Раз уж мы так подробно разобрали «эволюцию американских улиток», то давайте тогда разберем еще и «эволюцию русских ракушек». Под «русскими ракушками» имеется в виду история с «эволюцией» двустворчатых моллюсков в Аральском море, которую описали в своей работе Андреевы (Андреева, Андреев, 2003). Этот пример «установленной эволюции» довольно популярен именно среди российских почитателей Дарвина. Большинству же зарубежных любителей эволюции этот пример, как правило, не знаком. Потому что все свои работы (посвященные разным аспектам биологии аральских моллюсков и других элементов экосистемы Аральского моря), Андреевы публиковали только в российских журналах. Соответственно, вычитали и разрекламировали этот пример «наблюдаемой эволюции» (вбросив его в широкие народные массы) тоже российские проповедники дарвинизма.
За многие годы исследований, Андреевы проделали очень большую работу, устанавливая те многочисленные изменения, которые происходили с экосистемами быстро высыхавшего Арала. По сути, это был протокол «умирания» целого моря.
К сожалению, благодаря стараниям верующих дарвинистов, широкую известность получила лишь самая слабая часть этих исследований – та глава, в которой авторы рассуждают именно об «эволюции» двустворчатых моллюсков Арала.[15]
Если пересказать эту главу совсем кратко, то авторы пишут, что в условиях быстро разрушающихся исконных экосистем Аральского моря, они обнаружили очень быструю эволюцию отдельных видов ракушек. Такую эволюцию продемонстрировали ракушки из родов Cerastoderma и Syndosmya. Причем продемонстрировали они её всего за 10-15 лет, что примерно равно такому же числу сменившихся поколений. То есть, и здесь тоже (снова) речь идет об «эволюционных» изменениях, происходивших с такой скоростью, которая кажется слишком быстрой для эволюции по дарвиновским механизмам.
Авторы работы описывают многочисленные изменения, происходившие в высыхающем море:
1) Постепенно возрастающую соленость.
2) Другие физико-химические изменения, вплоть до выпадения некоторых солей на дно в виде «корки».
3) Массовое вымирание прежде доминирующих видов.
4) Увеличение численности прежде редких видов, в том числе, завезенных в Арал из других водоемов.
5) Радикальную перестройку пищевых цепей Арала.
6) Массовую гибель макрофитов,[16] вплоть до того, что всё дно моря оказывалось устланным мертвыми остатками этих водных растений.
7) Расцветом фитопланктона.
Много внимания авторы уделяют постепенно возрастающей солености воды Аральского моря, что приводит к вымиранию сначала множества пресноводных видов, а затем и солоноватоводных, с заменой их чисто морскими и континентальными галофильными видами.
И вот в таких условиях конкретно для ракушки Cerastoderma (по утверждению авторов) наступаёт «звёздный час». Во-первых, эта ракушка демонстрирует взрыв внутривидовой изменчивости. То есть, начинает наблюдаться множество форм этого моллюска с изменениями по целому ряду морфологических признаков. Во-вторых, из моря исчезают основные «сдерживатели» популяций ракушки Cerastoderma – моллюски других родов, Dreissena и Hypanis. H ypanis являлся конкурентом Cerastoderma - так же как и Cerastoderma, он «сидел» практически целиком погруженным в грунт, и отфильтровывал взвешенные в воде питательные вещества. То есть, использовал ту же пищу, которой питалась и Cerastoderma. Таким образом, само существование H ypanis сдерживало численность Cerastoderma.
А ракушка Dreissena поступала с Cerastoderma еще жестче – стоило какой-нибудь Cerastoderma высунуть свою раковину из грунта, как на неё оседали личинки Dreissena и закреплялись на ней. Потому что дрейссенам для закрепления нужна какая-нибудь твердая основа. В ходе дальнейшего роста, дрейссены под тяжестью собственного веса постепенно «топили» Cerastoderma в грунте, попутно затрудняя им питание еще и за счет оплетания их своими прикрепляющими нитями (биссусом).[17]
Таким образом, в исконных экосистемах Арала распространение Cerastoderma сдерживалось доминированием дрейссен и Hypanis.
Однако в связи с повышением солености воды Аральского моря, дрейссены и Hypanis вымирают за относительно короткое время. А Cerastoderma не вымирают, потому что возросшая соленость воды пока еще их устраивает (находится в тех пределах, которые допустимы для этого вида). Вот тут-то Cerastoderma и показывает, на что она способна (в плане «эволюции»). Авторы работы отмечают, что в девяностых годах двадцатого века большинство особей Cerastoderma уже «вылезло» из грунта и находилось практически на его поверхности. Что было совсем не характерно для тех популяций Cerastoderma, которые существовали в Аральском море еще 20 лет назад. Авторы объясняют это тем, что в связи с вымиранием дрейссен, освобождается экологическая ниша, где можно питаться мелким планктоном, находясь прямо на поверхности грунта. При этом можно сколько угодно высовывать свою раковину над грунтом – никто уже не «садился» на эту раковину, и не оплетал её своими нитями.
Авторы работы приводят соответствующий рисунок, демонстрирующий это яркое изменение в пищевом поведении Cerastoderma (Рис. 8):
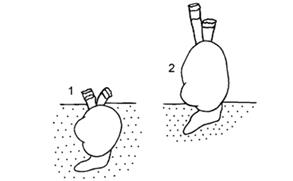
Рисунок 8 . Схема положения Cerastoderma в грунте Аральского моря. 1 – типичное положение Cerastoderma isthmicum , 2 – положение современных Cerastoderma вне грунта. Из работы (Андреева, Андреев, 2003).
Помимо этого изменения в экологии, Cerastoderma 90-ых годов показывают еще и целый комплекс морфологических изменений, которые соответствуют их новому «имиджу» моллюсков, питающихся на поверхности грунта. А именно, у них смещается положение макушек раковины, меняется форма самой раковины (например, удлиняется её задний край), изменяется число и выраженность зубов и ребер раковины, изменяется строение замка (Андреева, Андреев, 2003). Авторы заключают, что большинство этих изменений биологически целесообразны. Они помогают моллюску удерживать устойчивое положение в новых для него условиях - на поверхности грунта. Кроме того, у большинства экземпляров наблюдается расширение мантийной линии в области сифонов.
Авторы приходят к выводу, что поскольку размах только что перечисленных морфологических изменений (т.е. установленных различий между Cerastoderma девяностых и семидесятых годов) выходит за пределы различий, характерных для разных видов двустворок (в других регионах), то здесь уже можно вести речь о факте видообразования. То есть, о появлении нового вида двустворчатого моллюска. Андреевы предполагают, что это видообразование произошло в результате стремительной эволюции под действием естественного отбора.
Такое же предположение авторы делают и в отношении установленных ими новых форм у других ракушек, из рода Syndosmya. Они заключают, что и эти морфологические варианты тоже возникли в результате именно эволюции моллюсков под соответствующим давлением естественного отбора (вследствие приспособления к грунтам разных типов).
Ну что же. Благодаря предыдущей главе, мы с Вами стали теперь уже опытными специалистами по «эволюции моллюсков», и можем понять, что в этом исследовании не так и почему говорить об «эволюции в ходе естественного отбора» здесь явно преждевременно. Давайте перечислим эти недостатки (в порядке убывающей важности):
1. Перед нами исследования, выполненные только одним коллективом авторов. Каких-либо независимых исследований (сделанных другими авторами), мы не имеем. По одной только этой причине говорить о строго установленном примере эволюции здесь уже вряд ли имеет смысл.
2. Самый главный недостаток конкретно этой работы – авторы не выяснили даже, являются ли обнаруженные ими изменения вообще наследственными? Или же установленные изменения ракушек имеют исключительно прижизненный характер? Может быть, все описанные изменения – это результат обыкновенной пластичности (модификационной изменчивости)? Действительно, весьма вероятно, что изменившееся поведение и морфология ракушек – простое следствие изменения условий среды. Может быть, конкретно этот вид ракушек всегда переходит к питанию на поверхности грунта, если в придонных слоях воды повышается количество пищи (планктона). Допустим, моллюск делает это, откликаясь именно на повышенное содержание планктона в соответствующих слоях воды. И при выходе на поверхность грунта у моллюска соответствующим образом изменяется еще и форма раковины. В этом случае мы будем наблюдать именно такую картину, которую наблюдали Андреевы - моллюски быстро и массово «вылезут» на поверхность грунта, демонстрируя при этом ряд соответствующих морфологических изменений… если в нужных слоях воды появится необходимое количество пищи. А оно там появилось, в связи с полным вымиранием других видов двустворок, которые раньше снижали содержание планктона в воде, а теперь снижать перестали (в связи с собственным вымиранием). Более того, авторы сами пишут об установленном ими повышении обилия планктона в воде Аральского моря.
То есть, возможно (и даже весьма вероятно), озвученные морфологические и поведенческие изменения, установленные авторами для этих видов ракушек, на самом деле, были, во-первых, характерны для этих видов. А во-вторых, являются не результатом эволюции, а просто результатом пластичности в ходе их индивидуального развития. В предыдущей главе мы видели, как улитки разных видов в ходе собственного роста образовывали более толстые раковины, если в воде присутствовал запах хищных крабов. Возможно, таким же образом и описанные ракушки Аральского моря способны выбираться из грунта, при этом соответствующим образом изменяя форму раковины, если в соответствующих слоях воды повышается обилие планктона? Или, допустим, если исчезают химические следы конкурента (Hypanis). Авторы работы не показали, что это не так.
Между тем, именно такое предположение наиболее вероятно. Чтобы его проверить, авторам необходимо было вырастить эти, «эволюционно измененные виды» ракушек в нормальных условиях (т.е. имитирующих условия Аральского моря в его исходном состоянии). И посмотреть, в кого конкретно превратятся изучаемые ими ракушки в таких условиях. Если эти ракушки в ходе роста всё равно продемонстрируют комплекс подмеченных авторами изменений, то значит, эти изменения, действительно, имеют наследственный характер. Если же в нормальных условиях «эволюционно измененные» ракушки тоже вырастут в совершенно нормальные формы (известные прежде), тогда это результат пластичности. И ни о какой «эволюции» в таком случае не может быть и речи.
Авторы такую проверку не сделали. Тем не менее, они почему-то пишут именно про «эволюцию моллюсков», игнорируя более простые варианты объяснений.
3. Авторы работы, скорее всего, справедливо связывают наблюдавшийся ими «взрыв» морфологической изменчивости моллюсков со стрессовыми условиями среды, в которые эти ракушки попали. Например, вот что они пишут по поводу развития личинок Syndosmya segmentum в воде Аральского моря: [18]
…Отставание в выклеве личинок в аральской воде по сравнению с контрольной азовской составляло 4 суток, а в оседании – 5 суток. Также было отмечено, что повышенное содержание иона Ca2+ приводит к появлению у личинок синдосмии уродливых недоразвитых раковинок (Карпевич, 1964).
Кроме того, авторы связывают взрыв изменчивости еще и с исчезновением стабилизирующего отбора в условиях вымирания основных конкурентов этих ракушек. Действительно, если раньше была серьезная борьба за выживание, навязанная конкурирующими видами, то выживать могли только такие ракушки, морфология которых была наиболее оптимальной для выживания. При исчезновении же основных конкурентов, выживать стало легче, и поэтому те морфологические варианты, которые раньше просто не доживали до того счастливого момента, когда их соберут в коллекцию биологи, теперь стали доживать, и удивлять биологов своим «взрывом разнообразия».
Однако непонятно, почему авторы работы пишут о повышении общего мутационного фона у этих ракушек:
Эволюционные изменения акклиматизированного в Аральском море двустворчатого моллюска синдосмия кратко можно представить следующим образом. Перемещение группы особей в водоем с водой, нарушающей ход эмбрионального развития, провоцирует взрыв мутагенеза.
Да, действительно, стрессовые условия вполне могут приводить к сбоям в индивидуальном развитии, приводящим к разнообразным (прижизненным) уродствам. Но почему авторы решили, что эти стрессовые условия (или вызванные ими уродства) каким-то образом должны повышать частоту мутаций у ракушек, и как следствие, приводить к повышенному разнообразию генотипов? Это предположение авторов не подверждено ничем.
4. Наконец, вполне может быть и так, что установленные авторами различия между ракушками семидесятых и девяностых годов 20 века, действительно, являются наследственными. Но почему авторы решили, что за подобные изменения был ответственен именно естественный отбор? Авторы лишь предполагают, что какие-то из установленных ими форм ракушек - более приспособлены к одним условиям (например, к грунтам определенного типа), а какие-то формы более приспособлены к другим условиям. Однако авторы не попытались выяснить, а так ли это на самом деле. И даже если так, то на какую конкретно величину повышается приспособленность разных форм моллюсков в разных условиях? Ведь для того, чтобы всего за 10-15 поколений какая-то форма моллюсков (встречавшаяся раньше в единичных экземлярах) могла бы массово завоевать морское дно на соответствующих грунтах… для этого разница в общей выживаемости между разными формами ракушек должна составлять 200-300%. Так ли это на самом деле?
Например, авторы пишут, что «вылезшие» на поверхность грунта ракушки Cerastoderma находились на этом грунте в довольно неустойчивом положении – когда подплывал аквалангист, ракушки схлопывали створки… и в результате сразу же падали на бок. Возникает вопрос, насколько безобидны такие «побочные эффекты» для моллюска, и перекрывают ли выгоды от питания именно на поверхности грунта столь неустойчивое положение раковины? А если перекрывают, то насколько конкретно? Особенно если учесть, что раньше эти же моллюски вполне успешно выживали, полностью погрузившись в грунт. И продолжают успешно выживать именно таким (погруженным) способом на участках, подверженных действию волн (Андреева, Андреев, 2003).
В общем, сила давления естественного отбора в данной работе не измерялась вообще никак. Просто было сделано предположение о том, что такая сила имелась, и что эта сила была достаточно мощной, чтобы отобрать соответствующие формы ракушек всего за 10-15 поколений. Понятно, что в качестве предположения, такие рассуждения, наверное, могут быть озвучены. Но вот о «строго установленном примере эволюции под действием естественного отбора» здесь говорить, конечно, не приходится.
5. Авторы работы пишут:
…Эта чрезвычайная скорость, с которой «притираются» возникшие фенотипы к новым условиям обитания, создает иллюзию направленного формирования фенотипов средой обитания. Однако, массовая ненаправленная изменчивость среди двустворок Арала (см. глава 6–7) в критические периоды свидетельствует о случайном характере процесса. И только массовое вовлечение в этот процесс двустворок, имеющих колоссальную энергию размножения, способствует успешной работе отбора. Именно массовость материала позволяет сохраняться и размножаться фенотипам (и генотипам), соответствующим новой среде обитания и постепенно (примерно за 10–15 лет), как в случае с эпифаунистическими Cerastoderma, выйти в новую адаптивную зону.
После прочтения этой цитаты напрашивается вопрос – почему авторы решили, что наблюдаемое ими «направленное формирование фенотипов средой обитания» - это именно «иллюзия», а не настоящая причина установленных изменений? То есть, если что-то «выглядит как утка, ходит как утка и крякает как утка»… то почему это всё-таки не утка, а «естественный отбор»?
Авторы объясняют (свой вывод о естественном отборе) наблюдаемой ими «массовой ненаправленной изменчивостью среди двустворок Арала в критические периоды». Но разве «массовая ненаправленная изменчивость» не могла быть просто массовыми сбоями индивидуального развития вследствие экологического стресса (т.е. просто уродствами), а вот наблюдаемое «выползание» ракушек на поверхность грунта (с соответствующими морфологическими изменениями раковин) - следствием именно «направленного формирования фенотипов средой обитания»? То есть, может быть, здесь нужно рассматривать мух отдельно, а котлеты – отдельно?
В свою очередь, «направленное формирование фенотипов средой обитания» - это, может быть, просто пример банальной экологической пластичности этих моллюсков?
Например, всем известно, что сосна в благоприятных условиях – это одно дерево, а вот на болоте - это уже совсем другое дерево. Ну а на обдуваемых ветром скалах – это уже третье дерево. Во всяком случае, морфологические различия между этими соснами будут никак не меньше, чем установленные различия между морфологией аральских ракушек. Так неужели из этого факта следует сделать вывод, что сосна таким образом… «эволюционирует под действием естественного отбора»? Нет, в данном случае сосна демонстрирует лишь тот размах экологической пластичности, к которой способен данный биологический вид. А эволюция здесь вообще ни при чем.
Далее авторы аргументируют свои рассуждения (о том, что это была именно эволюция естественным отбором) массовостью биологического материала, который предоставляют для эволюции обсуждаемые виды ракушек, благодаря их «колоссальной энергии размножения».
Но если так рассуждать, тогда куда более массовый «материал для эволюции» смогут легко предоставить, например, практически все виды планктонных водорослей. Или почти столь же колоссальные по численности и скорости сменяющихся поколений разнообразнейшие представители зоопланктона. Или даже многие виды рыб.[19] То есть, следуя подобной логике, специалисты, изучающие фитопланктон рек, озер и океанов, должны докладывать о новых фактах «свершившейся эволюции» чуть ли не каждый год – в каком-нибудь очередном полувысохшем озере, где плавают миллиарды особей каких-нибудь диатомовых водорослей, подвергаясь соответствующему экологическому стрессу. Или каких-нибудь коловраток. Или нематод. Или каких-нибудь кальмаров (в океане).
Но что-то мы не видим мощного потока научных публикаций, рапортующих нам о многочисленных фактах свершившейся (морфологической) эволюции. Наоборот, мы знаем, что даже самые массовые и быстро размножающиеся виды почему-то устойчиво сохраняют свой набор морфологических признаков именно в тех пределах, которые характерны для этих видов. Несмотря на многочисленные локальные изменения среды в самых разных регионах Земли. И вместо «мощного потока научных публикаций», нам приходится искать днем с огнём наши несчастные «двенадцать с половиной примеров эволюции» во всей живой природе. И один из таких примеров мы находим в книге Андреевых… в виде теоретических рассуждений о том, что обнаруженные ими изменения ракушек – это именно «эволюция под действием естественного отбора», потому что эти ракушки обладают «колоссальной энергией размножения». Что и говорить, просто «убойный» аргумент.
6. И наконец, как я уже говорил выше, установленные авторами изменения у аральских ракушек всё-таки могут быть наследственными. Такая возможность не исключена. И даже достаточно вероятна. Но эти изменения вполне могли быть обеспечены какими-нибудь другими механизмами, отнюдь не дарвиновскими. Например, под действием ныне модных эпигенетических механизмов.
Допустим, авторы этой работы просто натолкнулись на такие два вида ракушек, у которых имеется определенный набор генов, в обычных условиях «спящих». Но если эти гены проснутся, то они «переводят» данный вид ракушек в другой эколого-морфологический «режим» - из погруженного в грунт моллюска - в ракушку, питающуюся на поверхности грунта. Допустим, соответствующие гены «просыпаются» у моллюска под действием соответствующих химических веществ, растворенных в воде. Если эти вещества присутствуют в морской воде, то организм ракушки каким-то образом маркирует соответствующие гены (в половых клетках). Какие-то гены включаются, какие-то, наоборот, выключаются. Таким образом, программа индивидуального развития моллюска становится несколько другой, и в результате получается несколько другая ракушка, с несколько иным спектром морфологических и экологических признаков. И поскольку соответствующие гены уже «отмаркированы», то такие (отмаркированные) гены будут передаваться потомству. То есть, соответствующие изменения морфологии и экологии станут наследственными. Таким образом, мы получим долговременную модификацию этого вида, которая будет устойчиво воспроизводиться в череде поколений, пока условия окружающей среды снова не вернутся к исходным.
Интересно, что характерными признаками подобного эпигенетического перехода должны являться, во-первых, быстрота этого перехода, а во-вторых, его массовость. То есть, сразу множество особей этих ракушек должны дружно продемонстрировать соответствующие изменения, причем это должно произойти быстро. Например, всего за 10-15 поколений. Потому что определенные химические вещества будут воздействовать на уже имеющиеся (соответствующие) гены сразу множества особей моллюсков. Таким образом, в случае подобного сценария генетических изменений, мы должны наблюдать именно такую картину «эволюции ракушек», которую и увидели реально авторы данной работы (массовые и быстрые изменения моллюсков).
В данном случае не представляется возможным как-то разделить, проверить и выбрать только один из двух возможных теоретических сценариев – «эволюцию вследствие естественного отбора» или же «эпигенетический переход между разными модификациями». Это можно сделать только в ходе дополнительных специальных исследований. А пока и «эволюция под действием естественного отбора», и «эпигенетический переход» являются вполне возможными сценариями. Действительно, эволюция за 10-15 поколений, хотя и с трудом, но всё же может быть обеспечена естественным отбором. Конечно, при условии, что соответствующее давление естественного отбора было очень сильным (если соответствующие признаки давали общее преимущество в выживании в пределах 200-300%).
Ниже я приведу такие примеры, где объяснение произошедшей «эволюции» именно давлением естественного отбора – уже точно не подходит (поскольку «эволюция» там произошла всего за 4 поколения). Но конкретно в случае аральских ракушек, сценарий с «эволюцией под действием естественного отбора» всё-таки является одним из возможных.
С другой стороны, столь же возможен и «эпигенетический переход» - эпигенетические эффекты сегодня уже достаточно известны, и даже неплохо изучены. Кроме того, как я только что сказал, сегодня известны и такие примеры «эволюции», которые уж точно «не влазят» в версию естественного отбора (см. ниже).
Авторы работы не показали, и даже не ставили себе такую задачу – разобраться, какой конкретно из возможных механизмов изменений здесь имел место – эволюция ли под действием естественного отбора? Или же эпигенетические изменения? Или вообще, банальная модификационная изменчивость?
Поскольку никаких комментариев к работе Андреевых, опубликованных в научной печати, лично мне найти не удалось, то в заключение приведу несколько высказываний с соответствующего (профильного) форума:
mastax писал: [20]
Андреевы начали заниматься форменной подтасовкой фактов, считая, что если моллюск питается детритом, то он детритофаг (детритом питаются еще и сейстонофаги в условиях, когда детрита много на поверхности). То, что у двустворчатых моллюсков один и тот же вид может занимать разные экологические ниши – это тоже известно. При этом форма раковины у них меняется адекватно условиям обитания. А в ленинградской школе малакологов вид двустворок определяют по характеру выпуклости створок (более нигде в мире такую методику не используют), поэтому нет ничего удивительного, почему в том же Арале они нашли так много нео-видов.
Gilgamesh писал: [21]
Mastax, а какой таксономический ранг считают более вероятным для новых аральских форм малакологи других научных школ?
mastax писал: [22]
Прошу прощения, что не ответил сразу: ни в каких. Т.е. все понимают, что это не новые виды и не подвиды. Если бы Андреевы показали, что у этих форм меняется число хромосом или строение спермиев (у солоноватоводных кардиид спермии исключительно видоспецифичны благодаря винтообразной форме - это было показано Карпевич), тогда еще можно было бы о чем-то говорить. А так иностранцы в очередной раз покрутили пальцем у виска и даже не стали переводить свои мысли вслух. Если кто знает немецкий и имеет доступ к хорошей библиотеке, то может прочитать вот эти статьи:
Reischü tz, P. L. Beiträ ge zur Molluskenfauna Niederö sterreichs, XI. Ist Starobogatovismus die Steigerung von Bourguignatismus? // Nachr. Bl. Erste Vorarlberger Malakol. Gesell. 1994. Bd 2. S. 51–52.
Meier-Brook C. Artaufassungen in Bereich der limnischen Mollusken und ihr Wahd im 20 Jahrhundert // Arch. Moll. 1993. Bd 122. S. 133–147.
Марков Александр писал: [23]
Довольно давно я обещал разузнать у специалистов из нашего института, почему они считают, что изменения у моллюсков в высыхающем Арале не имеют отношения к видообразованию. К сожалению, четких ответов мне получить не удалось. Но в общем ситуация примерно такая. Известно, что в стрессовых условиях онтогенез может нарушаться, возникают изменения дегенеративного характера. Это не видообразование, а просто уродства, вызванные неприемлемыми внешними условиями. Тому есть много примеров (например, в популяциях планктонных простейших, попавших с течениями из пресных вод в солоноватые, или наоборот, происходят подобные явления; кончается все гибелью популяций, в этих изменениях нет ничего адаптивного). По мнению Л.А. Невесской, Андреевы не представили достаточных доказательств того, что наблюдаемые в Арале изменения являются не такими вот дегенеративными, а именно видообразовательными, что там имеет место возникновение каких-то новых наследуемых адаптаций.
Вопрошающий писал: [24]
…недостаточность аргументации могу обосновать: 1) Детерминантность (важный критерий!) исходных особей к мутантным (?), насколько мне известно не доказана. А почему? Ведь Андреевым, там, на месте, это было совсем нетрудно сделать. 2) Чтоб понять мутация это или просто болезнь, достаточно было поместить нетипичные особи в воду нормальной (исходной), или промежуточной солености, и посмотреть какое будет потомство. Почему этого не сделали? 3) Просто сравнивать ракушки = что сравнивать скелеты обычных чел со скелетами йододефицитных кретинов. Вывод о факте видообразования будет весьма и весьма сомнительным. Согласитесь, на фоне титанической работы Андреевых, отсутствие столь простеньких исследований выглядит более чем странным.
Amage писал: [25]
Вид - это очень тонкая категория. Определения вида нет, кроме одного, что им следует считать то, что считает таксономист по группе. Поэтому то, что один считает видом, другой может таковым не считать и возможности переубедить друг друга у них нет. Обычно такие споры не возникают просто из-за того, что таксономистов на Земле и тысячи не неберётся и одной группой занимается один, а большинством - никто.
Меня убеждает, например, аргументация Андреевых о видообразовании в Арале. Поэтому я полагаю, что «достоверные факты свершившегося образования нового вида (в настоящем) наукой установлены». Другого их аргументы могут и не убедить и он даст другой ответ на Ваш вопрос…
Amage писал: [26]
Марков говорил, что малакологи ПИН неофициально высказывали сомнения. Меня они не убеждают (защита Андреевой была у нас на совете) и я сам таксономист. Более того, если посмотреть на эволюцию моллюсков понто-каспия по данным той же Невесской, то получается абсолютно то же самое, что получилось в Арале.
Итак, мы видим, что «взрывное видообразование аральских моллюсков» принимается далеко не всеми. Одних специалистов оно убеждает, другие специалисты к этим исследованиям относятся крайне негативно. Пусть читателей не смущают столь резкие формулировки некоторых биологов. Среди специалистов бывают споры и похлеще. Скорее всего, Андреевы совсем не занимались «подтасовкой фактов», а провели совершенно нормальное исследование. Причем исследование уникальное, поскольку повторять «на бис» своё усыхание Арал пока не собирается.
Тем не менее, очевидно, что этот пример «доказательства эволюции под действием естественного отбора» (так же как и «свершившегося видообразования») настолько далек от доказанности, насколько вообще может быть далеко что-либо от чего-либо.
3. ЕЩЕ ПРИМЕРЫ «НАБЛЮДАЕМОЙ ЭВОЛЮЦИИ», С КОТОРЫМИ К ВАМ МОЖЕТ ПРИСТАТЬ ВЕРУЮЩИЙ ДАРВИНИСТ
Итак, мы подробно разобрали примеры « наблюдаемой эволюции» улиток и ракушек. Теперь приведём некоторые другие примеры «наблюдаемой эволюции», с которыми к Вам может пристать верующий дарвинист. В этом отношении у верующих дарвинистов еще популярны (в порядке убывания популярности):
1. Эволюционирующие бактерии.
2. Знаменитые тли Шапошникова.
3. Эволюционирующие ящерицы в Америке.
4. Эволюционирующие жабы в Австралии.
Начнем с конца. То есть, с эволюционирующих жаб.
БЫСТРОНОГИЕ ЖАБЫ
Этот «пример эволюции», наверное, меньше всего известен верующим дарвинистам. Во всяком случае, лично мне его привели только один раз. Но поскольку всё-таки привели, то придётся разобрать и его тоже.
Итак, в 2006 году австралийские биологи в соответствующей статье поведали миру о том, что жабы-аги, ввезённые в Австралию более 70 лет назад, в настоящее время стремительно эволюционируют (Phillips et al., 2006).
История началась с того, что в 1935 году жабы-аги были завезены в Австралию для борьбы с вредителями посадок сахарного тростника. К сожалению, жаба-ага не оправдала надежд австралийских земледельцев. Её роль в истреблении вредителей «сахарных полей» оказалась более чем скромной. Зато эта жаба стала успешно размножаться на новой территории, и постепенно распространяться от места вселения на новые площади.
Авторы сравнили темпы экспансии жабы-аги в разные годы, и оказалось, что скорость её распространения по территории Австралии в последнее время намного выше (примерно в 5 раз), чем скорость её распространения в первые годы экспансии. На этом основании авторы предположили, что сама способность расселяться - у этих жаб эволюционировала.
Чтобы проверить эту версию, авторы измерили скорость передвижения жаб по земле, и выяснили, что более длинногие жабы передвигаются на большие расстояния, чем коротконогие. Потом исследователи обмерили жаб, находящихся на переднем крае экспансии, и сравнили их с жабами, живущими уже в «глубоком тылу». Исследователи выяснили, что в среднем, жабы-первопроходцы (взятые с переднего края распространения) являются более длинноногими (и соответственно, более резвыми), чем жабы из тех районов Австралии, которые были «завоеваны» ими уже давно.
Вот на этом основании австралийские исследователи и сделали вывод, что жабы… эволюционировали.
Что и говорить, этот вывод исследователей не может не вызвать улыбку. Такое ощущение, что эволюцию сейчас поминают вообще по любому поводу. Впрочем, и сама по себе эта история довольно забавная – стоит только представить себе этих жаб, упрямо марширующих вглубь Австралии в поисках светлого будущего. Поневоле вспоминаются американские переселенцы, штурмовавшие американский запад на своих фургонах. Причем первыми до побережья Тихого океана добирались либо самые длинноногие переселенцы, либо те, у кого фургоны оказались крепче. На этом основании можно заключить, что в ходе освоения Америки происходила эволюция фургонов и длинноногих переселенцев.
Действительно, мы знаем, что осваивать новые территории в Америке в своё время отправились далеко не все имевшиеся европейцы, а только некоторая их часть. Наверное, те, которые (от природы?) были склонны к подобным приключениям и авантюрам. Вот эта часть популяций европейцев и оказалась в Америке. А затем, еще более «подвижная» часть этих людей двинулась еще дальше – на запад. Таким образом, в конце этого пути весь «дикий запад», наверное, состоял именно из таких людей, которые были склонны к подобному поведению. Во всяком случае, значительная доля этих людей. Можно ли на этом основании заключить, что люди в этот момент проэволюционировали?
Вместе с людьми на запад катились еще и фургоны. И естественно, те фургоны, которые были крепче, укатились дальше, чем те фургоны, которые были менее крепкими. Можно ли на этом основании заключить, что и фургоны тоже проэволюционировали (вместе с людьми)?
Если я рассыплю по полу шарики разного веса, то более легкие шарики укатятся дальше, чем более тяжелые шарики. И возможно, закатятся даже в такие закоулки, из которых их мне потом выковыривать и выковыривать… Могу ли я на этом основании заключить, что мои шарики проэволюционировали, раскатившись по полу?
Вот так же и авторы данного исследования. Спустя 70 лет после вселения жабы-аги в Австралию, они измерили ноги этих жаб на самом переднем крае расселения, и выяснили, что эти ноги, в среднем, несколько длиннее, чем ноги «отставшей» части популяции жаб… и заключили, что ноги жаб эволюционировали. Но ведь здесь можно всё объяснить гораздо проще – что на переднем крае экспансии такие (длинноногие) жабы оказались именно потому, что они были способны передвигаться на более значительные расстояния, чем жабы с короткими ногами. То есть, исходные (смешанные) популяции жаб в ходе расселения по Австралии просто рассортировались на жаб с длинными ногами и жаб с короткими ногами. Точно так же, как и американские переселенцы в ходе расселения по Америке рассортировались на людей, более склонных к авантюрам, и людей, менее склонных к подобным вещам.
Примерно то же самое отмечают и австралийские исследователи. С 2006 года они отнюдь не прекратили изучать жаб-переселенцев, а продолжили свои наблюдения, и в более поздней статье доложили, что жабы-аги на переднем крае экспансии демонстрируют еще и поразительное поведение. А именно, жабы-первопроходцы оказались склонны очень долго и упорно двигаться строго в определенном направлении, в результате чего преодолевали за ночь такие расстояния, которые беспрецедентны для других амфибий (Phillips et al., 2007; Alford et al., 2009). Точь в точь, как американские переселенцы, упорно продвигавшиеся на запад за своей «американской мечтой».
Понятно, что подобные «марш-броски» для этих жаб даром не проходят. За склонность к столь затяжным ночным марафонам жабы-авантюристки расплачиваются повышенной частотой заболеваний, например, артритом позвоночника (Shilton et al., 2008). А также демонстрируют более слабую иммунную систему в целом (Llewellyn et al., 2012). Эти печальные обстоятельства компенсируются более качественным питанием, которое жабы, кажется, получают, первыми прибыв на новое место (Brown et al., 2013).
Попутно исследователи выяснили, что во всё увеличивающейся скорости распространения этим жабам неплохо помогают… разнообразные дороги (в том числе, автомобильные), по которым жабы-аги передвигаются значительно быстрее, чем по траве (Brown et al., 2006). Не знаю, как с этим обстоят дела в Австралии, но в России за последние 70 лет сеть автодорог, кажется, всё-таки немного «проэволюционировала». Таким образом, можно предположить, что в повышении скорости распространения жаб по Австралии виновна не только «эволюция жаб», но еще и «эволюция дорожной сети» за последние 70 лет.
Еще исследователи выяснили, что жабы-аги могут разъезжать по стране с перевозимыми на машинах стройматериалами (а также с материалами по озеленению) в качестве безбилетных пассажиров (White & Shine, 2009). Например, число жаб, достигших именно таким образом города Сидней, было очень высоким (White & Shine, 2009). Поэтому можно сделать вывод, что «эволюции» скорости распространения жаб способствовала еще и параллельная «эволюция грузоперевозок» в этой стране, которая за последние 70 лет, тоже, наверное, выросла.
Что еще можно добавить к этому примеру «эволюции жаб»?
Пожалуй, только то, что спустя несколько лет после публикации первой статьи, австралийские биологи, наконец, и сами догадались, что найденный ими «пример эволюции» - не слишком похож на ту эволюцию, которая обычно имеется в виду, когда о ней говорят дарвинисты. Исследователи опубликовали сразу две работы на эту тему (Shine et al., 2011(а); Shine et al., 2011(б)), где они приходят к следующему выводу:
…В отличие от обычной эволюции, которую ведет естественный отбор, мы думаем, что ускоренное вторжение жабы следует из «пространственной сортировки». Гены, нужные для того, чтобы перемещаться быстрее и дальше, концентрируются на всё более стремительном фронте вторжения. [27]
То есть, авторы буквально повторяют то, о чем я выше уже рассказал на примере «американских переселенцев». Это не эволюция (в том смысле, в котором этот термин общепринято употребляется), а просто пространственная дифференцировка соответствующих генов. Где гены, способствующие быстрому расселению, в итоге оказались в соответствующем месте – на переднем крае этого расселения. Причем эти гены, скорее всего, уже имелись в генофонде этого вида. Просто в результате экспансии они постепенно сконцентрировались именно там, где и должны были сконцентрироваться.[28]
В заключение осталось только посмотреть, чем же подобные «эволюции» заканчиваются. А вот чем.
В очередной статье на эту тему (Lindstrom et al., 2013) австралийские биологи исследовали этот вопрос, и выяснили, что дальность перемещения жаб на тех участках, где «волна расселения» уже прошла (всего несколько лет назад) опять снижается. Отсюда следует, что «эволюции быстроногости» у жаб-аг, скорее всего, не происходит. После прохождения волны экспансии «всё возвращается на круги своя».
И наконец, последнее. Всё-таки не исключено, что механизмы изменения подвижности жаб именно в самой «волне расселения», на самом деле, более сложные, чем просто «пространственная сортировка генотипов». Может быть, здесь задействованы специальные генетические механизмы, которые у этих жаб обычно «спят», а включаются только в новых условиях (например, при расселении). Эти включившиеся гены и переводят генотипы некоторой части особей в особый «режим расселения». В результате чего жабы и начинают демонстрировать то странное поведение, которое заставляет их упорно двигаться в одном направлении, в результате чего дальность такого «марш-броска» может составить до одного километра за ночь. А после расселения, соответствующие генетические комплексы опять засыпают, и жабы возвращаются к добропорядочному образу жизни.
Такой сценарий звучит несколько фантастично… пока не узнаешь о еще более фантастических вещах, демонстрируемых некоторыми другими животными в ходе аналогичных расселений на новом месте. Чуть ниже я расскажу о настолько невероятных «выходках», которые продемонстрировал конкретный вид жука (зигограмма), попав в новые местообитания, что по сравнению с ним «жабы-марафонцы» начинают выглядеть совсем бледно. Читайте об этом ниже.
Впрочем, может быть, в описанном механизме и нет ничего фантастического. Известно, что у многих животных при достижении некоего критического порога плотности популяции, включается миграционное поведение. Наверное, самые известные примеры подобных вспышек стремления попутешествовать – это саранча и лемминги. Можно предположить, что и у этого вида жаб тоже имеется что-то подобное – стремление путешествовать «включается» у определенных особей при достижении популяциями некоего порога плотности. Может быть поэтому в первые годы появления жабы-аги в Австралии, когда плотность их популяций была меньше, чем сейчас, скорость их расселения тоже была меньше, чем сейчас? [29]
ЯЩЕРИЦЫ И НЕДОРАЗУМЕНИЯ
Исследования с «эволюционировавшими ящерицами» гораздо более разрекламированы, чем пример с «эволюционировавшими жабами». Причем в отличие от одинокого примера жаб из Австралии - по ящерицам таких исследований опубликован целый ряд, и выполнены они разными авторами на разных ящерицах. Поэтому нам сейчас придётся «разгрести» все эти примеры «эволюции».
Во-первых, в некоторых ссылках на тему «эволюционировавших ящериц» могут быть опубликованы просто недоразумения. Приведу один, недавно установленный пример, в котором якобы произошла сверхбыстрая «эволюция» ящериц, заселённых сразу на несколько островков Карибского моря (Kolbe et al., 2012). В этом исследовании биологи поселили на семь (7) маленьких островков Багамского архипелага по паре ящериц с соседнего большого острова и четыре года (4 года) наблюдали за «эволюцией» их потомства.
Результаты исследований:
На всех (!) семи островках у ящериц синхронно укоротилась средняя длина ног примерно на 5% (у самцов на 6.5%, у самок на 4%) в течение всего 3 (!) поколений.
Вдохновленный столь сногсшибательными результатами, автор соответствующей научно-популярной заметки (Марков, 2012а) принялся рассуждать о какой-то там эволюции (за три поколения!) под действием естественного отбора, в ходе приспособления к более тонким веткам растений, растущим на этих островках, в сравнении с более толстыми ветками, растущими на исходном острове.
На самом деле, понятно, что такие результаты вообще не лезут ни в какие ворота представлений о дарвиновской эволюции. Во-первых, в рамках дарвинизма, изменения организмов случайны, следовательно, синхронность изменений (произошедших на всех семи островках за три поколения) автоматически опровергает механизм «случайные мутации + естественный отбор». Особенно если помнить, что каждый островок заселялся всего одним самцом и самкой. Таким образом, здесь для естественного отбора был вообще наименьший исходный материал, который только возможен.
Во-вторых, 3 поколения - слишком мало для того, чтобы новый признак завоевал популяцию. Даже если этот признак чудовищно полезен, например, повышает приспособленность аж на 400%, то даже в этом случае вряд ли стоит ожидать замены всех ящериц на более коротконогих всего за 3 поколения.
Тем более что на самом деле, естественный отбор в обсуждаемом случае совсем не был «чудовищным». Скорее уж, наоборот:
1) Ящерицы расселялись по островкам, на которых вообще отсутствовали их природные хищники.
2) Ящерицы данного вида не являются строго древесными - они могут сидеть не только на ветвях, но и на стволах растений (которые, конечно, толще, чем ветки), включая самое основание этих стволов, и даже вообще на земле (Рис. 9). То есть, эти ящерицы в обычных условиях (взятые оттуда, откуда их привезли) спокойно выживают со своими ногами: 1) на земле, 2) на нижней части стволов, 3) на тонких и толстых стволах и ветках (растительность исходных островов включает растения разной толщины).
3) Наконец, вряд ли укорочение длины конечностей на 5% так критически влияло на выживание ящериц даже на тонких ветвях. Трудно представить себе эту ящерицу, так хорошо приспособленную к древесному образу жизни… хронически падающей с веток из-за того, что её конечности чуть длиннее, чем у других.

Рисунок 9. Ящерица коричневый анолис (Аnolis sagrei), объект исследования в обсуждаемой работе ( Kolbe et al ., 2012) в своей естественной среде обитания: на тонких ветвях, на стволе дерева, и на земле.
Более того, соответствующие исследования (Jones & Jayne, 2012), наоборот, установили, что виды ящериц именно из рода Anolis с более длинными задними ногами передвигаются быстрее по любой поверхности (независимо от её толщины), чем их менее длинноногие собратья из этого же рода (в этой работе изучалась скорость передвижения ящериц по веткам диаметром: 5 мм, 10 мм, 30 мм, 60 мм, 100 мм).
Наконец, в других исследованиях, с другим видом ящериц (Husak, 2006) было установлено, что ящерицы во время охоты обычно развивают лишь половину той скорости, которую они развивают, стремясь избежать хищника. Таким образом, скорость передвижения по веткам, наверное, была не слишком актуальна для ящериц, оказавшихся на острове, где хищники отсутствовали вовсе.
В общем, совершенно ясно, что механизмы изменений здесь были не дарвиновскими. Даже если бы и имелась какая-то микро-причина для укорочения ног ящериц (хотя непонятно, какая), то естественный отбор за 2-3 поколения невозможен. Для того чтобы за 2-3 поколения на всех 7 островках получилось то, о чём рапортовали в статье исследователи - вместо естественного отбора на этих островках должен был сидеть разумный селекционер, который бы сортировал этих ящериц.
В связи с этим, в первую очередь, сразу же приходит на ум концепция номогенеза, предложенная нашим российским биологом Львом Семёновичем Бергом – «эволюция на основе закономерностей» (Берг, 1922). Потому что именно эта концепция постулирует одновременные (синхронные) эволюционные изменения живых организмов (под действием неких, не слишком понятных законов), затрагивающие сразу большое число особей на какой-то территории в целом. Получается, что обсуждаемое исследование с ящерицами - буквально «тютелька в тютельку» подтверждает именно постулаты номогенеза (одновременно опровергая дарвиновские механизмы эволюции).
И об этом следует сказать Вашему верующему дарвинисту. Потому что верующему дарвинисту лучше повеситься, чем признать номогенез справедливым, а дарвинизм - ошибочным. Особенно потому, что в номогенезе постулируется эволюция под действием неких неизвестных законов (или даже сил)... что может быть истолковано весьма неудобным (для атеистического мировоззрения) способом.
Однако в данном случае привлекать для объяснения полученных результатов номогенез вряд ли стоит, потому что можно предложить гораздо более простое объяснение.
Во-первых, следует (опять!) вспомнить, что длина ног конкретной особи - это признак, подверженный внутривидовой изменчивости.
Во-вторых, рассмотрим другое исследование (Husak, 2006), где изучалось влияние скоростных качеств ящериц Crotaphytus collaris и длины их задних ног на шансы выживания этих ящериц.
В этом исследовании получилось, что скоростные качества и длина задних ног не влияют на выживание взрослых ящериц Crotaphytus collaris. Но зато эти параметры влияют на выживание «младенцев» ящериц (Таблица 1):

Таблица 1. Из работы (Husak, 2006), суммирующая результаты выживания ящериц Crotaphytus collaris в зависимости от их возраста и длины задних ног.
Как видно из таблицы, выжили молодые ящерицы со средней длиной задних ног 54.83 мм, а погибли ящерицы со средней длиной ног 51.39 мм. Понятно, что такое избирательное (по длине ног) выживание немного смещает исходную среднюю норму длины задних ног в сторону их увеличения (примерно на 6%).
То есть, допустим, имеется разброс длины ног среди новорожденных в пределах 30%. Однако хищники за год уничтожают слишком коротконогих и медлительных. В результате популяция поддерживается в «спортивной форме» (из года в год).
И вот теперь, если данную популяцию каким-то образом переместить в такие условия, где полностью отсутствуют хищники, давление естественного отбора на слишком коротконогих полностью исчезнет, и выживание ящериц в этом отношении примет случайный характер. В таких (новых) условиях, средняя длина задних ног ящериц очень быстро сместится обратно в сторону некоторого укорочения ног. Потому что даже самые коротконогие ящерицы в новых условиях будут тоже выживать вполне успешно.
В диссертационной работе этого же автора (Husak, 2005) был установлен весьма интересный факт - что ящерицы ловят свою добычу, не слишком напрягаясь. Только нападение хищника заставляет их развивать максимально возможную скорость. А при ловле добычи, скорость, развиваемая ящерицами, примерно в 2 раза ниже. Цитата из статьи (Husak, 2006):
Молодежь и взрослые бегут в среднем между 0.8 и 1.3 м/c, добывая продовольствие, и 2.0 и 2.5 м/c - убегая от хищников (Husak, 2005).
Если озвученные факты, полученные в отношении ящерицы Crotaphytus collaris, примерно справедливы и для обсуждаемого нами вида Anolis sagrei, тогда получается, что таинственная и сверхбыстрая «эволюция» укорочения ног ящериц, не более, чем недоразумение. Недоразумение, основанное на том, что на родном острове анолисов в присутствии хищников выживали не все ящерицы. Самых медлительных и коротконогих, в среднем, съедали чаще. А когда анолисы попали на островки без хищников, фактор быстроногости стал совсем не актуальным, и выживать стали все (даже самые коротконогие) сразу на всех семи островках. Соответственно, средняя длина ног ящериц немного снизилась тоже синхронно на всех семи островках. Интересно, что в случае Crotaphytus collaris, хищничество как раз смещает (в сторону длинноногости) молодежь данных ящериц именно на те самые 6%, которые близки к 5% укорочения средней длины ног, полученной в обсуждаемом исследовании.
Понятно, что всё это пока лишь предположения, которые надо проверять. Например, непонятно, откуда взялось разнообразие по длине ног у ящериц, размножившихся за три поколения всего от одной пары. Но точно такой же вопрос пришлось бы задать и версии с естественным отбором. Естественный отбор из чего? Из непонятно откуда взявшегося разнообразия длины ног?
Поэтому, наверное, всё-таки не стоит умножать сущности без необходимости, объясняя сверхбыстрой и синхронной эволюцией то, что может быть объяснено совершенно прозаичными причинами - потерей «спортивной формы» ящериц в отсутствие хищников.
Итак, самое «эволюционное», что мы можем вытащить из приведенного примера «эволюции ящериц» - это действие стабилизирующего отбора.[30] Когда норма, характерная для того или иного вида, поддерживается не только внутренними границами изменчивости самого вида, но и внешними факторами среды. Например, хищничеством. В частности, у обсуждаемых ящериц размах внутривидовой изменчивости по длине задних ног оказался достаточно широким. Но хищники уменьшают этот размах изменчивости, поедая (чаще) самых коротконогих. Таким образом, хищники (там где они есть) поддерживают обсуждаемых ящериц в «спортивной форме», отбраковывая выбивающихся «из ряда вон» особей.
Дата: 2019-02-02, просмотров: 511.