Мы начали обсуждение реабсорбций и секреции с утверждения, что в этих процессах участвуют те же основные механизмы транспорта веществ через плазматическую мембрану, что и в любых иных плазматических мембранах. Однако теперь надо обсудить дополнительную сложность, которая возникает вследствие того, что мы имеем дело с эпителиальным слоем, например почечным канальцем (или желудочно-кишечным эпителием, эпителием желчного пузыря и др.), а не с отдельной неэпителиальной клеткой (например, мышечная клетка или эритроцит).
Посмотрите на рис. 4-1, и вы увидите, что есть два возможных пути для движения реабсорбируемого вещества из просвета в интерстициальное пространство. Первый — движение между клетками, т. е. через плотное соединение двух соседних клеток. Этот путь получил название парацеллюлярный. Парацеллюлярная реабсорбция может осуществляться посредством диффузии или за счет переноса вещества вместе с растворителем.
При диффузии направление электрохимического градиента вещества совпадает с направлением реабсорбций и плотное соединение должно быть проницаемо для данного вещества. При переносе веществ вместе с растворителем поток воды движется через относительно проницаемое плотное соединение (см. ниже). В последующих главах я опишу, как у различных веществ возникают концентрацион ные градиенты при прохождении через различные участки канальца, а также, как обеспечивается поток воды. Здесь я дам лишь краткое описание разности элект рических потенциалов, которая существует между просветом канальца и интер- стициалъной жидкостью в различных сегментах канальца.
Во-первых, важно осознать, что здесь мы имеем дело с трансканальцевой (или трансэпителиальной) разностью потенциалов, а не просто с мембранным потенциалом. Трансканальцевый потенциал — разность потенциалов между просветом канальца и интерстициальной жидкостью — это алгебраическая сумма индивидуальных потенциалов люминальной мембраны и базолатеральной мембраны2. Оба
Механизмы транспорта, участвующие в реабсорбции
85
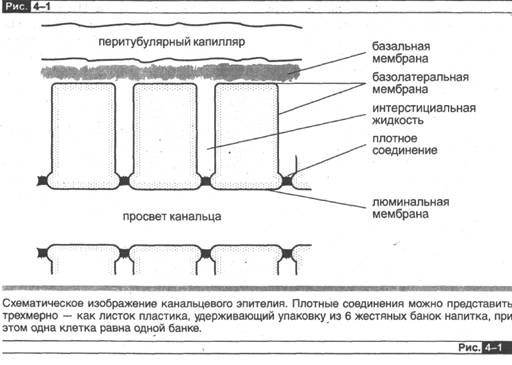
эти мембранных потенциала обеспечивают отрицательно заряженную внутреннюю часть клетки, т. е. внутренняя поверхность клетки является отрицательно заряженной относительно как просвета канальца, так и интерстициальной жидкости. Величина же этих двух отрицательных потенциалов плазматических мембран клетки может отличаться, и это определяет трансканальцевый потенциал как алгебраическую сумму двух упомянутых потенциалов. В проксимальном канальце трансканальцевый потенциал очень мал (объяснение этому будет дано ниже, когда в этой главе пойдет речь о «проницаемом» эпителии), всего несколько мВ, просвет несет отрицательные электрические заряды относительно интерстициальной жидкости в начальных участках канальца, а в средних и конечных участках проксимального канйльца просвет имеет положительное значение потенциала величиной несколько мВ. Поскольку потенциал очень невелик и меняется по длине канальца, мы будем считать трансканальцевую разность потенциалов равной нулю. Это позволит нам сосредоточить основное внимание на концентра ционных градиентах через стенку канальца этого сегмента при рассмотрении вопроса о парацеллюлярной диффузии ионов. В толстой восходящей части петли Генле трансканальцевый потенциал со стороны просвета всегда характеризуется значительной положительной величиной, поэтому он является фактором, влияющим на парацеллюлярную реабсорбцию катионов — натрия, калия, кальция и пр. Трансканальцевый потенциал в большинстве более отдаленных сегментов не-фрона имеет значительную электроотрицательность, и этот фактор приобретает значение для парацеллюлярной реабсорбции анионов, из которых самым важным является хлор.
86
Основные механизмы канальцееой реабсорбции и секреции
Второй путь реабсорбции — трансцеллюлярный («через* клетку), В этом чае реабсорбируемое вещество должно преодолеть две плазматические мембраны на своем пути из просвета канальца к интерстициальной жидкости — люминаль-ную (или апикальную) мембрану, отделяющую жидкость в просвете канальца от цитоплазмы клеток, и базолатеральную (или контрлюминальную) мембрану, отделяющую цитоплазму от интерстициальной жидкости. (Вещество должно, конечно, также пересечь цитозоль клетки между двумя мембранами.)
Вещества с хорошей растворимостью в липидах могут преодолевать обе мембраны (и цитозоль) путем диффузии; при этом происходит чистая пассивная ре-абсорбция одновременно с парацеллюлярным вариантом транспорта вещества, когда для данного вещества имеет место благоприятствующий его движению трансканальцевый электрохимический градиент. Трансцеллюлярный перенос веществ со слабой растворимостью в липидах, напротив, процесс активный. Принципиальным здесь является следующее положение: активная чистая транс-целлюлярная реабсорбция вещества подразумевает, (1) что люминальные и базо-латеральные мембраны асимметричны для данного вещества, т. е. содержат различные каналы и/или переносчики, (2) что для перемещения вещества из просвета канальца в клетку или из клетки в интерстициальную жидкость необходима энергия.
Возьмем в качестве примера реабсорбцию натрия в основных клетках собирательных трубок коркового вещества. Первый вопрос заключается в следующем: имеет ли место значительная по величине реабсорбция натрия в данном сегменте канальца? Правильный ответ — нет. Более того, реабсорбция натрия носит активный, трансцеллюлярный характер, как показано на рис. 4-2. Наблюдается чистая диффузия натрия в соответствии с электрохимическим градиентом (см. ниже) из просвета канальца через люминальную мембрану в цитоплазму по натриевым каналам. Затем натрий активно транспортируется против электрохимического градиента через базолатеральную мембрану в интерстициальную жидкость. Этот последний этап является первично активным процессом с участием Nа,К-АТФаз, обнаруженных только в базолатеральной мембране. Таким образом, чистая однонаправленная реабсорбция натрия в данном сегменте канальца происходит, поскольку (1) имеет место асимметрия люминальной и базолатеральной мембран — натриевые каналы находятся в первой из них и Nа,К-АТФаза (насосы) в последней, и (2) энергия для пересечения базолатеральной мембраны обеспечивается за счет расщепления АТФ с помощью Nа,К-АТФазы.
Данное описание ничего не говорит о том, что процессы в люминальной мембране зависят от того, что происходит в базолатеральной мембране. Это становится очевидным, если мы усвоим, что истинная диффузия натрия через люминальную мембрану зависит от наличия благоприятного электрохимического градиента — цитоплазматическая концентрация [Nа] меньше люминальной [Nа], и/или разница электрических потенциалов ориентирована таким образом, что внутренняя поверхность клетки заряжена отрицательно по отношению к просвету канальца. Принципиальным моментом является то, что насос в базолатеральной мембране (Nа,К-АТФаза) создает этот электрохимический градиент через люминальную мембрану, поддерживая цитоплазматическую концентрацию [Nа] на низком уровне, а внутреннюю поверхность клетки сохраняет заряженной отрицательно, т. е. также, как это осуществляют находящиеся в плазматических мембранах Nа,К-АТФазные насосы.
| 87 |
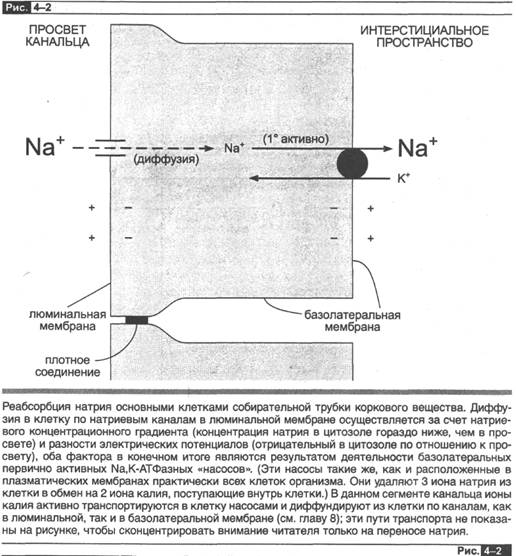
Приведенный пример показывает, как важно отличать трансканальце&ый по тенциал, который является движущей силой парацеллюлярной диффузии через эпителий, от разности потенциалов через отдельные мембраны — люминальные и базолатеральные, которые оказывают влияние на движение ионов через эти мембраны. Трансканальцевый потенциал в собирательной трубке коркового вещества почки характеризуется отрииртелтым значением просвета по отношению к ин-терстициальной жидкости, а потенциал люминальной мембраны является положи тельным со стороны просвета канальца по отношению к внутренней поверхности клетки.
Давайте рассмотрим другой пример — реабсорбцию глюкозы (рис. 4-3) в проксимальном канальце. Глюкоза движется против градиента из просвета канальца
88
Основные механизмы I
I реабсорбции и секреции
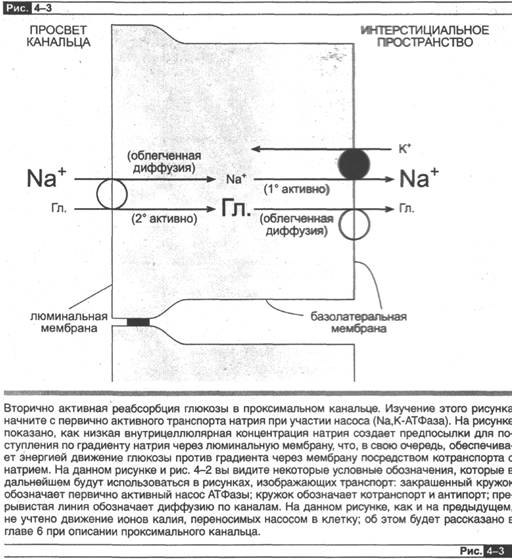
через люминальную мембрану в цитоплазму посредством системы котранспорта с натрием. Это движение глюкозы опосредовано участием переносчика и является вторично активным транспортом, поскольку энергия, необходимая для осуществления движения глюкозы через люминальную мембрану, вырабатывается за счет движения натрия по его электрохимическому градиенту, т. е. посредством котранспорта. Данный механизм котранспорта столь мощный, что позволяет полностью всасывать всю глюкозу из просвета канальца. После проникновений в клетку глюкоза должна преодолеть базолатеральную мембрану, что происходит посредством независимой от участия натрия облегченной диффузии, это движение по градиенту поддерживается за счет высокой концентрации глюкозы, накапливающейся в клетке вследствие активности люминального процесса котран-
Механизмы транспорта, участвующие в рёабсорбции
89
спорта. Итак, чтобы обеспечить активную трансцеллюлярную реабсорбцикю, функционирует система:. (1) с наличием 2 мембран, которые асимметричны по отношению к присутствию переносчиков глюкозы; (2) энергия выделяется только при преодолении одной мембраны, в данном случае люминальной.
Решающий фактор, который легко упустить из виду, состоит в том, что весь процесс рёабсорбции глюкозы зависит в конечном счете от первично активного транспорта натрия Nа,К-АТФазой в базолатеральной мембране. Ранее говорилось, что именно этот насос создает электрохимический градиент, необходимый для транспорта по градиенту натрия через люминальную мембрану, а его движение по градиенту обеспечивает поступление энергии для одновременного движения глюкозы против концентрационного градиента. Теперь, я надеюсь, ясно, почему реабсорбция глюкозы названа вторично активным транспортом: сама по себе она «направлена против градиента» («активная»), но вторична, поскольку зависит от первично активного транспорта натрия через базолатеральную мембрану. Вместо глюкозы мы можем в качестве примера взять аминокислоты, неорганический фосфат, сульфат или целую группу органических питательных веществ, поскольку они подвергаются вторично активной рёабсорбции при котран-спорте с натрием через люминальную мембрану тем же способом (см. главу 5).
Необходимо сделать еще одно замечание, касающееся терминологии. Транс-целлюлярный транспорт определяется термином активный, для краткости, хотя пересечение по меньшей мере одной из двух мембран осуществляется посредством первично или вторично активного процесса, т. е. имеет место перенос вещества против его электрохимического градиента, который происходит между просветом канальца и интерстициальной жидкостью. Все вышесказанное дает нам право говорить, что глюкоза подвергается активной рёабсорбции.
Дата: 2018-09-13, просмотров: 842.