Теперь перейдем к рассмотрению других фактов «наблюдаемой эволюции», которые могут быть известны дарвинисту, вступившему с Вами в дискуссию.
Но дальше будет уже проще. Потому что строго установленные примеры дарвиновской эволюции в природе, собственно, уже закончились (на «эволюции березовых пядениц»). Остальные известные примеры «наблюдаемых эволюций» уже далеки от какой-либо строгой доказанности. Они или установлены недостаточно надежно, или это примеры таких изменений, которые совершенно точно не вышли за рамки внутривидовой изменчивости. Или же это изменения, которые произошли не по дарвиновским механизмам, а по каким-то другим. Причем часто всё это бывает смешано в одном флаконе, то есть, в одном и том же примере «наблюдаемой эволюции». Однако верующий дарвинист всё равно может представить Вам подобную «гремучую смесь» в качестве «строго установленного примера дарвиновской эволюции».
Для того чтобы Вам было легче разбираться с подобными «гремучими смесями», ниже я приведу показательный пример одной такой (реально состоявшейся) дискуссии на эту тему. Частично в виде «прямого репортажа», с цитированием точных реплик из состоявшегося диалога. Взаимными оппонентами здесь выступили два кандидата биологических наук (одним из которых был Ваш покорный слуга). Естественно, некоторые реплики я буду сопровождать пояснительными комментариями.
Дискуссия получилась не по проблеме «наблюдаемой эволюции» вообще, а конкретно про «эволюцию» моллюсков (ракушек и улиток). Но я всё равно рекомендую внимательно проследить за ходом приведенной беседы, поскольку многое из прозвучавшего крайне характерно и для всех остальных споров об эволюции.
Эта «эпическая сага» началась с того, что в завязавшейся беседе с очередным верующим дарвинистом я написал свою традиционную просьбу:
- Не могли бы Вы привести несколько строго доказанных примеров зафиксированной эволюции какого-нибудь видового морфологического признака в природе именно под действием естественного или полового отборов, опубликованных в научных изданиях? Заранее спасибо.
Верующий дарвинист не замедлил с ответом:
- Да полно (и дал соответствующую ссылку).[1]
Я прошел по ссылке и увидел опубликованную статью (Seeley, 1986), с которой был еще не знаком. Насколько я понял сразу же из резюме статьи, речь шла о зарегистрированных эволюционных изменениях формы раковин, произошедших примерно за 100 лет под предполагаемым действием естественного отбора. Я написал об этом дарвинисту:
- Пардон, но Вы предъявили мне ссылку... на предположение. Я же Вас просил - строго доказанные факты. Не могли бы Вы привести еще примеры (строго доказанные)? Заранее спасибо.
Дарвинист принял вид оскорбленной добродетели:
- Я Вам привел уже примеры эволюции у бактерий, у моллюсков, у мышей.[2] Давайте Вы напишите заранее, сколько статей Вам нужно привести, а то так и будете требовать больше и больше. И это не предположение, а хорошо обоснованное утверждение, опубликованное в PNAS.
По опыту я знаю, что если начать разбираться с подобными статьями, то как правило, обнаруживается вопиющее несоответствие заявлений дарвиниста (о наблюдавшейся эволюции) с тем, что написано либо в самой статье, либо в других статьях, посвященных этой же теме. Поэтому я не стал продолжать спор с дарвинистом до того, как ознакомлюсь с материалом. Я лишь попросил дарвиниста привести еще какие-нибудь опубликованные материалы из этой же области, которые бы подтверждали результаты первой статьи:
- Ох, только сейчас увидел, что публикация 1986 года. В связи с этим, не могли бы Вы привести независимые исследования, подтвердившие результаты этого исследования? А то это такая скандальная область, где невоспроизводимость результатов - массовое явление.
Дарвинист не замедлил привести мне еще три опубликованные научные статьи:
- Хорошо. Вот независимые аналогичные исследования с различными моллюсками.[3] То есть эволюция раковин моллюсков наблюдалась разными людьми на разных модельных объектах.
Поскольку было уже поздно, я взял тайм-аут и пошел спать. А с утра сел за статьи, и то, что я в них прочитал, меня не удивило. Но зато ввергло в печальные мысли по поводу деградации общего уровня профессионализма многих российских биологов.
Итак, самая первая приведенная статья была опубликована в 1986 году и в переводе называется примерно так (Seeley, 1986):
«Интенсивный естественный отбор вызвал быстрый морфологический переход у морских улиток»
Автор этой работы сравнил формы раковин моллюска Littorina obtusata, собранные в разных районах восточного побережья США в 1871, 1898 и 1915 годах, с формой раковин этого же моллюска, собранных примерно в этих же местах, но уже в 1982-1984 годах. Исследователь обнаружил, что за прошедшие 100 лет форма большинства раковин (но не всех) заметно изменилась.
Вот соответствующая часть рисунка из обсуждаемой статьи (Рис. 1):
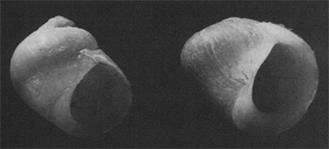
Рисунок 1. Верхняя часть рисунка из оригинальной статьи ( Seeley , 1986). Слева показана раковина Littorina obtusata, собранная в 1871 году с острова Appledore (Мэн, США). Справа показана раковина Littorina obtusata, собранная на этом же острове, но уже в 1982 году.
Как видим, морфология раковин, действительно, заметно различается.
Автор обсуждаемой статьи предположил, что эти изменения связаны с появлением в 1900 году на восточном побережье США пришлого вида краба Carcinus maenas, который (в том числе) питается улитками данного вида. Автор предположил, что измененная форма раковины L. obtusata обладает лучшей защитой, чем исходная, а именно, является более прочной, и тяжелее взламывается крабом, чем раковина исходной формы.
Наконец, автор предположил, что подобные изменения раковин произошли в результате естественного отбора этих раковин (под хищническим давлением крабов).
Для проверки этих предположений, автор провел соответствующие полевые и лабораторные эксперименты. Результаты исследований ясно показали, что раковины измененной формы, действительно, тяжелее преодолеваются хищными крабами, чем раковины исходной формы.
В полевых исследованиях автор работы прикреплял попарно раковины с исходной формой (которые еще встречаются на некоторых участках этого побережья) вместе с раковинами измененной формы, в местообитаниях трех разных типов, где наблюдалось: 1) большое изобилие крабов; 2) крабы обычны, но менее изобильны; 3) крабы встречаются изредка.
Результаты оказались такими:
1) На участках, где крабы были обильны, выжило 57% улиток с «толстой» (измененной) раковиной (от исходного числа всех выставленных «толстых» раковин) и только 14% улиток с «тонкой» раковиной (исходной формы).
2) На участках, менее заселенных крабами, выжило 89% «толстых» раковин и только 33% «тонких».
3) И наконец, на участках, где крабы были редки, выживание улиток с «толстой» и «тонкой» формами раковины оказалось одинаковым (100%).
В лабораторных экспериментах все 100% «тонких» раковин были успешно атакованы крабами, против всего 12% успешно разрушенных «толстых» раковин.
Из этих результатов становится ясно, что на тех участках побережья, где крабы достаточно обильны, улитки с измененной («толстой») формой раковины, действительно, получают существенные преимущества в выживании.
В связи с такими результатами, автор работы предположил, что изменение формы раковины улитки Littorina obtusata за прошедшие 100 лет произошло в результате именно естественного отбора под действием хищнического давления краба-вселенца. В разделе «Discussion» автор специально подчёркивает, что эволюция в ходе естественного отбора в качестве объяснения произошедшего изменения формы раковин - является лишь предположением (поскольку время уже ушло и точно сказать уже невозможно). Однако поскольку, во-первых, изменение формы раковины имеет явно защитный характер, а во-вторых, хорошо совпадает со временем вселения в данный регион краба Carcinus maenas (начало 20 века), то автор заключает, что именно эволюция в ходе естественного отбора является наиболее вероятной причиной зафиксированных изменений.
Итак, в итоге мы получаем, что наш дарвинист, вроде бы, ни в чём нам не соврал. Приведенная им статья действительно является, пусть и не совсем уж строгим, но весьма убедительным примером именно эволюционных морфологических изменений, причем именно под действием естественного отбора - в качестве наиболее вероятной (или даже практически очевидной) причины.
Правда, автор обсуждаемой статьи получил еще и дополнительный результат - что форма раковин изменилась подозрительно быстро. Ведь 100 прошедших лет - это для улиток данного вида - всего примерно 50 сменившихся поколений.[4] Более того, промежуточная форма раковины была установлена уже у улиток, собранных в 1915 году (Seeley, 1986). Получается, что переход от исходной формы к промежуточной вообще произошел менее чем за 9 поколений. В рамках представлений об эволюции в ходе естественного отбора, такая скорость изменений получается чрезвычайно высокой. Для того чтобы подобная скорость эволюции оказалась возможной именно по механизму естественного отбора, общая выживаемость раковин с «толстой» морфологией должна превышать выживаемость раковин с «тонкой» морфологией в три-четыре раза. Это, конечно, очень много. Тем не менее, чисто теоретически, разница в общей приспособленности 300-400%, в принципе, возможна. И поскольку автор работы верит в свой естественный отбор, то он делает вывод - значит, естественный отбор способен работать даже так стремительно.
Ну что же, теперь нам осталось почитать еще и другие, более поздние статьи на эту тему, и посмотреть, насколько подтвердились выводы, сделанные исследователем в 1986 году.
Но сначала давайте познакомимся поближе с самим героем нашего «эволюционного подвига» - с улиткой Littorina obtusata.
И вот тут мы узнаём, что форма раковины этой улитки чрезвычайно сильно варьирует от популяции к популяции. То есть, данный вид улиток демонстрирует очень большую внутривидовую изменчивость именно по форме раковины (Рис. 2):
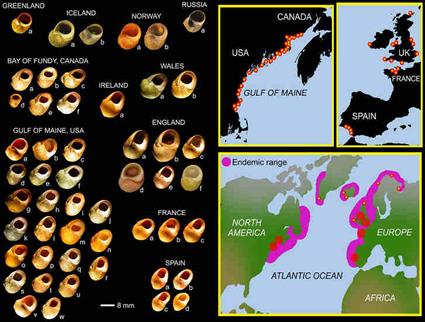
Рисунок 2. География и внутривидовая изменчивость улитки Littorina obtusata.[5]
Не правда ли, на фоне подобного морфологического разнообразия размера, цвета и формы раковин одного и того же биологического вида Littorina obtusata, требование нашего дарвиниста признать именно эволюцию данного вида - начинает выглядеть весьма поспешным, или даже можно сказать, маниакальным. Становится понятно, что отмеченные в рассмотренной выше статье изменения формы раковины (случившиеся в одной из частей ареала обитания этого вида) - вряд ли вышли за границы внутривидовой изменчивости, уже известной для этого вида и прежде.
Получается, что мы попросили (у нашего дарвиниста) пример, иллюстрирующий эволюционные морфологические изменения биологического вида. А в ответ получили очередной пример «эволюции»… из «стройной» улитки Littorina obtusata в более «толстую» улитку Littorina obtusata. Причем заранее известно, что такие «толстяки» внутри рамок этого биологического вида встречались и раньше, в других местах обитания этого вида (см. например, форму раковин «английских» улиток на рисунке 2).
То есть (сократив все лишние слова) мы получили пример «эволюции» Littorina obtusata в Littorina obtusata.
Таким образом, верующие дарвинисты (практически всегда) демонстрируют крайнюю небрежность по отношению к аргументам своих оппонентов. Или же это не небрежность, а сознательная подтасовка - когда Вы ясно пишете о примерах морфологической эволюции именно видового уровня... а взамен получаете эволюцию какого-нибудь самца гуппи из одной популяции... в самца гуппи, известного по другой популяции.
Собственно, на этом можно было бы и закончить. Но оказывается, самое интересное ожидает нас впереди, в ходе дальнейшего ознакомления с этой замечательной улиткой.
Дальше мы узнаём, что изменение формы раковины - это, на самом деле, вообще не эволюционные изменения, а модификационные (т.е. приобретаемые в течение жизни). Оказывается, улитка данного вида (L. obtusata) просто соответствующим образом изменяет форму раковины (в ходе собственного роста), если в воде присутствуют химические «следы» крабов. Или не изменяет (форму раковины), если химические следы крабов в воде отсутствуют. Другими словами, улитка точно так же меняет форму своей раковины в ответ на химические следы присутствия крабов, как человек наращивает себе мышечную массу в ответ на физическую нагрузку.
Это классический пример пластичности (модификационной изменчивости). Когда организм в течение жизни изменяется под действием тех или иных факторов окружающей среды. Данный эффект (пластичность особи) в рамках конкретно дарвинизма - не имеет никакого отношения к какой бы то ни было эволюции.
То есть дело было, видимо, так. Вначале (в далеких восьмидесятых годах) исследователь зафиксировал интересное явление - изменение формы раковин за 100 лет. Этот исследователь предположил, что в таком изменении раковин «повинна» именно эволюция под действием естественного отбора.
Затем другие заинтересовавшиеся биологи провели специальные исследования… и выявили интересный морфо-физиологический эффект. Оказывается, форма раковин улиток пластично изменяется в зависимости от присутствия или отсутствия химических следов крабов в воде. А вовсе не в результате какой-то там «эволюции».
Например, автор работы (Edgel, 2008) проводил исследование именно с L ittorina obtusata. К моменту проведения этого исследования было уже известно, что «потолстение» улиток - результат пластичности. И в данном исследовании выяснялось, различается ли величина этого эффекта у представителей британских популяций этих улиток в сравнении с их американскими «коллегами». Исследователь вырастил британских и американских улиток в присутствии и в отсутствии химических следов хищников. И вот что получил:
...После четырех месяцев, «следы опасности» вызвали адаптивные изменения в индивидуальной форме раковин – от тонких к толстым стенкам раковин.
Вот еще одно исследование, посвященное изучению (некоторых аспектов) эффекта пластичности у этого же вида Littorina obtusata (Trussell & Nicklin, 2002). Авторы исследования пишут в резюме:
...Химические «следы опасности» сразу от обеих реплик - и от крабов и от поврежденных остатков (других) улиток вызвали самое большое увеличение толщины раковины, тогда как подвергание только «следам опасности» от крабов вызвало промежуточные уровни утолщения…
Вывод, который следует из всей этой информации, возможен только один - ни о каком строгом доказательстве эволюции американских улиток (да еще и в результате естественного отбора) здесь не может быть и речи. Скорее всего, мы наблюдаем просто эффекты пластичности. Присутствуют в окрестностях крабы - форма раковин улиток изменяется. Отсутствуют крабы - форма раковин не изменяется.
Отметим, что в рамках представлений именно об эффекте пластичности (как о причине обсуждаемых изменений) легко объясняется и очень быстрая (для естественного отбора) скорость реакции на прибытие хищника - изменение формы раковин всего за 9 поколений. То есть, не успел еще хищник появиться (в новых для него местообитаниях) и как следует размножиться … а соответствующая реакция на него улиток уже тут как тут.
Наконец, следует обратить внимание, что ареал распространения конкретно этого вида улитки (см. рис. выше) – это оба побережья Северной части Атлантического океана (с двух сторон). И популяции этих улиток у побережья Европы, как говорится, испокон веков жили совместно с обсуждаемым видом краба (для которого это побережье - исконный ареал). Следует ли после этого удивляться, что американские популяции L ittorina obtusata после прибытия к ним их исконного хищника… стали реагировать на него примерно так же, как и улитки из европейских популяций L. obtusata? Действительно, не стоит. Ведь это единый биологический вид.
Итак, в свете обнаружившихся фактов - категоричное требование дарвиниста признать эволюцию на основании подобного «факта наблюдаемой эволюции» - не может вызвать ничего, кроме недоумения. Действительно, вникал ли вообще этот дарвинист в тот пример, который он решил привести в качестве «строгого доказательства эволюции»?
На этом можно было бы и закончить, если бы этот же дарвинист не накидал нам еще три (другие) научные статьи, утверждая, что и они тоже являются независимыми доказательствами эволюции в этой области (т.е. в наблюдаемой эволюции улиток).
Ну что же, давайте наберемся терпения, и посмотрим еще и на эти «доказательства».
Первая статья (Appleton & Palmer, 1988) была опубликована уже давно и имеет весьма длинное название, которое можно перевести примерно так:
«Переносимые водой стимулы, выпущенные хищными крабами и поврежденной добычей, вызывают развитие более защищенных от хищников раковин у морских брюхоногих моллюсков»
Резюме статьи:
Морфологически изменяющиеся особи улиток скалистых приливных зон (Thais (или Nucella) lamellosa) развивают большие раковинные зубцы, когда находятся в присутствии продуктов хищного краба Cancer productus, чем в их отсутствие, независимо от того, питались улитки или нет. Кроме того, среди питавшихся улиток большие раковинные зубцы продуцировались в присутствии крабов, питавшихся остатками улиток, чем в присутствии крабов, питавшихся замороженной рыбой. Поcкольку все улитки содержались в контейнерах, через которые вода протекала от физически отделенного аквариума с содержащимися крабами, эти результаты показывают, что водорастворимые химические «реплики», выпущенные этими хищными крабами и поврежденными остатками улиток, вызывают у T. lamel losa улучшение защитной эффективности их раковин. Наконец, когда предоставлялся доступ к еде, улитки, подвергавшиеся «стимулированию» химическими «репликами» крабов, съели меньше моллюсков и выросли меньше, чем улитки в контроле.
Итак, уже из содержания резюме очевидно, что в этой работе опять исследовалась именно пластичность раковин улиток (уже другого вида, Thais lamellos a), по отношению к химическим сигналам присутствия в воде хищных крабов (и остатков их добычи). Ни о каком «доказательстве наблюдаемой эволюции» в этой статье нет даже намёка.
Поэтому остаётся совершенно неизвестным, зачем дарвинист привёл эту статью. Такое ощущение, что он вообще не вникал в предоставляемые им ссылки, а накидывал просто всё, что было связано с первой статьей (нашлось в интернете в базах данных научных публикаций). За такие вещи (за ссылки на литературу, не имеющие никакого отношения к поднятой теме) мы обычно снижаем оценки студентам по их дипломным и курсовым работам. Остаётся только надеяться, что этот конкретный дарвинист при подготовке своей диссертации был более аккуратным, чем в спорах с «этими невежественными креационистами».
Вторая статья (Pascoal et al., 2012), которую представил дарвинист в качестве еще одного подтверждения «наблюдаемой эволюции улиток», тоже имеет длинное название:
«Модификационная и наследственная изменчивость толщины раковины брюхоногого моллюска Nucella lapillus, связанная с риском хищничества крабов и действием волн, и половым созреванием»
Уже из названия статьи ясно, о чем в статье речь - о модификационной и наследственной изменчивости толщины раковин, в зависимости от разных факторов среды. Ни о какой «наблюдаемой эволюции улиток» (да еще и под действием естественного отбора) здесь речи вообще нет.
Резюме статьи настолько подробно, что необходимость читать саму статью уже, наверное, отпадает:
Улитка приливной зоны Nucella lapillus обычно имеет более толстую раковину в местообитаниях, защищенных от действия волн, где крабы обильны и представляют высокую угрозу хищничества, чем в подверженных воздействию волн местах, где крабы редки. Мы изучили две популяции, показывавшие противоположные тенденции. Мы взаимно пересадили улиток между полевыми участками и измерили длину раковины, ширину и толщину «губы» этих раковин 12 месяцев спустя. Улитки, пересаженные на защищенный участок, выросли больше, чем «родные» улитки с этого защищенного участка, которые, в свою очередь, выросли больше, чем улитки, пересаженные на участки, подверженные действию волн. Относительная толщина раковинной губы была больше у «родных» улиток участка, подверженного волнам, чем на защищенном участке. Улитки, пересаженные с защищенного участка на участок, подверженный действию волн, развивали сравнительно более толстые раковины, чем в контроле, и относительно более тонкие раковины при пересадке с участка, подверженного действию волн, на защищенный участок. Потомство двух популяций выращивалось в течение 12 месяцев в садке со «сточными водами» от крабов, питающихся сломанными остатками улиток в эксперименте, и в чистой морской воде в контроле. В эксперименте со «сточными водами крабов» улитки уменьшили активность питания, снизили одновременно и соматический рост, и плодовитость. Молодые раковины, получающие «сточные воды» (химические следы) крабов, наращивали длину раковины медленнее, пока не развивали более толстые раковинные губы, чем в контроле, степень ответа была сходной между линиями. F2-поколение линии с участка, подверженного действию волн, показало тенденции, сходные с первым поколением; F2-поколение с защищенного участка оказалось слишком малочисленным для статистического анализа. В состоянии половой зрелости, толщина раковинной губы была больше у улиток, получавших «сточные воды» крабов, чем в контроле, демонстрируя пластичность, но была также больше в линии из защищенных местообитаний, чем в линии, подверженной воздействию волн, указывая на наследственную изменчивость. Результаты подтверждают гипотезу, что «защитное» утолщение раковины – это просто пассивное последствие голодания и что наследственный и пластичный контроль защитной морфологии раковины – действует синергетически. Утолщение раковин молодых улиток было сходным между линиями, вопреки гипотезе, предсказывающей разную силу пластичности в популяциях из местообитаний с низким и высоким риском.
Как видим, в статье изучаются исключительно особенности модификационной и наследственной изменчивости между двумя линиями улиток - из популяции с высоким риском хищничества (где крабы обильны) и популяции с низким риском хищничества (где крабы редки). Интересно, что авторы получили некую «смесь» результатов (см. текст, выделенный жирным шрифтом). В то время как по некоторым признакам (например, по толщине раковинной губы) были выявлены наследственные различия между популяциями, по другим «защитным» признакам, например, по утолщению молодых раковин, теоретически ожидаемую разницу между популяциями с высоким и низким риском хищничества обнаружить не удалось (обе линии оказались сходными в этом отношении). Но это уже не относится к теме нашего обсуждения.
Где здесь наш дарвинист разглядел хотя бы упоминание об эволюции или о естественном отборе - снова остаётся загадкой. Что практически подтверждает наши подозрения – «просветлённый дарвинист» просто накидывал «примеры», не то что не вникая в их содержание, но даже не читая резюме этих работ!
И наконец, посмотрим на последнюю работу (Fisher et al., 2009). Хотя в целом уже и так всё ясно, но зато эта работа, сама по себе, весьма интересна. Статья называется:
« Улитка приливной зоны показывает драматичное изменение размера в течение прошлого столетия»
Поскольку уже само название статьи звучит весьма многообещающе (как бы намекая на морфологическую эволюцию), давайте познакомимся с текстом самой статьи.
Вот что пишут авторы статьи во введении:
…В течение прошлого столетия, береговые сообщества залива Мэн (северо-запад Атлантического океана) испытали существенные сокращения в размере и обилии эксплуатируемой рыбы и беспозвоночных (1,2,9), изменения температуры (10), и внедрение и распространение инвазивных видов, включая краба Carcinus maenas (1,11). В течение этого периода, исторические изменения в форме раковины и её толщины у Nucella lapillus (11) и других улиток (12) в пределах залива Мэн объяснялись исключительно, как ответ на появление краба C. maenas.
То есть, оказывается, на северо-западном побережье Атлантического океана за прошедшее столетие поменялось еще много чего (помимо появления хищного краба).
Далее авторы статьи сравнивают размеры раковин Nucella lapillus, собранные в 2007 году ровно на тех же участках (судя по сохранившимся описаниям), где эти раковины собирались в 1915-1922 годах. И делают заключение, что раковины явно увеличились в размерах:
…Наше самое поразительное открытие, о котором не сообщали ранее для N. lapillus, состоит в том, что длина раковины увеличилась на всех 19 участках, в среднем, на 22.6% между 1915-1922 и 2007 периодами сбора.
Итак, авторы зарегистрировали весьма существенное увеличение среднего размера этого вида морских улиток, которое произошло в этом регионе за прошедшее столетие. Как же они объясняют это изменение? Эволюцией? Естественным отбором?
Давайте посмотрим раздел « Результаты и обсуждение». Приведу обширную цитату:
Предыдущие экспериментальные исследования (14,15) продемонстрировали, что присутствие водорастворимых химических «следов» крабов вызывает утолщение раковинных губ и уменьшение темпов роста; но хотя мы и нашли увеличение толщины губы между 1915-22 и 2007 годами, это увеличение происходило из-за полного увеличения размера, а не изменения формы.
В исследованиях, посвященных изучению индуцированных ответов (изменения формы раковины в ответ на присутствие крабов) использовались улитки намного меньшего размера [средняя длина, 14.7-21.6 мм (14); средняя длина примерно 6.5 мм (15)] чем были найдены и в 1915-1922 годах (23.6 мм) и в 2007 годах (28.9). Мы предполагаем, что когда присутствуют «химические следы» крабов, улитки сначала испытывают утолщение раковины, а затем, будучи уже лучше защищены от хищников, сокрушающих раковины, растут в длину до величины, при которой крабы не могут успешно атаковать их. Этот сценарий объяснил бы результаты, полученные в обоих экспериментах (изменения формы раковин в ответ на присутствие крабов) и в нашем исследовании.
Другие возможные факторы существуют, и различение среди множественных потенциальных факторов, которые, возможно, вызвали увеличение в размере между 1915-1922 и 2007 годами, и в изменении изменчивости между разными участками – является трудным. Направленный отбор C. maenas, возможно, мог этому поспособствовать; наибольшее увеличение размера N. lapillus произошло на защищенных территориях, которые наиболее подходят для крабов (20,27). Но наши результаты не могут оказать определенную поддержку для этой гипотезы, потому что другие сопутствующие изменения произошли в пределах этой экосистемы тоже . Современные N. lapillus могут быть больше, потому что они просто растут быстрее, возможно, из-за различия в температуре воды между этими периодами; среднегодовые температуры водной поверхности на прибрежном контрольном участке долгосрочного мониторинга в 120 км юго-западнее МDI были на 1.9 градусов выше в течение 1994-2007, чем в течение 1915-1922 (10).
Кроме того, содержание питательных веществ в области увеличилось за прошлое столетие, и сейчас эвтрофированные скалистые берега увеличили покрытие мидий и усоногих ракообразных по сравнению с неэвтрофированными участками (5); таким образом, долгосрочное увеличение береговой продуктивности, возможно, увеличило темпы роста улиток из-за увеличения доступности добычи. Наконец, хотя C. maenas в настоящее время считается важным хищником (11,15), N. lapillus может теперь фактически испытывать более низкое хищническое давление или высокий темп роста, чем в 1915-1922 годах, из-за потери других хищников. Колтон не упоминает крабов, как источник смертности для N. lapillus, но идентифицирует рыб и чаек, как хищников (18,19). Принимая во внимание, что ранние выводы о хищничестве рыб были основаны на косвенных свидетельствах (22), большая и обильная хищная рыба, которая в прошлом присутствовала близко к берегу (28) теперь заметно отсутствует, делая исключительно трудным сравнение современного хищнического давления инвазивных видов с наблюдавшимся ранее хищническим давлением в условиях изобилия родных хищников в пределах этого региона (1,2,9).
Итак, давайте обратим внимание на поразительное различие между примитивным уровнем анализа, сделанным нашим незадачливым дарвинистом, и уровнем анализа проблемы, выполненным нормальными исследователями.
Верующий дарвинист, выхватив из названия статьи только одну фразу про « драматичное изменение размера»… немедленно требует от своего оппонента признать факт « наблюдаемой эволюции», да еще и по механизму естественного отбора! А нормальные ученые подходят к обнаруженному факту увеличения размеров раковин N. lapillus в исследованном районе весьма вдумчиво, рассматривая его со всех сторон (с точки зрения всех возможных причин).
Мы видим, что на первое место исследователи выдвигают предположение о всё том же явлении пластичности (модификационной изменчивости) формы раковины в ответ на присутствие хищника (а никакой не эволюции). Но не исключают в качестве возможных причин увеличения размера и другие факторы. Причем этих факторов столько, что предположение о направленном отборе улиток крабом С. maenas - буквально тонет в других возможных причинах, которые выглядят гораздо правдоподобней, и опять-таки, отражают не эволюционные изменения, а просто модификационную изменчивость. Начиная с того, что раковины могли просто начать быстрее расти из-за увеличившегося обилия их кормовой базы (которое, в свою очередь, увеличилось вследствие эвтрофикации данной области) и кончая возможностью суммарного снижения (общего) хищнического давления (несмотря на появление нового хищного краба) за счет исчезновения старых исконных хищников данной улитки.
Последнее, что нам осталось, это оценить степень возможной внутривидовой изменчивости улитки N. lapillus и посмотреть, вышли ли зафиксированные изменения средней величины раковин за пределы внутривидовой изменчивости, известной для этого биологического вида раньше. И здесь тоже оказывается всё как всегда. А именно:
1) Сами авторы статьи пишут по этому поводу, что данный вид известен обширной морфологической изменчивостью в разных районах по обе стороны Северной Атлантики (Fisher et al., 2009:5211).
2) На рисунке 3 можно оценить изменчивость улиток данного вида из разных географических областей и в зависимости от разных факторов среды (солености, обилия пищи, расположения на защищенных участках или на участках, подвергающихся воздействию волн):
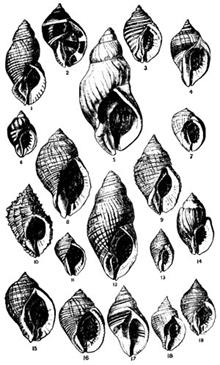
Рисунок 3. Географическая и экологическая изменчивость раковин Nucella lapillus. Рисунок взят из работы (Crothers, 1985:310).
Очевидно, что внутривидовая изменчивость улитки N. lapillus весьма впечатляет.
3) Размеры раковин улиток, увеличившиеся, в среднем, на 22,6% за прошедшее столетие на участках, исследованных авторами работы (Fisher et al., 2009), тем не менее, не вышли за пределы наблюдаемой внутривидовой изменчивости улиток этого вида. В литературе указывается, что размер этих улиток обычно колеблется между 20 и 35 мм длины раковины (Crothers, 1985:291). Но нередко достигает значительно больших величин, доходя до 40 мм, или даже (в отдельных случаях) существенно больше (в одном из источников указано 60 мм).
Если мы посмотрим на ту длину, которую зарегистрировали авторы работы (Fisher et al., 2009), то увидим, что эта длина не вышла даже за границы обычных значений. О преодолении же границ внутривидовой изменчивости вообще не может быть речи (Рис. 4):
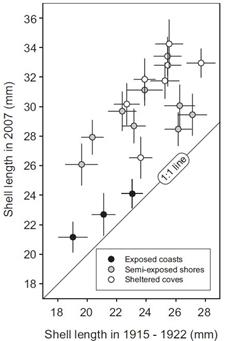
Рисунок 4. По горизонтали – средняя длина раковин Nucella lapillus, собранных в 1915-1922 годах в местообитаниях: защищенных от действия волн; незащищенных от действия волн; промежуточных. По вертикали – длина раковин этого же вида, собранных в тех же самых местах в 2007 году. Рисунок взят из обсуждаемой статьи ( Fisher et al ., 2009).
То есть, всё как всегда - в качестве « строго доказанного примера эволюции видового признака под действием именно естественного отбора»… верующий дарвинист опять приводит пример «эволюции» биологического вида Nucella lapillus... в биологический вид Nucella lapillus. И еще оказывается, что даже такая «эволюция», скорее всего, совсем не эволюция. А просто пример модификационной изменчивости. То есть, прямое изменение формы и размера раковин (в ходе их роста) в ответ на тот или иной фактор окружающей среды. Из возможного десятка разных экологических факторов, которые изменились в этом регионе за последние сто лет.
Наконец, стоит посмотреть еще и на окраску раковин этого вида (Рис. 5):

Рисунок 5. Внутривидовая изменчивость раковин Nucella lapillus по окраске.[6]
Наблюдаемое разнообразие окраски интересно в связи с тем, что в качестве основных хищников этой улитки указываются разные птицы (например, кулики-сороки, чайки, обыкновенные гаги и еще целый ряд видов). А как известно, птицы имеют превосходное зрение, да еще и цветное. В связи с этим непонятно, о чем думает естественный отбор, которому так молятся наши верующие дарвинисты. Почему этот естественный отбор до сих пор не убрал из популяций улиток варианты с выделяющейся окраской?
Причем если кто-то сейчас предположил, что вариации окраски у данной улитки встречаются в соответствии с тем основным фоном, на котором они обитают в разных районах… то он ошибся. Разные варианты окраски у этого вида встречаются очень часто, иногда буквально на одном «пятачке». Примерно вот так (Рис. 6):

Рисунок 6. Разнообразие окраски раковин Nucella lapillus в пределах одного и того же местообитания.[7]
Значит, естественный отбор под хищническим давлением птиц отбирает бабочкам окраску, соответствующую фону, всего за 50 лет (березовым пяденицам). Но этот же естественный отбор, под тем же хищническим давлением птиц, не отбирает окраску, соответствующую природному фону, улиткам. Что и говорить, естественный отбор – весьма неисповедимый товарищ.
Впрочем, может быть, это так постарался половой отбор? Может быть, самки этих улиток придирчиво отбирают своим самцам такую выделяющуюся окраску? Действительно, если бы у этих улиток было хоть сколько-нибудь приличное зрение, то дарвинисты, без всякого сомнения, воспользовались бы таким объяснением.
И наконец, хорошо после всего этого дать верующему дарвинисту как следует помедитировать еще и на слизня (Рис. 7):

Рисунок 7. Разные представители обширной группы слизней – брюхоногих моллюсков, которые либо совсем не имеют раковины, либо имеют очень маленькую «раковинку».[8]
Пусть дарвинист попредставляет себе (используя своё богатое эволюционное воображение), почему в то время как морские улитки столь серьезно озабочены хищническим давлением, что изменяют себе форму и толщину раковины, стоит только хищным крабам объявиться в окрестностях... в это же самое время слизень ничем не обеспокоен, спокойно ползает себе по лесу, не имея почти никакой защиты (кроме собственной слизи), несмотря на то что им охотно питаются: хищные жуки, ежи, кроты, землеройки, некоторые грызуны, лисицы, барсуки, грачи, галки, куропатки, дрозды, скворцы, дикие утки, чайки, лягушки, жабы, саламандры, ящерицы, змеи и другие животные.
Видимо, наш вездесущий естественный отбор просто куда-то запропастился в тот момент, когда создавался слизень? Наверное, естественный отбор в это время занимался исключительно морскими ракушками?
Дата: 2019-02-02, просмотров: 550.