) приводятся из работы О.В. Ковалева (Ковалев, 2001).[35]
Что и говорить, разгулялись так разгулялись. Окраска жуков, попавших на новую территорию, почему-то стала демонстрировать сильную изменчивость, которую до этого не демонстрировала вообще. Понятно, что это опять:
1) Пример внезапного проявления скрытой изменчивости, никогда ранее не наблюдавшейся, но потенциально имевшейся у данного вида.
2) При очевидном отсутствии какой-либо роли естественного отбора в этих проявлениях.
Может быть, это явление аналогично «буму» расцветок многих аквариумных рыбок и некоторых домашних птиц (кур, волнистых попугайчиков и пр.), которые попали в цепкие «лапы» селекционеров в новых для себя (искусственных) условиях. И в этих «лапах селекционеров» отдельные виды животных показали весьма впечатляющую способность к вариации некоторых признаков (например, окраски). А другие признаки (или другие биологические виды) оказались уже не столь отзывчивыми (даже в «лапах селекционеров»), или вообще практически не отзывчивыми. Всё это опять косвенно указывает на то, что одни признаки (и биологические виды) способны к высокой степени вариации, а другие - не способны.
И наконец, обсуждаемые жуки-листоеды продемонстрировали еще и новое (изменившееся) поведение. На участках своего расселения они удивили биологов тем, что стали формировать так называемые «уединенные популяционные волны» (с очень высокой концентрацией жуков в пределах этих «волн»), которые никогда не формировали в своих исходных местообитаниях.
Понятно, что в этом месте можно провести определенные аналогии с уже разбиравшимися выше жабами-агами из Австралии, где биологи тоже зафиксировали изменившееся поведение этих жаб в ходе их расселения по новой территории (см. выше).
ЭВОЛЮЦИЯ ПО ВОЛШЕБСТВУ
Всё-таки, думаю, надо обратить особое внимание еще на одну поразительную деталь из того списка удивительных вещей, которые продемонстрировал нам амброзиевый листоед в новых для него местообитаниях. Если подойти к этой детали внимательно, то получится, что пример с жуками зигограммами, возможно, не подпадает даже под объяснение скрытой изменчивостью. А может быть объяснён только в рамках эволюции по волшебству.
Действительно, пример с «полетевшими жуками» (за 5 поколений) лучше всего укладывается в русло концепции номогенеза - недарвиновской эволюционной теории, предложенной русским биологом Л.С. Бергом (Берг, 1922).
В своей теории Берг постулирует, что одни и те же морфологические изменения происходят дружно, то есть, имеют массовый характер (затрагивают сразу большое число особей). Причем эти массовые особенности проявляются (или даже обусловлены?) именно разностью географических регионов.
Понятно, что в рамках дарвинизма такое просто невозможно – здесь изменения начинаются с отдельных особей (у которых происходит соответствующая случайная мутация). И дальше полезный признак постепенно (за ряд поколений) автоматически распространяется в популяции, в результате преимущественного размножения тех особей, которые обладают этим признаком. Поэтому массового появления нового признака сразу у значительного числа особей в рамках дарвинизма не может быть по определению. А вот в рамках номогенеза, такие дружные изменения являются одним из исходных постулатов теории.
Так же и с «географией». У Берга, кажется, сама география определяет соответствующие изменения. То есть, само нахождение особей того или иного вида на определенной территории, похоже, заставляет этих особей приобретать соответствующие черты, характерные именно для этой территории. В дарвинизме же география играет роль лишь постольку, поскольку она обеспечивает разные условия среды и изоляцию популяций.
Однако Берг особенно не распространяется, какой же конкретно механизм обеспечивает синхронные эволюционные изменения в разных географических регионах. Он ограничивается приведением конкретных примеров синхронных изменений некоторых морфологических признаков у животных и растений, в зависимости от того, в какой конкретно географический регион эти организмы попали. И говорит об эволюции на основе неких «закономерностей». Но что это за закономерности – остаётся почти «за кадром». По сути, получается как бы эволюция по волшебству. Где животные, попадающие в определенную местность, изменяются сообразно этой местности как бы по волшебству.
И вот, обретение жуками способности к полёту происходило совершенно удивительным образом, напоминая, в буквальном смысле, эволюцию по волшебству. Давайте внимательно вчитаемся в соответствующую цитату (Голубева, Опарина, 2011):
При обретении зигограммой способности к полету на первом этапе начинается отвердение задних крыльев, после появляются первые зигограммы, летающие «акробатически» (жук, пытающийся лететь, делает кувырок и падает вниз). И, наконец, вот уже тысячи жуков самостоятельно поднимаются в воздух — а прошло всего-то пять поколений (за лето сменяется как минимум одно поколение, в благоприятных условиях — два-четыре). Дальше «летуны» полностью замещают акклиматизированных «пешеходов».
А вот как описывается это поразительное явление в оригинальной статье (Ковалев, 2001):
Формирование полета у акклиматизированной популяции проходило в несколько этапов. Уже у первых трех поколений УПВ началась склеротизация крыла, поэтому отнесение еще не летающих особей к «канадской популяции» некорректно. До 4-го поколения, когда появились первые «летуны», обычным явлением в популяции был так называемый «кувыркающийся полет» - падение жука при попытке оторваться от субстрата; в 5-м поколении в воздух одновременно поднимались уже тысячи насекомых.
При чтении этих строк охватывает оторопь. Понятно, что об одном «включении», приводящем сразу ко всем изменениям, необходимым для полёта, здесь речь тоже не идет. Мы видим несколько последовательных стадий, которые направлены именно в нужную сторону. Всё выглядит так, как будто жуки на каком-то общем собрании договорились между собой дружно начать летать. И целеустремленно осуществили поставленную задачу. Причем сразу на всех новых для них географических территориях: в Европе, Китае, на Дальнем Востоке. Это им удалось сделать в массовом порядке, но с помощью нескольких последовательных шагов, всего за 5 поколений.
В общем, поразительный факт. Ясно одно - этот пример не влезает ни в какие ворота дарвинизма. Кстати, точно к такому же выводу пришел и сам автор статьи про «полетевших жуков» (Ковалев, 2001):
Таким образом, формирование в УПВ в течение четырех поколений новой формы с четкими таксономическими признаками и вытеснение этой формой уже акклиматизированных «пешеходов», а также появление необычайного для вида фенотипического разнообразия не может быть объяснено исходя из представлений синтетической теории эволюции о плавном (градуальном) превращении популяции исходного вида в некоторую новую популяцию, контролируемую естественным отбором.
Сам Ковалев предпочитает говорить об этом факте, как о механизме видообразования на базе «системных мутаций», связанных с наличием определенного комплекса внутренних (геномных) и внешних (средовых) условий (Ковалев, 2001). То есть, говоря простыми словами, о той самой скрытой изменчивости (скрытых возможностях), уже «запрятанных» в генотипе этого вида, и пробуждаемых внешними условиями.
В любом случае, это изменения не по Дарвину. Это либо проявление мощной скрытой изменчивости, либо вообще эволюция по волшебству.
И наконец, последнее. Столкнувшись с этим примером (жуки, которые дружно решили начать летать), верующий дарвинист, скорее всего, согласится с Вами, что это не пример эволюционных новшеств, а реализация возможности, которая уже имелась у этих жуков. Но (скажет дарвинист) это у них «включилось» их эволюционное прошлое. То есть, эти жуки раньше умели летать, а потом у них эта способность деградировала (по тем или иным причинам). Но эта деградация (как оказалось!) еще не окончательная - в данном случае (и в новых условиях) эволюция еще могла повернуть вспять, что она (эволюция) и продемонстрировала.
Такие рассуждения дарвиниста лично для Вас некритичны. Поэтому здесь поступайте так, как Вам больше нравится - можете избрать агрессивный стиль, а можете, наоборот, спокойный:
1. Вы можете не согласиться с дарвинистом. Сказать, что его предположения - не более чем теоретические спекуляции о гипотетическом эволюционном прошлом жуков, которые невозможно проверить. И потребуйте от дарвиниста предоставить Вам огарок той свечи, которую этот дарвинист (наверное?) держал всё то время, пока эти жуки (на его глазах?) постепенно «отказывались от полета за ненадобностью».
2. Или же Вы можете сначала согласиться с дарвинистом. Сказать, что пусть будет так. Но это не отменяет того установленного факта, что скрытая изменчивость у разных биологических видов - имеется (хотя бы в варианте «призраков прошлого»). Поэтому прежде чем приводить «строго установленные примеры эволюции»... следует трижды подумать, насколько действительно «строги» эти примеры. Не являются ли и эти примеры тоже, на самом деле, просто «призраками прошлого»?
Получается, что мы не можем быть строго уверены ни в одном примере видимой эволюции, какой бы пример изменений мы ни нашли в живой природе. Ведь соответствующие «эволюционные изменения» вполне могут оказаться проявлением той самой скрытой изменчивости, а вовсе не наблюдаемой эволюцией.
И когда дарвинист освоит эту мысль, далее скажите ему, что и предположения о «призраках прошлого» тоже недостаточно строги. Потому что когда речь идёт о прошлом, то это ведь, на самом деле, гипотетическое прошлое. А как известно, гипотетическое прошлое далеко не всегда оказывается реальным прошлым. Поэтому строго оперировать мы можем лишь самим фактом существования скрытой изменчивости. А соответствующую эволюцию - лишь предполагать. Таким образом, у нас вообще не остаётся строго установленных примеров эволюции (никаких).
Кстати, такое постепенное подведение дарвиниста к нужной мысли через другую, «переходную» мысль, напомнило мне еще один установленный пример очень быстрых «эволюционных изменений», тоже весьма интригующий. Это знаменитые «тли Шапошникова» (Шапошников, 1961).
ТЛИ ШАПОШНИКОВА
В этом исследовании Шапошников тоже вывел «новый вид» тлей (очень быстро, практически за один сезон), которые смогли жить на новом растении хозяине, ранее абсолютно непригодном для этих тлей… после того, как Шапошников «потренировал» этих тлей на другом, «переходном» хозяине, более подходящем для исходных тлей.
Самое удивительное в этих исследованиях было то, что всё происходило очень быстро. Шапошников пересаживал примерно по 10 тлей на новое растение, а потом отсаживал обратно, для «восстановления жизненных сил». Таким образом, тли «тренировались» выживать на новом растении, на котором они исходно не питались, и не размножались. И по прошествии очень небольшого числа поколений (не более восьми!) тли вдруг начинали и питаться, и размножаться на новом растении. Причем этот переход всегда носил скачкообразный характер - сначала выживаемость тлей на новом растении имела неустойчивый характер. Но после определённого, конкретного поколения, тли вдруг дружно приживались на новом растении.
Остановлюсь только на самом ярком получившемся результате. Вот графики приживаемости тли Dysaphis anthrisci majkopica на новых (для этого вида тлей) растениях Chaerophyllum bulbosum и Сh. maculatum (Рис. 17):
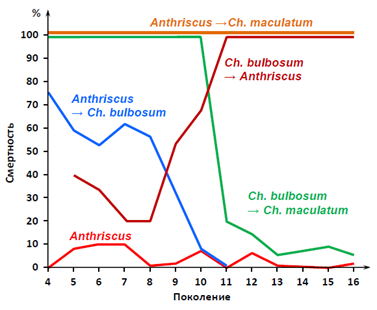
Рис y нок 17 . Графики, показывающие смертность и приживаемость тлей в ряду поколений на новых для них растениях в экспериментах Шапошникова.[36]
Из графика видно, что тли этого подвида:
1) Неплохо прижились на растении Anthriscus (красная линия внизу). Хорошо видно, что смертность тлей на этом растении - минимальна (это контрольная группа, которая никуда не пересаживалась).
2) При прямом пересаживании тлей с растения Anthriscus на растение Ch. maculatum - тли данного подвида не питались, не размножались и гарантированно (100%) гибли (верхняя оранжевая линия).
3) Зато тли оказались способны «пересесть» c растения Anthriscus на Сh. bulbosum (синяя линия на графике). Причем до 8-го поколения наблюдались резкие колебания плодовитости и смертности, но после 8-го поколения - тли прижились на новом растении, и смертность резко упала.
4) Более того, тли, прижившиеся на новом растении (Ch. bulbosum), почему-то потеряли способность питаться на старом растении (Anthriscus). Из графика видно, что после всё того же восьмого (опять восьмого!) поколения - смертность этих «новых» тлей на старом растении резко повышается, достигнув к 11 поколению отметки в 100% (темно-красная линия на графике, резко ползущая вверх).
5) В то же время, тли, прижившиеся на новом растении (Ch. bulbosum) - неожиданно приобрели способность питаться и размножаться на растении Ch. maculatum, которое изначально было вообще непригодным для исходных тлей данного подвида. Хорошо видно, что зеленая линия (смертности тлей) на графике крайне резко «падает» вниз на 10-11 поколениях. Что говорит о весьма синхронном «переключении» тлей. Это выяснилось после того, как тлей с Ch. bulbosum начали регулярно пересаживать на Ch. maculatum. В результате этих «тренировок», уже на 8-9 поколениях тли получили способность питаться на новом для них растении Ch. maculatum (причем дружно), а с 10—11 поколений уже нормально размножались на этом растении.
В итоге получилось, что тли необратимо «пересели» с одного растения на другое... всего за 11 поколений. Необратимо потому, что тли перестали питаться на старом растении. Более того, Шапошников провел скрещивания между полученными линиями тлей, и выяснил, что даже жизнеспособность оплодотворенных яиц между этими линиями упала чуть ли не до нуля, в то время как внутри линий - с жизнеспособностью яиц всё было хорошо (Шапошников, 1961). То есть, возникла, по сути, еще и репродуктивная изоляция.
Более того, при переходе тлей с одного растения на другое произошли еще и морфологические изменения тлей. Причем изменения весьма интересные - тли, перешедшие на новые растения… стали напоминать два других вида тлей (из этого же рода), всегда питавшихся на этих растениях. Например, проявилось сходство в строении хоботка и трубочек (Шапошников, 1965). То есть, получилась еще и определенная конвергенция форм (за 10 поколений).
Другой подвид тлей (Dysaphis anthrisci chaerophyllina) тоже повел себя весьма характерным образом: [37]
В опытах с A. chaerophyllina тли для формирования контрольной группы были пересажены на Ch. maculatum (вторичный хозяин), где прижились и до осени 1958 года дали на этом растении 47 поколений. Из контрольной группы насекомые пересаживались на новое кормовое растение (Ch. bulbosum), где также прижились, хотя до 8-го поколения их жизнедеятельность была неустойчивой из-за резких колебаний смертности.
И опять обратим внимание на назойливую цифру «8». Я затрудняюсь подробно интерпретировать эти результаты. Картина получается сложной, возможных вариантов объяснений много, и в них можно запутаться. Заострю внимание только на наиболее ярких деталях:
1. Мы опять наблюдаем очень быструю «эволюцию». Что довольно странно в рамках постулата о «случайных мутациях».
2. «Эволюция» успешно завершается каждый раз за 8 поколений, несмотря на очень малый биологический материал (около десятка «тренируемых» тлей). Что еще более странно для предположений о «случайных мутациях».
3. Наблюдается синхронность переходов тлей с одного пищевого объекта на другой. Постоянно фигурирует цифра - восемь (или около того). Это уже совсем противоречит предположению о «случайных мутациях».[38]
Возможно, все эти странности и можно как-то уместить в рамках постулируемого механизма «случайные мутации + естественный отбор». Но гораздо удобнее эти факты объясняются в рамках представлений о всё той же скрытой изменчивости (или даже о скрытых «переключателях» в геноме тлей), позволяющих им весьма быстро переключаться с одного субстрата на другой.
Интересно, что обсуждая полученные результаты, сам Шапошников в одних случаях пишет о «пищевых настройках» и «пищевых предпочтениях». Это когда пищевые изменения были обратимы (например, тли D. anthrisci chaerophyllina, перейдя на растение Ch. bulbosum, тем не менее, не потеряли возможность жить на исходном Ch. maculatum). А в других случаях Шапошников пишет о «пищевых адаптациях», в том числе, глубоких. Которые приводят к освоению нового растительного субстрата тлями, но с потерей способности питаться на прежнем субстрате. И далее рассуждает, являются ли «пищевые настройки», «пищевые предпочтения» и «пищевые адаптации» - проявлением неких скрытых потенций, то есть, возможностей, уже имевшихся у этих видов? Или же здесь имеет место изменение самих этих возможностей? Шапошников склоняется к тому, что в случае «пищевых настроек и предпочтений» реализуется первый вариант, а в случае «пищевых адаптаций» (во всяком случае, глубоких), реализуется второй вариант (Шапошников, 1961). То есть, Шапошников задаётся теми же вопросами, которыми сейчас задаёмся и мы. А что это такое продемонстрировали нам тли? Свои возможности по освоению новых видов кормовых растений, которые у них уже имелись в скрытом виде? Или же всё-таки настоящее изменение (эволюцию) своих пищевых адаптаций?
Более того, Шапошников приводит ряд других научных публикаций, где были зарегистрированы примеры таких же быстрых переходов насекомых фитофагов с одного растения на другое, которые тоже происходили за считанное число поколений. Например, установленные примеры с виноградной филлоксерой и т.п. (см. Шапошников, 1961). Получается, что такие быстрые переходы, возможно, достаточно широко распространены в природе. И является ли такой акт перехода насекомого-фитофага (с узкой специализацией) на новое (более-менее подходящее) растение - эволюционным актом? Или же эти насекомые «всегда так умели», то есть, такая способность (к быстрым пищевым переходам) исходна заложена в их геномах в виде «скрытых эволюционных потенций» - этот вопрос до сих пор висит в воздухе и «требует будущих исследований».
ЗУБАСТЫЕ КУРЫ
И в заключение темы скрытой изменчивости я приведу еще один пример. Выше я уже привёл: 1) возможные примеры реализации скрытой изменчивости, 2) явные примеры скрытой изменчивости, 3) самые яркие примеры скрытой изменчивости.
А теперь приведу самые необъяснимые примеры скрытой изменчивости. Авторы работы (Harris et al., 2006) умудрились пронаблюдать... зубастого цыпленка. Они выяснили, что у цыпленка имеется в скрытом состоянии некая генетическая программа развития зубов – челюсти птицы имеют скрытую способность участвовать в зубном морфогенезе (в той или иной степени). Поскольку эта генетическая программа восприимчива к определенным воздействиям, то в ответ на эти воздействия у цыпленка могут начать формироваться зубы.
Этот простой, единичный, но удивительный пример, на самом деле, является весьма тяжелым фактом для современной теории эволюции. И удивительна здесь не столько очередная демонстрация скрытой изменчивости. К ней мы уже привыкли, и уже не особенно удивляемся. Хотя на этот раз в раздел скрытой изменчивости попали уже морфологические признаки весьма серьезного таксономического уровня, явно не видового. Но удивительно здесь другое. В рамках общепринятой эволюционной парадигмы, свои последние зубы далекие предки современных птиц «растеряли» уже очень давно. Как минимум, 70-80 млн. лет назад (Harris et al., 2006). Естественно, возникает вопрос - каким образом эта скрытая способность к формированию зубов смогла сохраниться (в скрытом виде) все эти 70-80 млн. лет? То есть, не проходя «очищение» (проверку на работоспособность) естественным отбором, эта генетическая программа, тем не менее, умудрилась не сломаться под потоком случайных мутаций, которые случайно «выбивали нуклеотиды» из генов этой генетической программы все эти долгие 70 млн. лет?
Вся концепция «молекулярных часов», на которой, в свою очередь, построены все филогенетические деревья (эволюционные схемы родства) основана на том, что поток случайных мутаций «неотвратим, как смерть и налоги» (С). И только естественный отбор (теоретически) может противостоять этому разрушающему процессу, отбраковывая вредные мутации и оставляя здоровых, немутантных особей... А тут, зубы вот уже 70 млн. лет, как перестали появляться у птиц... поток случайных мутаций не ослабевает все эти 70 млн. лет... Но любознательные биологи взяли, и решили исследовать этот вопрос. И откуда ни возьмись, вполне работоспособная программа развития зубов вдруг «взяла под козырёк» и начала формировать цыпленку зубы! Это как вообще понимать?
Поневоле начинаешь задумываться, а были ли вообще эти 70 млн. лет в реальности?
И тут же вспоминаешь еще про 40 млн. лет. Ведь именно столько (предполагаемо) просуществовала у муравьев рода Pheidole их скрытая способность к формированию касты сверхсолдат (см. выше).
Но самый главный вывод из всего, что здесь прозвучало, лежит в несколько иной плоскости.
Если геномы биологических видов имеют некий «пласт» скрытой изменчивости, причем (в некоторых случаях) в скрытом виде могут храниться даже признаки надвидового уровня... То возникает вопрос - а можно ли вообще на сегодняшнем этапе развития биологии найти такой пример наблюдаемых морфологических изменений какого-нибудь признака (того или иного биологического вида), про который можно было бы сказать совершенно точно - да, вот это изменение точно произошло по механизму естественного (или полового) отбора случайных мутаций? Ведь стоит дарвинисту найти такой пример, как его оппонент, ссылаясь на установленные факты скрытой изменчивости, всегда может возразить, что пример дарвиниста - это, может быть, вовсе и не демонстрация эволюции по механизму «случайные мутации + естественный отбор», а просто проявление ранее скрытых возможностей, которые уже были предусмотрены для этого вида.
К сожалению, выхода из этого тупика, действительно, нет. До тех пор, пока мы не научимся не только читать, но еще и понимать, что же записано в геномах организмов (т.е. понимать содержание генетических программ), до тех пор мы не сможем уверенно ответить на озвученный выше вопрос. Получается, что на современном уровне развития биологии, со строгими доказательствами дарвинизма имеются огромные проблемы.
Во-первых, самих по себе строго установленных примеров эволюционных изменений (каких-нибудь морфологических видовых признаков), которые бы вышли за рамки наблюдаемой изменчивости (характерной для этого вида) - очень мало.
Во-вторых, в тех очень редких случаях, когда нужные изменения всё же были установлены - роль естественного отбора, как назло, не изучалась.
В-третьих, там, где давление естественного отбора как раз изучалось, и было установлено - там почему-то (видимо, опять назло) морфологические признаки вообще не выходят за рамки наблюдаемой изменчивости вида.
И наконец, в четвертых - если бы мы всё-таки отыскали в природе такой пример, где и нужное давление естественного отбора было бы установлено, и признак вышел бы за границы наблюдаемой изменчивости... то и в этом случае всегда останется возможность, что данный признак, на самом деле, никуда не «вышел», а просто реализовалась скрытая изменчивость, которая всегда имелась у этого вида.
А как же тогда строго доказать реальность предложенного механизма эволюции «случайные мутации + естественный отбор»? А никак не доказать. На современном этапе развития биологии (до того, как мы научимся понимать, что конкретно записано в геномах биологических видов) - никак не доказать. Имеется в виду, конечно, строгое доказательство.
Поэтому вместо строгих доказательств остаются только косвенные свидетельства (непонятно какой) «наблюдаемой эволюции», от которых при внимательном рассмотрении тоже мало что остаётся (см. выше). Это и есть та самая эмпирическая пустота современной теории эволюции, о которой я говорил выше. Как в сказке Андерсена про голого короля. И сегодня (как и 150 лет назад) в справедливость дарвиновских механизмов эволюции нам остаётся пока только верить (или не верить).
Однако как только Вы всё это озвучите верующему дарвинисту, «эмпирическая пустота» теории эволюции сразу же наполнится «скрежетом зубовным» - возмущенными репликами, недовольным ропотом (или писком) верующего дарвиниста. Дарвинист обязательно скажет Вам, что:
- Ваши критерии «строго доказанного примера» - чересчур строгие, а на самом деле, и так всё ясно (что там всё происходило именно под действием естественного отбора)!
Или что:
- Тезис о скрытой изменчивости - демагогический, приводящий к принципиальной невозможности доказательства справедливости механизма случайных мутаций.
На этот возмущенный «скрежет зубовный» Вам надо реагировать спокойно. Отвечайте следующее (и стойте на этом):
1. «Критерии строго доказанного примера» - отнюдь не строгие, а совершенно нормальные для любого научного исследования (в любой другой области, кроме дарвинизма).
2. Тезис о скрытой изменчивости, действительно, переводит дарвинизм в непроверяемую область (на современном уровне развития биологии). То есть, в область веры и теоретических предпочтений. Однако такая ситуация совсем не редка в науке. В науке сплошь и рядом складываются ситуации, когда ту или иную гипотезу невозможно проверить в существующих условиях (по самым разным причинам). И никто от этого почему-то не впадает в истерику. Такие вещи воспринимаются спокойно.
3. И наконец, даже без всякой скрытой изменчивости - теория естественного отбора в эмпирическом плане все равно представляет из себя практически пустоту.
ОПЯТЬ «ЭВОЛЮЦИЯ» У ГУППИ
В заключение представлю еще один (типичный) пример эмпирической пустоты современной теории эволюции. Это классический (наиболее распространенный) случай, когда за эволюцию выдается обычное колебание признака в рамках внутривидовой изменчивости.
Понятно, что подобные недоразумения встречаются не только в публикациях про ящериц, но и в статьях, посвященных «эволюции» других биологических таксонов. Именно такое очередное недоразумение было опубликовано в уже достаточно старом исследовании про гуппи, «эволюционировавших на водопаде» (Reznick et al., 1997). Рассматриваю это исследование только потому, что его результаты тоже каким-то образом сумели просочиться в список тех самых 12.5 примеров «строго установленной эволюции», которыми пользуются дарвинисты в пропаганде своего «единственно верного учения».[39]
Если кратко, в этой работе авторы рассказывают о том, что в популяциях гуппи, подверженных сильному хищническому давлению, время достижения самцами и самками гуппи половозрелости – меньше, чем в популяциях, где хищническое давление является низким. То есть, в тех популяциях, где эти рыбки подвергаются повышенному риску хищничества, они «созревают» раньше, чем в популяциях с низким давлением хищников. Точно так же и размер особей, достигших половозрелости, в популяциях гуппи с высоким хищническим давлением – меньше, чем в популяциях гуппи с низким хищническим давлением. Скорее всего, эти параметры связаны. То есть, средний размер тела вступающих в размножение гуппи меньше именно потому, что рыбки вынуждены созревать раньше.
Понятно, что там, где рыбки каждую минуту рискуют быть съеденными до размножения, становится выгодным созреть как можно раньше, чтобы успеть разножиться до того, как тебя съедят.
И вот авторы исследования нашли такие места в природных местообитаниях гуппи, которые различались степенью хищнического давления. Они нашли участки реки, разделенные водопадами, где в нижней части (под водопадом) гуппи обитали совместно с хищной цихлидой Crenicichla alta. А также с другими видами хищных цихлид и еще c хищными представителями семейства харациновых. Некоторые из этих видов (в том числе, и озвученная Crenicichla alta) предпочитают охотиться на любых взрослых гуппи обоих полов без разбора.
А вот в верхней части (над водопадом) живет только цихлида Rivulus hartii, которая является всеядной, и лишь иногда охотится на гуппи. Поэтому данный участок реки можно считать участком с низким хищническим давлением (для гуппи).
Авторы исследования нашли такие места (над водопадом), где гуппи вообще отсутствовали. И пересадили туда группу гуппи из-под водопада. То есть, исследователи пересадили гуппи из участков с высоким хищническим давлением в участки с исходно низким хищническим давлением. И потом в течение 11 лет наблюдали за изменениями, происходившими в популяциях гуппи «над водопадами».
О результатах этих наблюдений авторы доложили в той самой статье, про которую здесь идет речь (Reznick et al., 1997). В этой статье авторы утверждают, что они обнаружили эволюцию гуппи под действием естественного отбора. На основании того, что гуппи «над водопадом» на реке Aripo стали теперь (спустя 11 лет) достигать половой зрелости примерно на 9-20% позже того исходного значения, которое наблюдалось «под водопадом». Примерно в рамках таких же значений (13-17%) увеличилась и масса самцов и самок, впервые вступающих в размножение «над водопадом», по сравнению с массой самцов и самок, впервые вступающих в размножение «под водопадом».
На втором опытном участке, на реке El Cedro, спустя 7.5 лет после переселения, время достижения половозрелости «над водопадом» тоже увеличилось (примерно на 4-11%), и рыбки, впервые вступающие в размножение, тоже стали тяжелее (на 4-7%).[40]
Отсюда авторы почему-то сделали вывод, что гуппи эволюционировали под действием соответствующего естественного отбора. Правда, не очень понятно, на что конкретно здесь «давил» естественный отбор?
Ведь можно теоретически рассудить, что быстрое достижение половозрелости – лучше в любом случае, независимо от присутствия или отсутствия хищников. Потому что фактор времени работает против этих рыбок в любом случае. Помимо хищников, рыбки могут заболеть инфекционными заболеваниями, или даже просто травмироваться. Поэтому тянуть с размножением (в рамках дарвиновских представлений о природе) вряд ли стоит.
Однако на подобные теоретические рассуждения столь же теоретически можно возразить, что повышенный размер особей, вступающих в первое размножение, может быть, благотворно сказывается на дальнейшей продолжительности их жизни, и как следствие, на всё том же размножении (плодовитости) этой рыбки.
Например, одно дело, когда растение вырастает до высоты одного метра, и сразу же начинает размножаться. И совсем другое дело, когда растение «терпит», откладывая собственное размножение до «лучших времен», и вырастает высотой в 10 метров. И только после этого начинает размножаться. Понятно, что в этом случае растение сможет произвести на свет гораздо большее число потомков (или лучше обеспечить своё потомство ресурсами, или и то и другое вместе), чем растение, которое «поторопилось» с размножением, достигнув размера всего один метр. С другой стороны, пока растение растёт до высоты 10 метров, с этим растением может случиться много всяких неприятностей. В том числе, и роковых неприятностей, которые могут привести к гибели этого растения. В таком случае, оно вообще не оставит никакого потомства. Понятно, что это тоже плохо.
Поэтому описанные выше «торопливые» и «неторопливые» растения можно рассматривать, как разные стратегии размножения. Каждая из этих (практически противоположных) стратегий имеет свои достоинства и недостатки. И в результате, обе эти стратегии, в принципе, могут быть «взяты на вооружение» разными организмами. В современной биологии так и считается. Описанные стратегии размножения известны, как r,K-стратегии. Где к r-стратегам относятся организмы, «торопливые» до размножения, а к K-стратегам – организмы, «неторопливые» до размножения.
Как я только что сказал, каждая из этих стратегий в общем, вполне достойна, чтобы реализоваться в живой природе (теми или иными организмами). Однако конкретные условия окружающей среды (в каком-нибудь конкретном местообитании) могут сделать одну стратегию более выгодной (в этих условиях), чем другую.
Например, в нестабильных местообитаниях, где жизнь конкретной особи может в любой момент прерваться от каких-нибудь «факторов неодолимой силы», становится выгодной именно r-стратегия. То есть, становится выгодно вырасти и созреть как можно быстрее, чтобы успеть оставить потомство до того момента, как погибнешь.
И наоборот, в стабильных и благоприятных условиях определенные преимущества получает K-стратегия. Например, растение K-стратег в таких местообитаниях может не размножаться, а тратить все ресурсы на дальнейший рост. В результате оно перерастёт всех своих конкурентов (которые «поторопились» с началом размножения), и просто задавит их в конкурентной борьбе (например, за счет перекрывания доступа к свету). После этого такое растение может спокойно начать размножаться. Ведь оно уже захватило весь необходимый ему объем ресурсов, и теперь может спокойно и долго жить в этом богатом и стабильном местообитании, из года в год производя всё новое потомство. Причем каждый раз понемногу, чтобы не «надорваться», и не быть вытесненным конкурентами из этого (уже завоеванного) «райского уголка». Но общее количество потомства, произведенное таким растением-доминантом за все годы его жизни, может оказаться просто огромным.
Можно предположить, что что-то подобное имеет место и в популяциях гуппи. А именно, в популяциях с высоким риском погибнуть в зубах хищника становится выгодно как можно быстрее вырасти и дать (хотя бы первое) потомство. А в популяциях с низким риском хищничества, возможно, получают преимущество более «долгоиграющие» особи гуппи?
Может быть. Но всё это теория. Пускай и красивая. А вот в «суровой реальности», то есть, конкретно в этой статье - реальные преимущества, получаемые от «запоздалого» созревания – вообще не измерялись.
Между тем, скорость соответствующих «эволюционных» изменений, которые продемонстрировали гуппи «над водопадом», оказалась (опять!) чрезвычайно высокой для предположенного авторами механизма эволюции (естественный отбор). Действительно, гуппи «над водопадом» на реке Aripo продемонстрировали соответствующие эволюционные изменения всего за 18 поколений. А гуппи «над водопадом» на реке El Cedro успели продемонстрировать «эволюцию» уже примерно за 13 поколений. Причем самцы гуппи над водопадом реки El Cedro опередили в этом отношении самок, и продемонстрировали «эволюцию» вообще за 7 поколений!
Стоит ли говорить, что такие молниеносные изменения вряд ли возможны в рамках естественного отбора. Для столь скоростного «естественного отбора» необходимо, чтобы преимущество в общей приспособленности, которое даёт новый признак (впервые появившийся у какой-то отдельной особи), было просто чудовищным (не менее 200-300%).[41] Между тем, в обсуждаемой статье это (воображаемое) преимущество… вообще не измерялось.
Хотя и без всяких измерений ясно, что запаздывание со временем вступления в размножение примерно на 9-20% по сравнению с исходными гуппи вряд ли способно увеличить число потомков сразу на 200-300%. Таким образом, в лице «гуппи над водопадом», мы имеем перед собой не просто недоказанный пример «установленной эволюции под действием естественного отбора», а пример, где «эволюция под действием естественного отбора» является самым невероятным сценарием из всех возможных.
Уже понятно, какой сценарий здесь является наиболее правдоподобным. Это примерно такой же сценарий, как и в описанном выше примере с массовым укорочением ног ящериц всего за 3 (!) поколения.
А именно, столь скоростной сдвиг во времени достижения половозрелости и массы тела половозрелых особей мог быть достигнут только благодаря тому, что пересаженная часть гуппи на момент начала эксперимента уже имела достаточно много конкретных особей, которые созревали позже, чем другие особи. Например, допустим, исследователи пересадили «наверх» группу рыбок, где 70% особей созревали на 20% раньше, а остальные 30% особей созревали на 20% позже. Понятно, что «под водопадом» рано созревающие гуппи имели определенные преимущества – они успевали размножиться несколько чаще, чем поздно созревающие гуппи, до того момента, как их съедали хищники. И поэтому, допустим, в популяции и поддерживалось именно соотношение 70:30 в пользу «скороспелых» гуппи, а не меньше.
Но после того как исследователи пересадили гуппи из-под водопада туда, где давление хищников было низким, фактор времени стал играть уже меньшую роль в жизни этих маленьких рыбок. Поэтому те «долгоиграющие» гуппи, которых раньше в популяции было меньше (потому что они оставляли потомство несколько реже), в новых условиях стали оставлять потомство чаще, чем раньше. И в результате, доля таких особей в экспериментальной группе постепенно немного повысилась (в течение ряда поколений). Что и привело (в среднем) к установленному сдвигу сроков «созревания» в популяции в целом (на 10-20%). Действительно, такой сдвиг вполне мог произойти из-за небольшого изменения в пропорции «ранне-спелых» и «поздне-спелых» гуппи, уже имеющихся в популяции. Причем весьма быстро.
Таким образом, здесь мы имеем сдвиг нормы признака за счет того, что соответствующее давление естественного отбора было снято. То есть, это не пример сработавшего естественного отбора, а его противоположность.
Впрочем, можно предположить, что достижение половозрелости в ускоренные сроки имеет для гуппи определенную цену. Может быть даже, не все гуппи способны эту цену платить. Тогда в отсутствие хищников рыбки, созревающие немного медленней, получают некоторое преимущество. То есть, в рамках подобного предположения, установленный сдвиг сроков созревания можно рассматривать именно как результат естественного отбора – отбора, направленного в сторону меньшей «цены» взросления.
Но даже если этот сдвиг возник именно под действием естественного отбора», а не наоборот (не чисто автоматически в результате снятия соответствующего давления хищников), то разве этот пример является примером эволюции? Нет, не является. Потому что здесь просто произошел небольшой сдвиг признака в рамках тех значений, которые уже имелись исходно. Причем, во-первых, имелись сразу у значительного числа особей, а во-вторых, даже не в каких-то других популяциях этого биологического вида, а прямо в этой, конкретной популяции.
Все мы хорошо знаем, что у людей одни дети растут и взрослеют быстрее, чем другие. И можем предположить, что хотя бы какая-то часть этого признака имеет наследственный характер. И если вдруг произойдет что-нибудь такое, в результате чего поздно взрослеющие дети немного увеличат свою долю (например, на 20%) где-нибудь во Владивостоке, то разве это будет означать, что люди куда-то сэволюционировали? Или это будет означать лишь то, что норма этого признака просто немного передвинулась внутри (уже имевшихся) значений, характерных для этого биологического вида?
Можем ли мы на основании подобного факта сделать вывод, что люди эволюционируют, и через сто тысяч поколений станут взрослеть, допустим, только к сорока годам? Я думаю, нет, не можем. Подобное предположение будет абсолютно спекулятивным. То есть, это что угодно, но только не «строго установленный пример эволюции».
Наконец, не исключено, что в данном случае механизмы изменений были вообще другими, не такими, как это представляется с «высоты птичьего полёта». Может быть, за произошедшие изменения в данном случае ответственны какие-то специальные приспособительные механизмы, которые у гуппи «имеются про запас» на этот случай?
Например, в ботанике известен так называемый «эффект группы» - где молодые растения, оказавшиеся в достаточно плотной группе, растут существенно лучше и быстрее, чем растения, произрастающие поодиночке.
Или гораздо более известный пример – акселерация развития у человека в течение последних 150 лет. По поводу возможных механизмов этой акселерации выдвинуто много разных гипотез. И гипотезы «естественного отбора» (быстрее созревающих людей) занимают довольно скромное место в этом ряду.
Возможно, что-то похожее имеется и у гуппи? Допустим, если гуппи живут в воде, где присутствуют химические следы опасных хищников (или даже следы «трагедий» от других гуппи, попавших на зуб хищникам), то в этих условиях развитие гуппи, может быть, несколько ускоряется?
В связи с этим очень полезно вспомнить, что в недавнем исследовании, о котором я уже рассказывал выше (Ruell et al., 2013) было установлено, что самцы гуппи в ходе индивидуального развития изменяют степень яркости своей окраски в зависимости от присутствия или отсутствия хищников. Но если гуппи могут поступать так со своей окраской, то почему бы им не поступать таким же образом еще и со скоростью своего созревания?
Тем более что в ряде других исследований (посвященных изучению других биологических объектов) подобные эффекты были реально установлены именно в отношении скорости «созревания». Например, в недавней работе (Pose et al., 2013) было установлено, что растение резуховидка Таля (Arabido psis thaliana) вполне успешно регулирует время своего зацветания в зависимости от температуры окружающей среды. Причем механизм этой регуляции – генетический (альтернативный сплайсинг).
А раз механизм генетический, то здесь не исключена возможность эпигенетических эффектов, когда соответствущие изменения в генах могут специально закрепляться и становиться наследственными (в течение хотя бы определенного ряда поколений). В этом случае мы получим долговременную модификацию. И это было бы так похоже на «эволюцию», что дарвинисты, без сомнения, не упустили бы шанс объявить это эволюцией… Если бы такие изменения не включались (внутри организма) определенными факторами внешней среды, и если бы подобная реакция не была уже заложена в геноме биологических видов заранее.
И такие эффекты тоже были установлены. Например, в недавнем нашумевшем исследовании (Dias & Ressler, 2013) было показано, что благодаря эпигенетическим механизмам, дети и даже внуки мышей продолжают бояться того самого запаха, которым исследователи напугали еще их отцов и даже дедов. Это открытие, без сомнения, чрезвычайно интересно само по себе. А уж какие «поправки» оно может внести в теорию биологии - просто не пересказать. По сути, это оживший ламаркизм в действии.
Таким образом, мы видим, как дарвинизм мешает нормальному научному поиску – вместо углубленного изучения всех возможных причин того или иного биологического явления, дарвинизм предлагает универсальную «пилюлю» на все случаи жизни – что бы (и где бы) в природе ни случилось – это непременно «эволюция под действием естественного отбора» и никак иначе.
И последнее. Выше я так красочно рассуждал об r,K-стратегиях. Об их достоинствах и недостатках. Что и говорить, наши теоретические схемы весьма часто получаются красивыми и убедительными. Однако это еще не значит, что эти теоретические схемы работают в реальной природе. Наоборот, живая природа имеет обыкновение ломать красивые теоретические схемы. За счет потрясающего разнообразия примеров «на все случаи жизни». Например, выше я уже рассказывал о пальме Tahina spectabilis, которая упорно растёт много лет, чтобы завоевать себе место под солнцем в таких местообитаниях (богатых и благоприятных), где завоевать себе это место весьма тяжело. И вот когда эта пальма, наконец, становится огромным растением, с потенциальными возможностями произвести огромное количество собственного потомства в течение целого ряда лет… В этот момент пальма приступает к своему первому и единственному размножению, после которого благополучно… умирает. Понятно, что этот конкретный биологический пример не лезет ни в какие ворота наших (таких очевидных) теоретических рассуждений о разных стратегиях размножения организмов в рамках биологической выгоды (или не выгоды). Совершенно ясно, что этой удивительной пальме просто начихать на биологическую выгоду в частности, и на дарвиновскую теорию в целом. Потому что дарвиновские представления о живой природе целиком и полностью основываются именно на рассуждениях о биологической выгоде (или не выгоде). А эта пальма не следует нашим теоретическим рассуждениям о биологической выгоде, а поступает совершенно противоположным образом.
И таких примеров, когда даже самые очевидные теоретические представления разбиваются реальными биологическими примерами – в живой природе вагон и маленькая тележка. Начиная с того, что рассуждения об эволюции в ходе естественного отбора - тоже кажутся весьма убедительными (и даже очевидными). Однако найти в природе такую эволюцию мы почему-то не можем.
И только верующие дарвинисты твердо «знают», что фундаментальные законы, по которым существует живая природа, уже давно раскрыты. И перечислены в книжке Докинза «Эгоистичный ген».
Дата: 2019-02-02, просмотров: 526.