В живых организмах существует целая группа органических фосфатов, гидролиз которых приводит к освобождению большого количества свободной энергии. Такие соединения называют высокоэнергетическими фосфатами. Центральное место среди этих соединений занимает аденозинтрифосфорная кислота (АТФ).

Цикл АТФ-АДФ:

| 5. Продукция тепла |
Цикл АТФ/АДФ связывает процессы, генерирующие высокоэнергетическую фосфатную группу (~P), с процессами, потребляющими ~P.
Имеется 3 главных источника ~P, обеспечивающие улавливание и запасание энергии:
1. Окислительное фосфорилирование (аэробные организмы).
2. Гликолиз.
3. Цикл лимонной кислоты.
Три стадии катаболических превращений основных питательных веществ клетки →
На стадии І сотни белков и многие виды полисахаридов и липидов расщепляются на составляющие их строительные блоки.
На стадии ІІ эти строительные блоки превращаются в один общий продукт – ацетильную группу ацетил КоА.
На стадии ІІІ различные катаболические пути сливаются в один общий путь – цикл лимонной кислоты.
В результате всех этих превращений образуются только три конечных продукта (аммиак, вода, углекислый газ).
Расщепление нуклеиновых кислот происходит также поэтапно, но здесь этот процесс не показан, поскольку его вклад в удовлетворение энергетических нужд клетки сравнительно невелик.
Окисление пирувата до ацетил-КоА происходит при участии ряда ферментов и коферментов, объединенных структурно в мультиферментную систему, получившую название «пируватдегидрогеназный комплекс».
Суммарную реакцию, катализируемую пируватдегидрогеназным комплексом, можно представить следующим образом:
Пируват + НАД+ + HS-KoA→ Ацетил-КоА + НАДН + Н+ + СO2.
Реакция сопровождается значительным уменьшением стандартной свободной энергии и практически необратима.
Образовавшийся в процессе окислительного декарбоксилирования ацетил-КоА подвергается дальнейшему окислению с образованием СО2 и Н2О. Полное окисление ацетил-КоА происходит в цикле трикарбоновых кислот (цикл Кребса). Этот процесс, так же как окислительное декарбоксилирование пирувата, происходит в митохондриях клеток.
Пируват декарбоксилируется в присутствии тиаминдифосфата, при этом происходит перенос гидроксиэтильной группы на тиазольное кольцо, связанного с ферментом тиаминдифосфата (кофермент); далее это гидроксиэтильное производное вступает в реакцию с окисленным липоамидом с образованием ацетиллипоамида.
A – окислительное декарбоксилирование пирувата пируватдегидрогеназным комплексом.
Б – липоевая кислота. Она присоединяется амидной связью к остаткам лизина трансацетилазного компонента комплекса.
В пируватдегидрогеназный комплекс входит примерно 29 молекул пируватдегидрогеназы, около 8 молекул флавопротеина (дигидролипоилдегидрогеназы) и 1 молекула трансацетилазы.

 Всасывание моносахаридов из кишечника происходит путем облегченной диффузии с помощью специальных белков-переносчиков (транспортеров). Кроме того, глюкоза и галактоза транспортируются в энтероцит путем вторично-активного транспорта, зависимого от градиента концентрации ионов натрия. Белки-транспортеры, зависимые от градиента Na+, обеспечивают всасывание глюкозы из просвета кишечника в энтероцит против градиента концентрации. Концентрация Na+, необходимая для этого транспорта, обеспечивается Na+,К+-АТФ-азой, которая работает как насос, откачивая из клетки Na+ в обмен на К+. В отличие от глюкозы, фруктоза транспортируется системой, не зависящей от градиента натрия.
Липопротеины плазмы крови:
Всасывание моносахаридов из кишечника происходит путем облегченной диффузии с помощью специальных белков-переносчиков (транспортеров). Кроме того, глюкоза и галактоза транспортируются в энтероцит путем вторично-активного транспорта, зависимого от градиента концентрации ионов натрия. Белки-транспортеры, зависимые от градиента Na+, обеспечивают всасывание глюкозы из просвета кишечника в энтероцит против градиента концентрации. Концентрация Na+, необходимая для этого транспорта, обеспечивается Na+,К+-АТФ-азой, которая работает как насос, откачивая из клетки Na+ в обмен на К+. В отличие от глюкозы, фруктоза транспортируется системой, не зависящей от градиента натрия.
Липопротеины плазмы крови:
 Типы липопротеинов, их состав, свойства и функции
Типы липопротеинов, их состав, свойства и функции
 Существует 5 специфических транспортных систем, каждая из которых функционирует для переноса определённой группы близких по строению аминокислот:
· Нейтральных, с короткой боковой цепью (Ала, Сер, Тре). Это Na+-зависимые переносчики.
· Нейтральных, с длинной или разветвлённой боковой цепью (Вал, Лей, Иле).
· С катионными радикалами (Лиз, Арг).
· С анионными радикалами (Глу, Асп).
· Иминокислот (Про, оксипролин). Это Na+-зависимые переносчики.
Основываясь на схеме, дайте определение амфиболическим путям –
Пример:
Реакция фосфорилирования глюкозы свободным фосфатом с образованием глюкозо-6-фосфата является эндергонической:
(1) Глюкоза + Р3РО4 → глюкозо-6-фосфат + Н2О ( Δ G =+13,8 кДж/моль).
Для протекания такой реакции в сторону образования глюкозо-6-фосфата необходимо её сопряжение с другой реакцией, величина свободной энергии которой больше, чем требуется для фосфорилирования глюкозы.
(2) АТФ → АДФ + Н3РО4 ( Δ G =-30,5 кДж/моль).
При сопряжении процессов (1) и (2) в реакции, катализируемой гексокиназой, фосфорилирование глюкозы легко протекает в физиологических условиях. Равновесие реакции сильно сдвинуто вправо, и она практически необратима:
(3) Глюкоза + АТФ → Глюкозо-6-фосфат + АДФ ( Δ G =-16,7 кДж/моль).
При переносе электронов по ЦПЭ часть энергии рассеивается в виде теплоты, которая используется теплокровными животными для поддержания температуры тела. При использовании АТФ для совершения работы значительная часть энергии также превращается в теплоту. При снижении температуры тела включается механизм дрожания (несогласованного сокращения отдельных групп мышц). При этом за счет АТФазной активности актомиозина происходит гидролиз АТФ до АДФ и Н3РО4, что стимулирует тканевое дыхание. Полезной работы при этом не происходит, большая часть энергии переходит в теплоту и температура тела повышается.
Существует 5 специфических транспортных систем, каждая из которых функционирует для переноса определённой группы близких по строению аминокислот:
· Нейтральных, с короткой боковой цепью (Ала, Сер, Тре). Это Na+-зависимые переносчики.
· Нейтральных, с длинной или разветвлённой боковой цепью (Вал, Лей, Иле).
· С катионными радикалами (Лиз, Арг).
· С анионными радикалами (Глу, Асп).
· Иминокислот (Про, оксипролин). Это Na+-зависимые переносчики.
Основываясь на схеме, дайте определение амфиболическим путям –
Пример:
Реакция фосфорилирования глюкозы свободным фосфатом с образованием глюкозо-6-фосфата является эндергонической:
(1) Глюкоза + Р3РО4 → глюкозо-6-фосфат + Н2О ( Δ G =+13,8 кДж/моль).
Для протекания такой реакции в сторону образования глюкозо-6-фосфата необходимо её сопряжение с другой реакцией, величина свободной энергии которой больше, чем требуется для фосфорилирования глюкозы.
(2) АТФ → АДФ + Н3РО4 ( Δ G =-30,5 кДж/моль).
При сопряжении процессов (1) и (2) в реакции, катализируемой гексокиназой, фосфорилирование глюкозы легко протекает в физиологических условиях. Равновесие реакции сильно сдвинуто вправо, и она практически необратима:
(3) Глюкоза + АТФ → Глюкозо-6-фосфат + АДФ ( Δ G =-16,7 кДж/моль).
При переносе электронов по ЦПЭ часть энергии рассеивается в виде теплоты, которая используется теплокровными животными для поддержания температуры тела. При использовании АТФ для совершения работы значительная часть энергии также превращается в теплоту. При снижении температуры тела включается механизм дрожания (несогласованного сокращения отдельных групп мышц). При этом за счет АТФазной активности актомиозина происходит гидролиз АТФ до АДФ и Н3РО4, что стимулирует тканевое дыхание. Полезной работы при этом не происходит, большая часть энергии переходит в теплоту и температура тела повышается.


2. Цикл лимонной кислоты (цикл Кребса): последовательность реакций и характеристика ферментов. Реакция субстратного фосфорилирования в цикле лимонной кислоты, макроэргические соединения. Энергетическая и пластическая функции цикла Кребса. Регуляция активности пируватдегидрогеназного комплекса и цикла лимонной кислоты
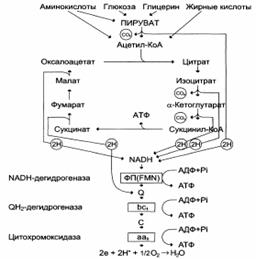 Общая схема цитратного цикла (цикла Кребса, цикл лимонной кислоты, цикл трикарбоновых кислот):
Общая схема цитратного цикла (цикла Кребса, цикл лимонной кислоты, цикл трикарбоновых кислот):
 Цифры 1-6 обозначают реакции цикла.
Использование метаболитов ЦТК в синтезе различных соединений (пластическая функция)
Синтез заменимых аминокислот (1, 2, 3), глюкозы (4, 5, 6), жирных кислот (7), гема (8):
Цифры 1-6 обозначают реакции цикла.
Использование метаболитов ЦТК в синтезе различных соединений (пластическая функция)
Синтез заменимых аминокислот (1, 2, 3), глюкозы (4, 5, 6), жирных кислот (7), гема (8):


|
Зелёным цветом выделена реакция субстратного фосфорилирования
Цикл лимонной кислоты – главный катаболический путь ацстил-СоА у аэробных организмов. Ацетил-СоА – продукт катаболизма углеводов, белков и липидов вступает в цикл вместе с Н2О и окисляется до СО2, поставляя восстановительные эквиваленты (2Н). Последующее окисление 2Н в дыхательной цепи происходит в условиях сопряжения с фосфорилированием ADP. За один оборот цикла 11 связей ~Р образуется путем окислительного фосфорилирования и одна связь ~Р образуется на субстратном уровне при превращении сукцинил-СоА в сукцинат.
 – дыхательная цепь,
ФП – флавопротеин,
Цит – цитохром,
~Р – высокоэнергетический фосфат. – дыхательная цепь,
ФП – флавопротеин,
Цит – цитохром,
~Р – высокоэнергетический фосфат.
|
Дата: 2018-11-18, просмотров: 1367.