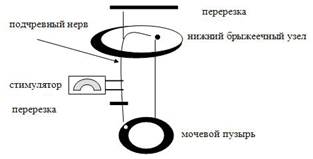
После перерезки симпатических и парасимпатических нервов многие органы способны продолжать присущие им функции без изменений. Например, всасывательная и перистальтическая функции кишки, сократительная способность сердца, изолированные полоски матки, мочеточника, желчного пузыря продолжают сохранять свою сократительную активность.
Во многом функциональная автономия этих органов объясняется наличием в ее стенках ганглиозной системы. Эта система обладает собственным автоматизмом, а также имеет свои местные рефлекторные дуги, которые состоят из чувствительного, вставочного и двигательного звена и медиатора. Становится понятным, что в этих органах управление их работой обеспечивается не только симпатическим и парасимпатическим отделами автономной нервной системы, но и рефлекторными дугами, замыкающимися в пределах стенки самих органов.
Эта местная система имеет ряд черт, которые делают ее похожей на автономную нервную систему:
1) общность структурной и функциональной организации;
2) общность онто- и филогенеза;
3) общность конечных эффектов и т.д.
Это позволило А.Д.Ноздрачеву ввести термин метасимпатической нервной системы.
Однако метасимпатическая нервная система имеет ряд особенностей:
1) иннервирует только полые внутренние органы, обладающие собственной моторной активностью;
2) находится под контролен симпатической и парасимпатической нервной системы;
3) имеет собственные чувствительные клетки;
4) более независима от ЦНС, чем симпатический или парасимпатический отделы автономной нервной системы;
5) устранение метасимпатической регуляции приводит к потере координированной ритмической моторной функции;
6) метасимпатическая нервная система имеет собственное медиаторное звено.
7. Классификация вегетативных рефлексов, примеры истинных автономных рефлексов.
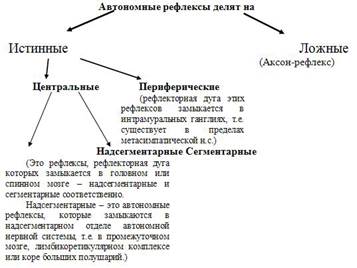
В 1877 году Н.Н.Саковнин описал следующий феномен.
Он показал, что электрическое раздражение центрального конца перерезанного подчревного нерва, производимое в условиях отделения нижнего брыжеечного узла от ЦНС и сохранении интактного другого подчревного нерва приводит к сокращению мочевого пузыря (рис.21).
Он оценил этот феномен как периферический рефлекс, замыкающийся в этом ганглии.
Схема опыта Н.Н.Саковнина
Рис. 21. Схема опыта Н.Н.Саковнина
Однако, Лэнгли, подтвердивший факты Н.Н.Саковнина, объяснил их как ложный рефлекс, псевдорефлекс или аксон-рефлекс. Аксон-рефлекс впервые был описан на электрическом органе нильского сома. Считали, что аксон-рефлексы широко распространены, однако они имеют довольно ограниченное значение. Они могут возникать при распространении возбуждения по разветвлениям аксона. Например, при механическом раздражении кожи происходит ее покраснение. Это обусловлено тем, что одна веточка аксона оканчивается в коже (чувствительная), а другая иннервирует сосуд (сосудодвигательная).
Дальнейшие исследования доказали, что в ганглиях могут замыкаться истинные рефлексы. Этому были получены следующие доказательства. Если перерезать аксоны, то те их части, которые потеряли связь с телом нейрона дегенерируют. Оказалось, что после перерезки подчревного нерва не все волокна, расположенные ниже перерезки, идущие к мочевому пузырю перерождаются. Это доказывает, что тела этих нейронов лежат в стенке мочевого пузыря.
Примеры истинных рефлексов:
1. Рефлекс Данини-Ашнера – надавливание на глазные яблоки приводит к урежению частоты сердечных сокращений.
2. Ортостатический рефлекс – если человек перемещается из горизонтального положения в вертикальное, то у него увеличивается частота сердечных сокращений. Этот рефлекс имеет важное приспособительное значение. Благодаря ему сохраняется постоянный минутный объем кровообращения.
3. Клиностатический рефлекс – это рефлекс противоположный ортостатическому. Когда человек перемещается из вертикального положения в горизонтальное, частота сердечных сокращений уменьшается.
4. Сердечно-сосудистые автономные рефлексы. Помогают контролировать артериальное давление и частоту сердечных сокращений. Одним из них является барорефлекс. Суть его в следующем: растяжение барорецепторов, расположенных в стенке крупных артериальных стволов (включая сонные артерии и дугу аорты), возникающее вследствие повышения артериального давления приводит к передаче сигналов в ствол мозга, где они ингибируют активность симпатического отдела автономной нервной системы. Это приводит к расширению сосудов и уменьшению частоты сердечных сокращений, что возвращает давление к норме.
5. Желудочно-кишечные рефлексы. Верхняя часть желудочно-кишечного тракта и прямая кишка находятся под влиянием автономных рефлексов. Например, запах пищи или попадание пищи в рот инициируют появление сигналов из носа и рта, которые поступают в слюноотделительное ядро продолговатого мозга. Затем по парасимпатическим нервам возбуждение передается к слюнным железам, железам желудка, вызывая выделение слюны и желудочного сока.
Другой пример. При наполнении каловыми массами прямой кишки, она растягивается и от рецепторов растяжения возбуждение передается в крестцовый отдел спинного мозга. Затем по парасимпатическим волокнам импульсы снова возвращаются к толстому кишечнику, и вызывают его сильное перистальтическое сокращение.
6. Другие автономные рефлексы. Опорожнение мочевого пузыря контролируется так же, как и опорожнение прямой кишки, посредством крестцового отдела спинного мозга.
Автономные рефлексы регулируют секрецию поджелудочной железы, опорожнение желчного пузыря, потоотделение, содержание глюкозы в крови и т.д.
Дата: 2019-12-10, просмотров: 440.