Рассмотрим еще один пример «эволюционировавших ящериц» (Losos et al., 2006).
Здесь авторы исследования подселили на шесть островков с обитавшими там ящерицами Anolis sagrei, хищных ящериц Leiocephalus carinatus. А еще шесть островков, где жили Anolis sagrei, но хищных ящериц там не было, взяли в качестве сравнения (контроль).
Уже через полгода (шесть месяцев) авторы исследования обнаружили, что численность бедных Anolis sagrei, к которым подселили хищников, упала примерно вдвое. И еще исследователи обнаружили, что выжившие анолисы имеют, в среднем, более длинные задние ноги, чем до появления хищников. Очевидно, что прибывшие хищные ящерицы (Leiocephalus carinatus) устроили настоящий террор среди бедных анолисов (за шесть месяцев съели примерно половину). И заставили этих анолисов хорошенько побегать. При этом более скоростные анолисы (с более длинными задними ногами), очевидно, получили некоторое преимущество, в результате чего и получилось, что выжило больше длинноногих анолисов. Однако спустя еще полгода... всё вернулось на круги своя. Анолисы перебрались на ветки растений... и средняя длина ног ящериц вернулась обратно (к более коротким значениям).
Что мы можем извлечь уже из этого (очередного) «примера наблюдаемой эволюции»?
Прежде всего ясно, что здесь ни о какой эволюции вообще не может быть речи. За шесть месяцев в этой популяции не сменилось ни одного (!) поколения ящериц. Все озвученные изменения произошли в пределах (внутри) одного поколения. Поэтому данный «пример эволюции» автоматически выбывает из примеров эволюции.
Всё, что мы здесь наблюдаем - это естественный отбор анолисов под давлением прибывших хищников. За первые шесть месяцев естественный отбор (в лице хищников) «вырезал» из обсуждаемого поколения анолисов преимущественно менее скоростных, оставив преимущественно более длинноногих (из всё того же состава ящериц). Спустя еще шесть месяцев, анолисы перебрались на ветки растений, и средняя длина ног выживших ящериц опять укоротилась. Авторы исследования объясняют это тем, что с короткими ногами удобнее лазить по тонким ветвям. Правда, такое предположение авторов уже успело слегка устареть, потому что (как я уже говорил выше) недавнее исследование (Jones & Jayne, 2012) показало, что более длинноногие виды анолисов передвигаются быстрее по любым веткам (хоть толстым, хоть тонким). Поэтому последний результат авторов (с естественным отбором более коротконогих анолисов на тонких веточках) не слишком понятен.
Однако в любом случае ясно, что ни о какой «наблюдаемой эволюции» в данном исследовании говорить вообще не приходится. Ибо внутри рамок единственного поколения, никакой эволюции не может быть просто по определению.
Отмечу, что и само поведение анолисов - тоже не изменилось (в рамках вида). Anolis sagrei - это такой вид ящериц, который способен обитать и на земле, и на толстых стволах деревьев, и на тонких ветвях.
Поэтому если дарвинист в ответ на Вашу просьбу привести строго установленный пример эволюции видового морфологического признака начнет забрасывать Вас «эволюционировавшими ящерицами» из перечисленных выше опубликованных работ, Вы должны объяснить дарвинисту, что собственно эволюции ящериц в приведенных примерах не наблюдалось. Будут ли задние ноги ящериц Anolis sargei (в условиях отсутствия хищников) продолжать укорачиваться до тех пор, пока не выйдут за границы всех ранее известных значений? Или наоборот, в присутствии хищников - начнут ли ноги анолисов бесконечно удлиняться? Это, как говорится, вилами по воде писано (в обсуждаемых исследованиях не показано). А может быть, удлинение или укорочение ног - быстро упрётся в какие-то пределы, характерные для этого биологического вида?
«Есть ли жизнь на Марсе, нет ли жизни на Марсе - это науке неизвестно» (С)
То есть, мы опять не имеем примера строго зафиксированной эволюции форм под действием естественного отбора (не имеем примера эволюционного выхода какого-либо морфологического признака за пределы всех наблюдавшихся ранее значений, известных для данного вида).
Кстати, по поводу возможных «лимитов эволюции». Интересно, что в недавнем исследовании (Logan et al., 2012) авторы исследовали две изолированные популяции уже другого вида анолиса (Anolis lemurinus) на предмет морфологических различий между ними.
Эти изолированные популяции интересны тем, что они находятся на двух островках архипелага Кайос Качинос (Гондурас) - Кайо Менор и Кайо Майор. Эти островки расположены друг от друга на расстоянии 2.5 км. А от побережья материка оба острова удалены уже на 18 км.
Предполагается, что Anolis lemurinus попали на эти два островка, как минимум, 10.000 лет назад. А потом (уже с этих островов) попали на еще более удаленные острова (Bay Islands) и заселили и их тоже. В предыдущих исследованиях было установлено, что на одном из этих удаленных островов (о. Утила) популяция анолисов в настоящее время морфологически весьма существенно отличается от материковых популяций данного вида (Klutsch et al., 2007). Однако на исходных островках архипелага Кайос Качинос (послуживших «плацдармом» для захвата других островов), ящерицы этого вида оказались морфологически и генетически неотличимы от материковых популяций (Klutsch et al., 2007). Что весьма странно, поскольку изоляция анолисов на островках Кайос Качинос (теоретически) продолжалась самое долгое время.
Этот парадокс, видимо, заинтересовал других исследователей (Logan et al., 2012) и они решили исследовать популяции двух разных островков архипелага Кайос Качинос на предмет морфологических отличий более тщательно, чем в предыдущем исследовании. Тем более что авторы пишут - несмотря на близость друг к другу, островки Кайо Менор и Кайо Майор довольно серьезно различаются между собой условиями обитания. Во-первых, на острове Кайо Майор в изобилии произрастают пальмы рода Attalea, что серьезно меняет структуру растительности, а на острове Кайо Менор этого не наблюдается. Во-вторых, на острове Кайо Майор - больше разнообразие хищников (млекопитающих и змей), опасных для анолисов. Например, Rattus rattus (крыса), Dasyprocta punctata (агути), Leptophis mexicanus (змея) живут на Кайо Майор, но на Кайо Менор отсутствуют. Наконец, на Кайо Майор имеется постоянный источник пресной воды, что должно уменьшать «водный стресс» для анолисов (Logan et al., 2012).
В связи с этим, авторы обсуждаемой работы, во-первых, решили посмотреть более внимательно, имеются ли морфологические различия между двумя разными популяциями. А во-вторых, имеются ли различия в использовании среды изолированными популяциями ящериц этих двух островков. Ведь как никак, 10.000 лет прошло. Исследователи проделали следующую работу:
1) Измерили 10 разных параметров окружающей среды. В том числе, высоту и диаметр ветвей и стволов, на которых находили ящериц в разных популяциях.
2) Измерили 12 разных морфологических параметров ящериц, пытаясь найти различия между двумя популяциями. Или хотя бы различия в половом диморфизме между двумя популяциями (самки и самцы на обоих островках измерялись, конечно, раздельно).
Результаты исследований:
Оказалось, что ящерицы из двух изолированных популяций имеют некоторые мелкие различия в морфологии и половом диморфизме. В частности, у самцов анолисов с острова Кайо Менор наблюдалось немного лучшее состояние тела, немного более длинные задние ноги и вырост под горлом (dewlap) немного меньшего размера. Однако никаких различий в использовании среды (из измеренных 10 параметров) между разными популяциями ящериц - выявить не удалось. Интересно, что чуть более длинные задние ноги были установлены для популяции, в которой хищническое давление было меньшим (Кайо Менор), и при этом различий в высоте и диаметре веток и стволов (с которых снимали данных ящериц) выявить не удалось. Казалось бы, в таких условиях теория (основанная на результатах исследований, обсуждавшихся выше) предсказывает нам укорочение ног... Ан нет. Задние ноги ящериц Кайо Менор оказались, наоборот, чуть-чуть длинее. Авторы пишут (стр. 220):
...Длина задних ног часто положительно коррелирует с диаметром (ветвей), так как это обеспечивает биомеханическое преимущество (Caksbeek et al. 2007., Irschick & Losos 1999, Langerhans et al. 2006). Хотя самцы с острова Кайо Менор имели более длинные задние ноги, чем самцы с острова Кайо Майор, мы не нашли различий в использовании веток между этими популяциями.
В итоге, авторы исследования заключают, что те мелкие морфологические отличия, которые они установили, вряд ли обусловлены различиями в условиях среды. Кроме того, по причине «микроскопичности» отличий (накопившихся за 10.000 лет) авторы предполагают, что они в данном случае столкнулись со случаем «эволюционного консерватизма».
И здесь авторов можно понять. Действительно, когда читаешь про «бешеную эволюцию анолисов» за три (!) поколения... а потом читаешь еще и про «эволюцию анолисов» за одно (!) поколение... то складывается ощущение, что через 10.000 поколений у этих «бешено эволюционирующих анолисов», наверное, ноги станут длиннее, чем у кенгуру... или наоборот (при обратном давлении среды), ноги укоротятся совсем, атрофируются, а сами ящерицы удлинятся, зашипят и превратятся в змей... Но стоит только посмотреть на эту «бешеную эволюцию» с расстояния в 10.000 лет... как куда что девается - различия между популяциями, во-первых, оказываются минимальными, а во-вторых, влияние среды оказывается вообще не при чём.
Авторы пишут:
Эта нехватка расхождений в использовании среды обитания особенно поразительна, учитывая очевидные различия в структуре растительности между островами. Схожесть между популяциями может быть продуктом консерватизма экологической ниши (Holt & Barfield 2008, Losos et al. 2003, Warren et al. 2011), филогенетических ограничений (Harvey & Pagel 1991), или потока генов (Klutsch et al. 2007, Lenormand 2002).
Мы видим, что авторы пишут и о «консерватизме экологической ниши» (то есть, по сути, о консерватизме биологического вида). И о «филогенетических ограничениях». А ведь в переводе на обычный язык, «филогенетические ограничения» как раз и означают те самые границы изменчивости вида, о возможности существования которых мы говорили выше. Ну а что касается обмена генами между популяциями, то это возможно, но всё же «поток генов» был, наверное, не слишком интенсивным, чтобы совсем уж разрушать эффекты изоляции. Трудно представить себе древесных ящериц анолисов, регулярно штурмующих 2.5 км океана, чтобы наведаться в гости к соседям. Еще труднее представить этих же ящериц, штурмующих уже 18 км океана, чтобы «обменяться генами» еще и с материковыми популяциями ящериц, от которых они, судя по проведенным исследованиям, тоже мало отличимы (Klutsch et al., 2007).
Итак, давайте подчеркнем еще раз это важное обстоятельство:
1) Смотрим на исследования по «галопирующей эволюции ящериц» (за одно или три поколения) и нам кажется, что «эволюция правит миром»...
2) «Отодвигаемся» на 10.000 лет... и не видим никакой эволюции. А видим «экологический консерватизм» и «филогенетические ограничения».
Впрочем, не будем делать на основе этой работы каких-либо глобальных выводов. На самом деле, эта работа ничего особенного не доказывает, и ничего не опровергает. Хотя между популяциями двух исследованных островков различия оказались минимальными, но ведь в работе указывается пример и другого острова, где эти же анолисы имеют весьма заметные отличия от материковых популяций. Кроме того, и между средой обитания и морфологией организма - далеко не всегда удаётся обнаружить зависимости. Это ведь не математика, а полевая биология. Например, эти зависимости могут иметь слишком низкую силу, чтобы их можно было уловить, исследовав всего 220 ящериц (как в данной работе).
Кстати, еще в одной работе (Calsbeek & Cox, 2009), влияние хищнического давления на морфологию ящериц того же биологического вида (Anolis sagrei) - установить не удалось.
Здесь исследователи сравнили, к чему приведет естественный отбор ящериц через четыре месяца. На изолированных островках для ящериц были созданы разные условия:
1. Полное отсутствие хищников, при низкой плотности особей самих ящериц (на единицу площади).
2. Полное отсутствие хищников при высокой плотности ящериц (на единицу площади).
3. В присутствии хищников. В качестве хищников выступали только хищные птицы.
4. В присутствии хищников. В качестве хищников выступали не только хищные птицы, но и (специально завезенные на островки) змеи. То есть, в последнем случае было самое высокое хищническое давление.
Через четыре месяца авторы исследовали морфологию и поведение всех выживших ящериц, чтобы оценить направление естественного отбора в разных условиях.
Результаты:
1) Хищническое давление соответствующим образом изменило поведение ящериц. Попросту говоря, хищники «загнали» ящериц на более высокие ветки - выжившие ящерицы были зафиксированы на большей высоте (Рис. 10):
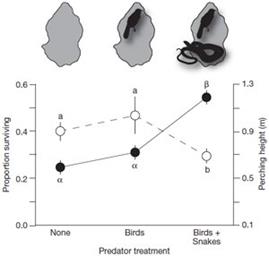
Рисунок 1 0 . Взаимосвязь выживания ящериц и высоты их положения на ветвях в присутствии и отсутствии хищников из работы (Calsbeek & Cox, 2009). «Proportion surviving» - доля выживших ящериц. «Perching height» – средняя высота (ветвей), на которой сидели ящерицы. Рассмотрены три местообитания: «None» - хищников нет, «Birds» - только хищные птицы; «Birds+Snakes» хищные птицы и змеи.
2) Хищническое давление никак не повлияло на морфологию ящериц. Даже в том случае, когда ящерицы подвергались хищничеству со стороны хищных птиц и змей, морфология оставшихся ящериц (размер тела и длина задних ног) не отличалась от морфологии исходных ящериц.
3) Зато в популяциях с высокой плотностью самих ящериц, где, видимо, играла роль конкуренция, морфология выживших ящериц стала отличаться от исходной. А именно, оставшиеся в живых ящерицы оказались (в среднем) более крупными и с более длинными задними ногами, чем среднее значение этих признаков у исходных ящериц (Рис. 11):
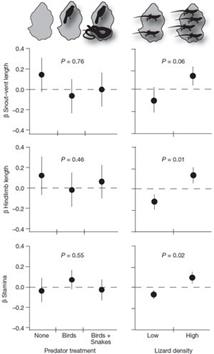
Рисунок 1 1 . Взаимосвязь размера тела и длины задних ног ящериц Anolis sagrei от хищников (слева) и от плотности популяции (справа) из работы (Calsbeek & Cox, 2009). Видно, что хищники не сумели изменить эти морфологические признаки, зато их сумела изменить плотность популяции самих ящериц.
В связи с этими результатами, авторы делают вывод, что внутривидовая конкуренция для ящериц Anolis sagrei является более важным фактором отбора (на морфологию), чем хищническое давление (Calsbeek & Cox, 2009).
И это, собственно, всё, что мы можем сказать по поводу данной работы. Поскольку здесь опять не имеется никакой эволюции. Авторы работы исследовали лишь давление естественного отбора (в разных условиях). О какой-либо «наблюдаемой эволюции» в этой работе - ни слова.
Тем не менее, давайте запомним, на что конкретно нацелено (установленное) давление естественного отбора в случае повышенной внутривидовой конкуренции между ящерицами - оно благоприятствует большим размерам тела, большей длине задних ног и большей выносливости ящериц.
Дата: 2019-02-02, просмотров: 471.