В главе 1 мы убедились, что научение и выживание тесно связаны между собой. Классическое обусловливание позволяет организмам различать способствующие и препятствующие выживанию стимулы. Как только эти стимулы начинают узнаваться, инструментальное и оперантное обусловливание позволяют организмам научиться реагировать в соответствии с поступающими из окружающей среды стимулами. Хотя такие ассоцианистские теории, как, например, теория Павлова, имеют непосредственное отношение к выживанию, именно функциональные теории, например Торндайка и Уотсона, отводят эволюционной теории основное место в своих объяснениях научения. Необходимо отметить, что также возможно исследовать отношения между невыученным поведением и выживанием. В период расцвета бихевиоризма группа этологов подчеркивала важность видоспеци-фичного (невыученного) поведения, направленного на выживание. В эту группу входили Карл фон Фриш (1886-1983), Конрад Лоренц (1903-1989) и Нико Тин-берген (1907-1988), которые получили Нобелевскую премию по биологии в 1973 г. Обычно этологи изучали специфическую категорию поведения (например, агрессию, миграцию, коммуникацию, территориальное поведение) в естественной для животного среде и пытались объяснить такое поведение в терминах теории эволюции. Методы, которых придерживались этологи, отражены в трудах некоторых современных психологов. Уильям Тимберлейк и его коллеги (Timberlake, 1997, 1999; Timberlake & Lucas, 1989; Timberlake & Silva, 1995) были, например, сторонниками «биологического бихевиоризма, сконцентрированного на животных», подхода, который объединяет биологическое, эволюционное и физиологическое понимание специфических категорий поведения, имеющего место в естественной среде. Этологи впервые заговорили о том, что для полного понимания поведения необходимо принять во внимание и выученные и невыученные стремления. Осознание этого проложило путь к значительным изменениям в бихевиористской теории, которые мы обсудим в этой главе.
В последнее время детально исследовался вклад эволюционной теории в понимание процесса научения. Например, было обнаружено, что некоторые виды животных с легкостью научаются тому, что особям другого вида дается с трудом или не дается вообще. Объяснение этих видоспецифичных различий является одной из задач эволюционной психологии, которая детально исследует вклад теории Дарвина и неодарвинизма в объяснение поведения организмов. В этой главе мы рассмотрим дальнейшие разработки теории эволюции в области понимания процесса научения.
После краткого обзора теории эволюции мы остановимся на научной деятельности Боллса (1928-1994), пытавшегося, как и многие другие ученые, объяснить процесс научения согласно эволюционным принципам. В заключении мы кратко осветим взгляд эволюционной психологии на научение человека.
Теория Дарвина и эволюционная психология. Естественный отбор и адаптация
Хотя ранние биологи и натуралисты временами обращались к различиям видов и биологических структур, именно работа Дарвина «Происхождение видов в результате естественного отбора» (On the original of species by means of natural selection, Darwin, 1859/1958) предложила причину этих различий — естественный отбор. Основные принципы естественного отбора и их значение для эволюционной психологии изложены ниже.
Во-первых, существует естественное разнообразие внутри видов. Это разнообразие может выражаться в большей остроте зрения, физической силе или скорости научения некоторых особей внутри одного вида. Эти индивидуальные различия служат базовыми составляющими, необходимыми для процесса эволюции (Buss, Haselton, Shackelford, Bleske & Wakefield, 1998; Crawford, 1998).
Во-вторых, лишь некоторые индивидуальные особенности передаются по наследству. То есть только некоторые черты дети могут наследовать от своих родителей и передать последующим поколениям. Как правило, изменения вследствие генетических мутаций или влияний среды невыгодны членам рода и не передадутся потомству. Аналогично приобретенные паттерны поведения вне зависимости от их адаптивной функции могут передаваться последующим поколениям через научение, но не наследуются. Теория эволюции скорее имеет дело с передаваемой по наследству вариативностью поведения, чем с отклонениями поведения, возникшими в результате других причин.
Наконец, естественный отбор может существовать благодаря взаимодействию между свойствами организма и требованиями среды его обитания. По словам Басса и коллег (Buss, Haselton, Shackelford, Bleske & Wakefield, 1998):
Организмы с определенными наследуемыми свойствами дают больше потомства в среднем, чем особи, у которых эти черты отсутствуют, потому что эти особенности помогают решить специфические проблемы и способствуют репродукции в данной среде. Различная репродуктивная успешность вследствие обладания наследственными особенностями служит двигателем эволюции путем естественного отбора.
Адаптацией называется физиологическая или анатомическая структура, биологический процесс или паттерн поведения, которые исторически способствовали выживанию и размножению вида (Wilson, 1975). По определению адаптация происходит путем естественного отбора и должна передаваться по наследству (Buss, Haselton, Shackelford, Bleske & Wakefield, 1998; Tooby & Cosmides, 1992). Так, определенное генетическое отличие — например, цветовое зрение — может привести к лучшей выживаемости и, что еще важнее, к более высокой репродуктивное™ среди особей, обладающих данными адаптационными способностями. В результате эта адаптационная особенность распространяется на последующие поколения, даже если в будущем она уже не принимает непосредственного участия в выживании и воспроизводстве.
Заблуждения относительно адаптации. Кроуфорд (Crawford, 1998) предостерегает от ошибочного понимания принципа «выживания сильнейшего». Бытует мнение, что при естественном отборе предпочтение отдается наиболее сильным и агрессивным членам рода и за эволюционными успехами кроются жестокие битвы, в которых побеждают только доминантные особи. Однако в некоторых видах успешными могут быть особи, чьим приспособлением служит способность прятаться или избегать столкновений, носящих угрожающий для жизни характер. Иными словами, эволюционная приспособляемость, определяемая репродуктивной успешностью, часто не зависит от физической силы особи, с которой мы обычно связываем это понятие.
Другие ученые (Buss, Haselton, Shackelford, Bleske & Wakefield, 1998, p. 538) также призывали нас избегать заблуждений о том, что естественный отбор приводит к оптимальной адаптации в данных условиях. «Селекция не похожа на инженера, последовательно от начала до конца выполняющего проект. Отбор работает только с имеющимся в наличии материалом и не предвидит результат». Таким образом, в ходе медленного процесса эволюции, использующего исключительно генетический материал организма и скованного другими биологическими характеристиками организма, вырабатываются приспособления, применимые к проблемам специфической среды, которая может измениться в будущем. «Приспособления не являются оптимально сконструированными механизмами. Они скорее описываются как оснащенные улучшенные решения... сдерживаемые по качеству и конструкции различными историческими и современными силами». Также важно разрушить общепринятое мнение, что у эволюции есть конечная цель, к которой она движется. Например, многие верят, что эволюция разворачивается в соответствии с заданным планом. Это не так. Эволюция не обязательно несет прогресс. Естественный отбор означает, что организмы, обладающие адаптивными для данной среды чертами, скорее выживут и размножатся. Как сказано выше, эволюция не обладает «предвидением».
Наконец, Басе с коллегами (Buss, Haselton, Shackelford, Bleske & Wakefield, 1998) и Гоулд (Gould, 1991) требовали не злоупотреблять адаптационными объяснениями. Использование биологической структуры с определенной целью не означает, что данная структура развивалась с этой целью. В качестве примера Гоулд обращает внимание на то отношение, какое оперение птиц имеет к полету. По его мнению, оперение эволюционировало как механизм терморегуляции тела птицы и позже было приспособлено для полета. Гоулд называет кооптирование органов (в данном случае оперения) к полезному, но не соответствующему назначению (полету в сравнении с терморегуляцией) коадаптацией.
Наравне с коадаптацией кооптированными могут быть и побочные результаты, так называемые надстройки, которые могут сопровождать определенную адаптацию. Например, развитие способностей человеческого мозга может предоставить множество адаптивных возможностей. Эти возможности могут включать в себя усовершенствованные навыки решения проблем, навыки изготовления инструментов более высокого качества, лучшую память в отношении местонахождения пищи или мест обитания опасных хищников и т. д. К побочным результатам развития способностей человеческого мозга можно, видимо, отнести способности к созданию музыки, письменности и сложных социальных норм, которые можно ошибочно все без исключения считать приспособлениями, ведущими к выживанию и улучшению репродуктивной способности.
Совокупная приспособленность и неодарвинизм
Как мы видели, Дарвин измерял приспособляемость количеством воспроизведенного организмом потомства. В 1964 г. Уильям Гамильтон (1936-2000) расширил границы определения Дарвина, предложив идею совокупной приспособленности. В совокупной приспособленности фокус смещается от успешности воспроизводства конкретной особи вида к закреплению индивидуальных и общевидовых генов. Так мы подходим к рассмотрению родительского поведения или сотрудничества внутри семейной группы как адаптивного, поскольку они способствуют выживанию и, возможно, репродуктивной успешности скорее через общие, чем индивидуальные гены. С точки зрения совокупной приспособленности поведение, которое может быть опасным для отдельных особей, рассматривается как адаптивное, так как жертва индивида может обеспечить выживание другим членам вида, которые имеют общие с ним гены.
Возникшая в рамках неодарвинизма концепция совокупной приспособленности оказалась чрезвычайно эвристической. Помимо трактовки «альтруистического» поведения, ее использовали для объяснения совершенно иных тем, например суицида и гомосексуальности (см. Hergenhahn & Olson, 1999).
Теория научения Боллса
Роберт Боллс родился в 1928 г. в городе Сакраменто, штат Калифорния, и находился на домашнем обучении до 12 лет. Он получил степень бакалавра гуманитарных наук в Стэнфордском университете в 1948 г., а через год там же — степень магистра в области математики. Он был принят на работу в Американскую морскую радиологическую оборонную лабораторию, расположенную около Сан-Франциско, штат Калифорния, где он встретил своего будущего коллегу и друга на всю жизнь Джона Гарсиа (вспомните знаменитый эффект Гарсиа), находившегося здесь по докторской программе в области психологии от Калифорнийского университета в Беркли (Garcia, 1997). Вскоре Боллс присоединился к Гарсиа и тоже стал работать по этой программе в Беркли, где они оба обучались у Толмена. Именно тогда, во время учебы в аспирантуре, Боллс и Петрайнович провели ранние эксперименты, направившие интересы Боллса на теорию эволюционного научения (Bolles & Petrinovitch, 1954; Petrinovitch & Bolles, 1954). После защиты докторской диссертации и получения степени доктора философии в 1956 г. Боллс некоторое время работал на факультете Университета Пенсильвании, а затем в Принстонском университете. В 1959 г. он перешел в университетский колледж Холлинз, а в 1964 г. — в Университет Вашингтона, где и проработал всю оставшуюся жизнь, до дня своей кончины от сердечного приступа 8 апреля 1994 г.
В ходе своей научной карьеры Боллс опубликовал более 160 научных статей и выпустил три важных учебника, включая книгу о теории научения. В 1981-1984 гг. он был редактором журнала «Поведение и научение животных» ( Animal Learning and Behavior ), и многие его студенты с успехом продолжили исследование этой темы и внесли важный научный вклад, отнеся эволюционные процессы к научению (см. Bouton & Fanselow, 1997).

Основные теоретические положения
Ожидания. Согласно Боллсу, научение включает в себя выработку ожиданий. То есть организм научается тому, что какое-либо одно событие, как правило, предшествует другому. Мы уже знаем из главы 7, что Боллс объясняет классическое обусловливание как выученное ожидание того, что за одним предъявленным стимулом (условным стимулом) последует другой (безусловный стимул). В повседневной жизни иллюстрацией этого типа ожидания стимул-стимул (С-С) может быть молния, за которой должен последовать гром. Если классическое обусловливание включает в себя развитие ожиданий С-С, то оперантное и инструментальное обусловливание включает в себя развитие ожиданий стимул-реакция (С-Р) (Bolles, 1972). Например, крыса научается ожидать, что после нажатия на рычаг в проблемном ящике Скиннера она получит подкрепление. В повседневной жизни ожидание услышать звук после нажатия на кнопку звонка является примером ожидания по типу Р-С. При обсуждении Р-С ожиданий можно представлять С как последствие реакции. Научение ожиданию в теории Боллса не требует подкрепления. Вообще очередность и близость двух стимулов или реакции и ее последствий определяет природу выученных ожиданий: вспышка молнии становится предвестником грома, и нажатие на кнопку предвещает мелодию звонка, а не наоборот (Staddon, 1988). Следовательно, Боллса можно отнести к теоретикам, отводящим особое место «направленной» смежности стимулов.
Врожденная предрасположенность. Акцентирование Боллсом ожидания свидетельствует о влиянии на него Толмена (см. главу 12). Так или иначе между этими теориями существовали различия. В то время как Толмен главным образом фокусировался на выученных С-С и Р-С ожиданиях, Боллс подчеркивал важность врожденных С-С и Р-С ожиданий в анализе поведения, и именно выделение им врожденных С-С и Р-С ожиданий поставило его в один ряд с другими психологами, которые интересовались объяснениями поведения с точки зрения теории эволюции. Примером врожденной связи С-С служит проявление страха при громком звуке у младенца, на основе чего можно предположить, что младенец ожидает появления опасности вслед за звуком. Врожденные Р-С ожидания проявляются в стереотипном поведении, которое начинают демонстрировать разные виды животных в присутствии еды, воды, опасности и других биологически важных объектов или событий.
Согласно Домжану (Domjan, 1997, р. 32), недостатком таких традиционных эмпирических теорий научения, как, например, Торндайка, Уотсона, Скиннера и Халла, является утверждение, известное под названием эмпирический принцип эквипотенциальности (не путать с законом эквипотенциальности Карла Лэшли). Эмпирический принцип эквипотенциальности гласит, что законы научения «относятся в равной степени ко всем типам стимулов и реакций». Таким образом, эмпирический принцип эквипотенциальности подтолкнул ученых к исследованию научения конкретных видов животных без учета эволюционной истории этих видов. Более того, было сделано неверное предположение, что феномен научения, наблюдаемый у одного вида, например крыс, можно перенести на большинство, если не на все другие виды. А когда члены вида не научались выполнять реакцию при заданных условиях, неудовлетворительный результат относили к поломкам оборудования, ошибке экспериментатора или необъяснимым «помехам».
В опровержение положения об эквипотенциальности Боллс говорил (Bolles, 1988):
Я убежден, что невозможно многого достичь, делая предположение о наличии некоей структуры в событиях, о которых узнает животное, и о существовании соответствующей структуры в научающемся организме... Организм может достичь успеха, если будет способен научаться тому, что необходимо выучить. Здесь работает генетически запрограммированная способность к научению нативиста, а не проявляющаяся время от времени способность к научению эмпирика.
Ниже мы увидим, как эволюционная психология, подчеркивающая важность скорее врожденных, чем приобретенных С-С и Р-С ожиданий, помогает прояснить многие аномалии, обнаруженные в период ранних исследований в области научения. Мы уже рассмотрели один такой пример в главе 5 при обсуждении «аномального поведения организмов».
Мотивация ограничивает гибкость реакции. Некоторые ученые, теории которых мы рассмотрели, минимизировали или отрицали роль мотивации в процессе научения (например, Газри и Толмен). Другие исследователи (например, Халл) придавали большое значение мотивационному состоянию организма. Очевидно, что Боллс разделял точку зрения последних. Для него мотивация и научение были неразделимы. Однако в подходе Боллса необходимо знать как текущие потребности организма, так и действия организма в естественных условиях при наличии данных потребностей. Согласно Боллсу (Bolles, 1979,1988), хотя организм может быть достаточно гибким в отношении С-С ожиданий, которым он научается, Р-С ожидания более ограничены, поскольку мотивация создает искажения реакции. То есть животному будет очень трудно научиться поведению, которое противоречит естественному поведению в данной ситуации. Например, оно не выучит реакцию, относящуюся к избеганию, в целях получения доступа к пище, равно как и реакцию пробуждения аппетита для избежания опасных или болезненных стимулов.
Дискуссия по поводу ниши. Боллс (Bolles, 1988) настаивал на том, что понимание научения неотделимо от понимания истории эволюции организма. Он утверждал, что животные обязаны научиться одному и не научиться другому в зависимости от ниши, которую они занимают в окружающем мире. Нам следует ожидать, что какой-то опыт отразится в научении, а какой-то нет... Задача на выработку навыка, нарушающая априорную биологическую принадлежность животного к определенной нише, скорее всего спровоцирует аномальное поведение. Задача на выработку навыка, учитывающая предрасположенность животного к определенным типам поведения, с большой вероятностью будет успешно выполнена. Это и есть дискуссия по поводу ниши.
Другие эволюционные психологи расширили тему дискуссии по поводу ниши, внеся идею о среде эволюционной адаптации (СЭА), понятии, которое относится к среде как социальной, так и физической, в которой возникла специфическая адаптация (Bowlby, 1969; Tooby & Cosmides, 1990). Эти и другие авторы (например, Sherman & Reeve, 1997) подчеркивали, что СЭА — это не просто доисторический временной период или место, существовавшее в эпоху развития видов. Это скорее комбинация социальных и средовых факторов, действующих в определенный временной период, и остается открытым вопрос, могут ли различные адаптации видов происходить при разных СЭА. Более того, возвращаясь к идее о том, что эволюция не гарантирует прогресс, они отмечают, что, возможно, сейчас организмы испытывают воздействие отбора, отличное от существовавшего в определенной СЭА. Существуют примеры, когда поведение, сформированное эволюционными влияниями, недостаточно приспособлено для современной среды.
Биологические границы научения
В этой части мы обобщим факты, которые продолжают свидетельствовать против утверждения о том, что формирование условных рефлексов происходит автоматически, если непроизвольно возникшая реакция получает подкрепление или какой-либо условный стимул сопровождается безусловным. Как мы уже видели, все большее признание получает тот факт, что в любом эксперименте, касающемся научения, необходимо принимать во внимание природные способности организма. Концепция Брилендов об инстинктивном дрейфе (глава 5) продемонстрировала значимость тенденции к инстинктивному реагированию в ситуации оперантного обусловливания. Как мы убедились, теория Боллса основывается на идее о том, что врожденные склонности ограничивают количество связей, которым организм может научиться, и реакции организма на специфические ситуации. Это положение поддерживает Селигман (Seligman, 1970), настаивающий на том, что одни виды осваивают связи с большей легкостью, чем другие, поскольку они к этому биологически подготовлены. Аналогично усвоение связей для некоторых видов будет сложной задачей, так как они биологически не готовы к подобному обучению. Таким образом, легкость научения определяется тем, какое место в континууме готовности занимает связь.
Инструментальное обусловливание
Затруднения с эмпирическим принципом эквипотенциальности возникали в некоторых ранних исследованиях научения. Торндайк (Torndike, 1898), к примеру, рассказывал, что кошки могут научиться ряду реакций, используя лапы для добывания пищи, но они не научатся вылизываться для получения подкрепления в виде еды. Очевидно, это был тот самый случай, когда естественная реакция кошки на голод не включала в себя поведение, связанное с вылизыванием. Или, как объяснил бы Селигман, кошка не была биологически готова ассоциировать чистку шерстки с едой. Боллс (Bolles, 1988 р. 5) так прокомментировал данные Торндайка: «Никто не обращал никакого внимания». В начале 1950-х, окончив университет в Беркли, Боллс и Петрайнович провели исследование, вызвавшее новую волну интереса к влиянию эволюции на научение.
В предварительном эксперименте Петрайнович и Боллс (Bolles & Petrinovitch, 1954) обучали одну группу крыс поворачивать налево, а вторую направо в Т-образном лабиринте. Т-образный лабиринт назван так, потому что по своей форме он напоминает заглавную букву Т. Крыса начинает движение из «подножия» Т и бежит к точке выбора на пересечении горизонтальной и вертикальной секций, где она может повернуть направо или налево. Половине крыс в обеих группах не давали воды, и когда они выполняли поворот в нужную сторону, в качестве подкрепления они получали воду; оставшихся крыс не кормили и давали подкрепление
в виде пищи за правильный поворот. В этом исследовании крысы, которые утоляли жажду, получая воду как подкрепление, осваивали задание быстрее и делали меньше ошибок, чем голодные крысы, работавшие за пищу. Было интересно: почему тип подкрепления (еда либо вода) воздействовал на эффективность научения?
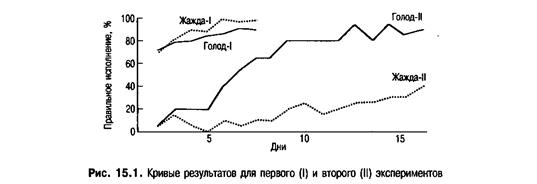
Во втором эксперименте крыс снова лишали воды или пищи. При первой попытке они получали воду или еду в качестве соответствующего подкрепления независимо от направления поворота, которое они выбрали, например направо. Во второй раз они подкреплялись только в случае выполнения поворота в другую сторону — в нашем примере налево. В третий раз они получали подкрепление за выбор, противоположный сделанному во второй попытке, и так на протяжении всего эксперимента. Таким образом, крысы получали подкрепление за чередование право- и левосторонней реакции. В этом исследовании голодные крысы, работавшие за пищу, научались выполнять задание быстрее, нежели те, которые хотели получить воду. Результаты обоих экспериментов изображены на рис. 15.1.
Почему подкрепление водой способствовало лучшему выполнению задания в первом эксперименте и не давало того же результата во втором? Такие ученые, как Халл и Скиннер, затруднились бы объяснить полученные результаты. Однако эволюционное объяснение разрешает проблему. Петрайнович и Боллс полагали, что поскольку крысы эволюционировали как всеядные, у них выработалось предубеждение против поиска пищи в том же месте, где попытка была успешной. Для всеядных пища — нестабильный ресурс, который вряд ли можно отыскать сегодня там же, где он был обнаружен вчера. С другой стороны, для крыс будет логично искать воду на том же месте, что и раньше. Вода — более стабильный ресурс; маловероятно, что река или пруд исчезнут за ночь. Итак, крысы в эксперименте продемонстрировали тенденции реагирования, сложившиеся в процессе эволюции. Иначе говоря, крысы привыкли идти в одном направлении за водой, но не были подготовлены искать еду в том же месте более чем один раз.
Бегство и избегание. Особи могут демонстрировать определенную гибкость реакций и исследовательские способности, добывая воду или пищу. Например, голодные крысы могут нажать на рычаг, пройти лабиринт, заглянуть в маленькую чашечку и пр., чтобы получить шарик корма. Однако Боллс (Bolles, 1970) обращает внимание на случаи, когда животное не получает вознаграждения при научении методом проб и ошибок. Бегство от хищника должно быть успешным с первой попытки, когда речь идет о выживании организма. Поэтому Боллс утверждал, что врожденные Р-С ожидания существуют для решения проблем во внешней среде, несущих угрозу выживанию. Боллс (Bolles, 1988) писал:
Стратегия крысы заключается в использовании фиксированных паттернов поведения для самозащиты, которые я называю видоспецифичными защитными реакциями (Bolles, 1970). В реакции самой по себе очень мало гибкости; животное главным образом научается распознавать стимулы; оно узнает, какие стимулы опасны и какие безопасны.
Боллс (Bolles, 1970, 1972) отмечал, что в видоспецифичные защитные реакции (ВСЗР) крысы входят застывание на месте, бегство, визг, подпрыгивание и агрессия по отношению к объекту. В естественной среде одна или несколько таких реакций приводят к спасению, и в этом случае проблема решена. Также в лаборатории, если одна из этих врожденных реакций на боль позволит животному избежать боли, реакция избегания будет быстро усвоена. При только что описанных обстоятельствах животное в действительности не научается никаким новым реакциям; негативная стимуляция просто вызывает ВСЗР.
Обусловливание избегания немного сложнее. Согласно Боллсу, обусловливание избегания предполагает как С-С, так и Р-С ожидания. Поскольку при обусловливании избегания сигнал предшествует неприятному событию, например удару током, животное научается ожидать появления боли, скажем, после звукового сигнала. Так как звук сигнализирует об опасности, он вызывает те же ВСЗР, которые вызывают болезненные стимулы сами по себе. Поэтому, по Боллсу, либо предвестники боли, либо сама боль вызывают ВСЗР.
Очевидный прогноз, вытекающий из анализа Боллса, состоит в том, что чем ближе реакция, требующаяся от животного в эксперименте, к его естественным действиям в данной ситуации, тем вероятнее научение данной реакции. Если необходимая реакция не является частью врожденного поведенческого репертуара животного, она будет усвоена с большим трудом, а возможно, и вовсе не закрепится. Примером служит тот факт, что голубей можно легко научить перелетать с одной ветки на другую для избегания удара током, но практически невозможно обучить их клевать кнопку для избегания удара (Bolles, 1979, р. 185). Таким образом, выбор реакции для научения животного в условиях эксперимента является главным фактором, определяющим результаты эксперимента.
Даже случайное наблюдение за крысами в различных ситуациях научения избеганию обнаруживает огромную значимость типа реакции, который требуется от животного; это может определить, состоится ли научение с первой попытки или не произойдет и за тысячу проб. Это гораздо важнее, чем вероятность подкрепления, различные параметры эксперимента, предыдущий опыт животного или различные виды физиологических вмешательств. Выбор необходимой реакции, очевидно, не является делом случая или просто вопросом убеждений, это главный фактор, от которого зависит скорость научения. Серьезным обвинением в адрес классических теорий научения является то, что они не делали и намека на важность выбора реакции.
Оперантное обусловливание
В главе 5, где упоминалась деятельность Брилендов, мы говорили, что оперантное научение, подобно вышеописанным типам научения, ограничено природными тенденциями реагирования, присущими организму. Случаи оперантных неудач из-за несоответствия требований задачи природной предрасположенности (или биологической готовности) к определенному набору реакций имеются в достаточном количестве. С другой стороны, довольно часто встречаются эксперименты, в которых голуби клюют подсвечивающиеся кнопки для получения подкрепления в виде пищи. Как и голуби, зяблики научаются клевать кнопку для получения корма; и хотя они способны выучить некоторые оперантные реакции для того, чтобы услышать запись с пением зяблика, они не научатся клевать кнопку с этой целью (Stevenson-Hinde, 1973). Клевание является естественной реакцией на наличие пищи, но зяблики биологически не предрасположены ассоциировать клевание с пением птиц. Было экспериметально доказано (Bolles, Riley, Cantor & Duncan, 1974), что крысы научаются ожидать появления пищи, если она предъявляется в одном и том же режиме подкрепления с фиксированными промежутками (ФП) (раз в день), но они не способны научиться предвидеть болезненные удары электрошока, если они подаются в том же ФП режиме. Боллс писал (Bolles, 1988):
Крысы без труда могут научиться бежать взад-вперед, чтобы избежать удара током. Но им необычайно трудно нажать на рычаг для предотвращения электрического удара. Также не раз отмечалось, что было легко обучить крысу нажимать на рычаг для получения корма, но совсем другое дело было обучить ее нажимать на рычаг, чтобы предотвратить негативные последствия. В литературе встречалось огромное количество таких аномалий. Никогда не сообщалось, что голуби, столь искусные в проявлении тонкой сенсорной проницательности при клевании кнопки для получения пищи, так же умело решали другую биологическую проблему (избежание негативных последствий).
Классическое обусловливание
Обусловленная вкусовая аверсия. В главе 7 мы вкратце познакомили вас с эффектом Гарсиа. В этом разделе мы предоставим более полное описание эксперимента, поставленного Гарсией и Келлингом (Garcia & Koelling, 1966), и рассмотрим важный вклад эффекта Гарсиа в понимание влияния эволюции на научение. Гарсиа и Келлинг дали возможность крысам, которые хотели пить, получить воду в четырех ситуациях. Первой группе предложили светящуюся и шумящую воду, питье которой сразу же сопровождалось ударом током по лапам. Светящаяся и шумящая вода была создана путем прикрепления электрода к питьевой трубе таким образом, что прикосновение организма к трубе вызывало вспышки света и громкие трещащие звуки. Второй группе дали светящуюся и шумящую воду, но вместо получения электрошока при питье их облучали рентгеном, чтобы вызвать тошноту. Третья группа получала воду без вспышек света и треска, но со вкусом сахарина; эти животные, как и в первой группе, подвергались электрошоку сразу после питья раствора сахарина. Четвертой группе дали раствор сахарина и просветили рентгеновскими лучами, после чего животные плохо себя чувствовали.
Гарсиа и Келлинг (Garcia & Koelling, 1966) обнаружили, что у животных из первой группы развилась аверсия к светящейся и шумящей воде, а с животными из второй группы этого не произошло. К тому же у третьей группы животных не сформировалась аверсия к воде со вкусом сахарина, тогда как у четвертой она появилась. План эксперимента и результаты отражены в данной схеме.
Группа 1. Светящаяся и шумящая вода -> электрошок. Появилось отвращение к воде.
Группа 2. Светящаяся и шумящая вода -> тошнота. Нет отвращения к воде.
Группа 3. Раствор сахарина -> электрошок. Нет отвращения к раствору.
Группа 4. Раствор сахарина -> тошнота. Появилось отвращение к раствору.
Видно, что светящаяся и шумящая вода становится действенным УС в паре с электрошоком, но совместно с тошнотой. Так же дело обстоит со вкусом сахарина, являющимся эффективным УС вместе с тошнотой, а не с электрическим разрядом. Гарсиа и Келлинг (Garcia & Koelling, 1966) объяснили полученные результаты наличием естественных взаимосвязей между внешними событиями и болью, которую испытывали животные. Иными словами, боль приходила «извне», и поэтому животные искали внешний предвестник боли, которым в данном случае были свет и шум, ассоциирующиеся с питьем. Тошнота же переживалась скорее внутренне, чем внешне. Поэтому животные связывали вкус сахарина (внутренний признак), а не светящуюся шумящую воду (внешний признак) с тошнотой. Используя терминологию Селигмана, можно сказать, что крысы были биологически готовы создать ассоциацию между светящейся шумящей водой и болью, но не способны к образованию связи между светящейся шумящей водой и тошнотой. Аналогично животные были биологически подготовлены сформировать ассоциацию между вкусом сахарина и тошнотой и не были биологически готовы связать вкус сахарина с болью.
Другое исследование (Wilcoxon, Dragoin & Krai, 1971) служит примером концепции готовности Селигмана, так как относится к различиям между видами. В этом исследовании крысам и перепелкам давалась соленая вода голубого цвета, которая вызывала у них болезненное состояние. После того как оба вида пили воду и заболевали, им предлагался выбор между голубой и соленой водой. Крысы избегали пить соленую воду, а перепелки — голубую. Эти данные отражают тот факт, что крысы в еде (или питье) полагаются на вкус, а перепелки — на визуальные признаки. Таким образом, каждый вид образовал взаимосвязь в соответствии со своими генетическими особенностями. Другими словами, хотя БС (голубая соленая вода) и БР (болезнь) были одинаковы для обоих видов, каждый вид выбрал УС в соответствии со своей генетической информацией. Для крыс вкус соли стал УС, в то время как для перепелки УС послужил голубой цвет. На языке Селигмана, крысы были более биологически подготовлены к созданию связи «соль — болезнь», а перепелки — взаимосвязи «голубой цвет — болезнь».
Исследование Гарсиа показывает, что внутри видов определенные связи будут легче сформированы, чем другие, из-за истории эволюции этих видов. Эксперимент Вилкоксона с соавторами (Wilcoxon, Dragoin & Krai, 1971) демонстрирует различия между видами: разные ассоциации будут оптимальными для разных видов. Однако, как отмечали другие исследователи (Logue, 1988; Rosin & Fallon, 1981), последним стимулом, который испытывает организм, прежде чем проглотить токсичное вещество, является вкус этого вещества. Поэтому данные авторы утверждают, что о хорошей приспособляемости свидетельствовала бы скорее способность организма приобретать аверсию к вкусу, чем к форме, цвету или фактуре еды или напитка, приводящих к заболеваниям. Конечно, несмотря на вышеописанные типы видовых различий, большинство позвоночных могут научиться испытывать аверсию только к оттенкам вкуса самим по себе (Gustavson, 1977).
Как и другие условные реакции, выученная вкусовая аверсия подвержена угасанию. Иными словами, если запах (УС) периодически присутствует без по следующего заболевания (БР), животные вновь начнут употреблять в пищу отвергнутое однажды вещество. Кэтлин Чамберз, одна из выпускниц Боллса в начале 1970-х, обнаружила: вкусовые аверсии быстрее угасают у крыс-самок, чем у крыс-самцов (Chambers, 1985;Chambers & Sengstake, 1976); и впоследствии она интенсивно исследовала половые различия (см. Chambers, Yuan, Brownson & Wang, 1977). Ее интерпретация различий в скорости угасания, с которой Боллс был согласен, очевидно, из ряда эволюционных. Чамберз считала, что поскольку самки ответственны за выживание плода и новорожденного, заботу о потомстве, для них чрезвычайно важно удовлетворять потребности в питании. Поэтому для них адаптивной является способность четко определить, была ли на самом деле болезнь связана с определенной пищей. То есть они биологически подготовлены «перепроверить» потенциально питательную еду с учетом того, что заболевание могло быть вызвано другими факторами.

Однако следует отметить, что для угасания вкусовой аверсии организму необходимо вновь попробовать еду на вкус (УС), не испытывая болезненных ощущений (БР). Принимая во внимание природу эффекта Гарсиа, угасание может произойти в лабораторных условиях, но в естественной среде организм будет продолжать избегать УС, делая угасание аверсии невозможным.
Автоформирование
В главе 5 мы говорили о загадке автоформирования. Наверное, вы помните, что в эксперименте Брауна и Дженкинса (Brown & Jenkins, 1968) диск для клевания ненадолго подсвечивался перед не зависящим от поведения предъявлением корма. Независимо от поведения голубей, продемонстрированного в этом эксперименте в интервале перед предъявлением пищи, фактически все голуби начинали клевать подсвеченный круг. Не потребовалось никакого систематического формирования реакции клевания диска. Как было отмечено в главе 5, Боллс (Bolles, 1979) предлагал объяснение, что автоформирование включает С-С научение, но никакого научения новой реакции не происходит. Скорее он интерпретировал клевания как врожденную реакцию на стимул, который благодаря временной ассоциации с предъявлением пищи вызывает свойственное приему пищи поведение.
Мы стали думать о реакции клевания кнопки как об оперантной, произвольной, удобной, легкоизмеряемой реакции, интенсивность которой можно контролировать за счет ее последствий. Но мы можем видеть в процедуре автоматического формирования условных рефлексов... что сила ответной реакции не зависит от ее последствий. Очевидно, она контролируется свойствами подсвеченной кнопки, сигнализирующими о появлении пищи.
Стаддон (Staddon, 1988), который провел исследование под руководством Боллса, подчеркивает прогнозируемую ценность подсвеченной кнопки для клевания и соглашается с трактовкой Боллса.
Если за каждым подсвечиванием кнопки следует пища (и она не предъявляется ни в какое другое время), вскоре свет будет рассматриваться как имеющий отношение к еде. Небольшие, связанные с питанием объекты стимулируют клевания как наиболее предпочтительную деятельность (автоформирование). Когда механизм устроен так, что клевания вызывают подачу корма, происходит дальнейшее закрепление корреляции между светом и пищей... Ситуации, ассоциирующиеся с приемом пищи, вызывают действия, связанные с питанием.
Неадаптивные ограничения этого типа С-С научения показаны в работе Уильямса и Уильямса (Williams & Williams, 1969), которую мы обсуждали в главе 5. Напомним, что в данном эксперименте, направленном на автоформирование, клевание снижало уровень подкрепления, но, несмотря на это, удары клювом по кнопке продолжались. Таким образом, подсвеченный диск служил признаком появления пищи и вызывал связанное с приемом пищи поведение вне зависимости от последствий этого поведения. Как мы установили в начале этой главы, у эволюции нет глобального плана, направленного на прогресс. Приспособление, которое адекватно в своей собственной нише (или СЭА), может столкнуться с затруднениями в современной среде или лаборатории, где оно потерпит неудачу.
Эволюционная психология и человеческое поведение
Несмотря на то что при освещении эволюционной психологии мы делали акцент на исследовании животных, нужно отметить, что эволюционная психология широко применяется для объяснения человеческого поведения. В первой работе такого рода «Социобиология: новый синтез» (Sociobiology: the new synthesis, Wilson, 1975) Эдварда Уилсона принципы эволюции прежде всего применяются для трактовки социального поведения животных; человеческое же поведение упоминается вскользь. Однако в книге, получившей Пулитцеровскую премию, «О природе человека» (On human nature, Wilson, 1988) Уилсон рассматривает биологическую основу социального поведения человека, заявляя, что эволюционирует как мышление, так и культура, поскольку они обеспечивают выживание. В своей последней книге «Согласие: единство знания» (Consiliense: the unity of knowledge, Wilson, 1988) он выступает за объединение естественных, социальных и гуманитарных наук. В этом синтезе решающее место отдано эволюционной психологии. Роль психологии в новом синтезе была охарактеризована в докладе Уилсона в 1999 г. на национальном собрании Американской психологической ассоциации в Бостоне.
То, что Уилсон обозначал как социобиологию, теперь носит название эволюционной психологии и является чрезвычайно популярной темой в современной психологической науке. В дополнение к разработкам Уилсона (Wilson, 1988) существуют другие исследования в области применения эволюционной психологии к человеку (Buss, Haselton, Shackelford, Bleske & Wakefield, 1998; Crawford & Krebs, 1998; Hergenhahn & Olson, 1999). Хотя в последующем изложении мы ограничимся обсуждением влияния предварительного научения на развитие фобий, выбор партнера, воспитание детей, насилие в семье, «альтруизм» и нравственное поведение, речевое развитие, существуют другие типы поведения, к которым применимы эволюционные принципы. Сюда входят агрессия и жажда войны; изнасилование, инцест и суицид; избегание инцеста; религия.
По многим параметрам принципы объяснения человеческого поведения в терминах эволюции сходны с боллсовскими принципами применения эволюционных трактовок к поведению животных (Bolles, 1972,1988). Эволюционная психология полагает, что, несмотря на значительный прогресс, достигнутый человечеством, особенно за два последних столетия, мы по-прежнему являемся продуктом тысяч лет эволюции. Поэтому, подобно другим животным, мы иногда склонны проявлять именно врожденные предрасположенности реагирования на стимул и более готовы обучаться одним видам ожиданий в противоположность другим. Как и другие животные, мы склонны к врожденным тенденциям реагирования, особенно когда нами управляют сильные, биологически значимые мотивационные состояния. Иными словами, концепция готовности Селигмана подходит к человеческому научению, так же как и к научению животных. И наконец, аргумент ниши применим к нам, так же как и к другим животным. Более точное понимание человеческого поведения достигается, если соотнести современное поведение с СЭА этого поведения. Ни одно из этих утверждений не подразумевает, что люди находятся в рамках негибкого инстинктивного поведения. Однако они свидетельствуют о том, что эмпирический принцип эквипотенциальности неадекватен при объяснении человеческого научения, а также при интерпретации возможностей научения животных. Боллс (Bolles, 1988) говорил о врожденной структуре, управляющей научением, и эволюционные психологи ссылаются на его структуру как на биологически запрограммированную (Barash, 1979). В обоих источниках предполагается, что научением руководят природные склонности и оно возможно благодаря нашей эволюционной истории. Так или иначе, по мнению эволюционных психологов, следует избегать принципа «ничего кроме» любой ценой. То есть ошибочно полагать, что человеческое поведение не представляет собой «ничего кроме» культурально или «ничего кроме» биологически заданного (т. е. врожденного). С позиции эволюционной психологии и культура и биология должны быть приняты во внимание для более полного понимания человеческого поведения.
Развитие фобий
Нижеприведенная концепция развития фобий, предложенная Люмсденом и Уил-соном (Lumsden & Wilson, 1981), во многом согласуется с выдвинутой Селигма-ном концепцией готовности.
Готовность человека к научению наиболее отчетливо проявляется в случаях фобий, т. е. страхов, характеризующихся набором черт. Прежде всего их отличает экстремальность реакции... Они обычно вспыхивают со всей силой даже после единичного негативного подкрепления [и] угасают с большим трудом... Примечателен тот факт, что явления, вызывающие эти реакции (замкнутые пространства, высота, гроза, текущая вода, змеи и пауки), содержат одни из самых страшных опасностей, присутствующих в древней среде обитания человека, в то время как пистолеты, ножи, автомобили, электрические розетки и другие, куда более страшные опасности технологически развитых обществ оказывают меньшее воздействие. Разумно сделать заключение, что фобии являются крайними случаями иррациональных реакций страха, толкающими людей на экстренные меры для обеспечения выживания... Лучше отползти от утеса с головокружением от страха, чем пройти по его краю.
Предметом обсуждения эволюционных психологов также является ксенофобия, или боязнь незнакомых людей. Эта фобия, как они считают, берет свое начало в примитивных установках разделять всех людей на членов своей группы (клана, деревни, племени) и на не принадлежащих к группе. Члены группы подчиняются одним правилам и придерживаются одинаковых убеждений (например, религиозных принципов) и в целом воспринимаются как друзья; не принадлежащие к данной группе индивидуумы живут по другим правилам и принципам и воспринимаются как потенциальные враги.
В ксенофобии можно увидеть природную склонность к предрассудкам. Здесь необходимо сделать два важных предостережения. Во-первых, согласно эволюционным психологам, природное происхождение тенденции не означает ее автоматическую пользу. Предполагать, что природное является благом, — значит принимать натуралистический софизм. Во-вторых, эволюционные психологи не утверждают, что люди «неотрывно привязаны» к определенному стереотипу поведения. Как мы отмечали, они говорят о биологически заложенных склонностях или тенденциях. Бараш (Barash, 1979) называет их «внутренним голосом». Более того, как мы только что видели, эволюционные психологи настаивают на том, что человеческое поведение всегда является результатом взаимодействия биологических наклонностей и влияния культуры. Таким образом, даже если наша биология склоняет нас к действиям, которые считаются неприемлемыми, на них могут наложить запрет культуральные влияния. Бараш (Barash, 1986) говорит:
К счастью, есть хорошие новости. Человеческие существа, разумные приматы, которыми мы являемся, имеют возможность выбора. Мы можем преодолеть наши примитивные ограничения и недальновидность. Мы можем научиться всем видам сложных действий, если убеждены в их важности или неизбежности. Мы даже можем научиться вещам, которые идут вразрез с нашей природой. Примат, который сможет научиться пользоваться туалетом, при дальнейшем обучении может стать даже влиятельным лицом.
Выбор партнера
Хотя есть общества, в которых браки устраиваются другими людьми, обычно сами мужчина и женщина принимают активное участие в выборе партнера и ухаживании. Как мы выбираем из множества потенциальных партнеров одного (или нескольких), с которым мы производим потомство? Ответ прост: мы ищем продолжительных партнерских взаимоотношений с наиболее привлекательными для нас индивидуумами. Но как мы определяем, что привлекательно, а что нет? На некотором уровне анализа может показаться, что существует столько же стандартов красоты, сколько и культур; и даже внутри одной культуры стандарты внешней привлекательности могут существенно различаться. Приверженец социального когнитивного научения предположит, что понятие красоты складывается путем следования явно выделяющимся моделям, признанным в нашей культуре (родителями, сверстниками, лидерами и т. п.) и в технологических обществах из тех моделей, чью привлекательность пропагандируют СМИ. Однако с позиции эволюционной психологии многие стандарты, навязанные культурой, поверхностны. Многие общественные идеалы красоты претерпевают изменения; например, прически, нательные украшения, стиль одежды и даже предпочитаемые формы тела видоизменяются. По мнению эволюционного психолога, должны существовать универсальные для всех культур критерии выбора партнера, куда более основательные, чем социальные стандарты физической привлекательности внутри конкретной культуры.
Дэвид Басе, выдающийся исследователь в области выбора партнера, замечает (Buss, 1998):
Мы никогда не выбираем пару случайно. Мы не привлекаем партнеров беспорядочно... Мы планируем встречи и разрабатываем стратегии специально для решения определенных проблем, связанных с успешным браком... Те в нашем эволюционном прошлом, кто не был удачлив в выборе партнера, не стали нашими предками. Все мы произошли от длинной и непрерывной череды предшественников, которые победили в борьбе за желанных партнеров, привлекли особей, способных к репродукции, удержали партнера на достаточное для воспроизводства время, отразили натиски соперников и решили проблемы, которые могли бы препятствовать успешному воспроизводству.

Таким образом, с точки зрения эволюционной психологии привлекательный партнер будет обладать чертами, возможно, не имеющими отношения к физической привлекательности. Сюда можно включить ценность партнера как добытчика и защитника, репродуктивную совместимость, достоинство партнера в качестве родителя и т. д. Иными словами, привлекательный партнер будет иметь характеристики, свидетельствующие о том, что он поможет обеспечить нам выживание, воспроизводство и повысит выживаемость наших детей.
Басе и его коллеги опросили более 10 ООО человек из 37 различных культур, чтобы выявить наличие универсальных признаков, которые ценятся в потенциальных партнерах (Buss, 1989, 1994, 1998; Buss & Schmitt, 1993). Результаты неопровержимо свидетельствуют о том, что, несмотря на внесенное культурой разнообразие, эволюция выбрала черты, которые мы распознаем (скорее, чем научаемся) в качестве важных характеристик хорошего партнера. Как показано в табл. 15.1, самые главные характеристики, выделенные мужчинами и женщинами, это доброта и понимание, вслед за ними интеллект и другие факторы, способствующие выживанию партнера, нас и нашего потомства.

Можно принять к сведению сходство критериев у мужчин и женщин с двумя важными исключениями. Мужчины имеют тенденцию ставить «физическую привлекательность» выше, нежели женщины, а женщины, в отличие от мужчин, дают более высокую оценку «способности хорошо зарабатывать». С позиций эволюционной психологии объяснение этих различий таково: женщины расходуют значительное количество биологических ресурсов при вынашивании и рождении ребенка, и до недавних пор женщины использовали свои биологические ресурсы как единственный родитель, способный выкормить новорожденного. Поэтому женщины придают большее значение способности мужчины защищать и обеспечивать нуклеарную семью, пока ее ресурсы (и силы) направлены на другое. И наоборот, мужчины должны уделять внимание физическим данным, свидетельствующим о хорошей репродуктивной способности женщины. В подтверждение этой гипотезы было обнаружено (Buss, 1989; Buss & Schmitt, 1993), что женщины больше, чем мужчины, ценят такие факторы, как амбиции партнера, трудолюбие, социальный статус и силу. С другой стороны, мужчины больше женщин обращают внимание на возраст партнера, состояние здоровья, чистую кожу и ясность взгляда.
Воспитание потомства
Хотя специфические роли, играемые родителями в образовании, социализации и воспитании своих детей, формируются под влиянием культуры, в них также присутствует влияние биологии. Например, Тайгер (Tiger, 1979, р. 96) отметил, что родительство влечет за собой жертвование времени и средств для «ряда крайне бесполезных и весьма неудобных занятий». Задача эволюционной психологии состоит в объяснении того, почему двое рациональных взрослых людей способны к истощению своих биологических и физических ресурсов (ставя, таким образом, свою жизнь под угрозу) ради других, которые, в чем убедятся родители, редко (если вообще когда-нибудь) говорят «спасибо» и могут не заметить значительность альтруистичных и малоприятных действий, долгие годы совершавшихся их родителями.
Выбор по родству. Эволюционная трактовка родительства начинается с неодарвинистского принципа родственного выбора, идеи о том, что эволюционная приспособленность включает в себя увековечивание не только наших собственных генов, но и генов наших родственников (включающая приспособленность). Правило Гамильтона (Hamilton, 1964) отводит ведущую роль родственным генам, так как они имеют отношение к альтруизму, представляющему собой совершение жертвенного поступка в пользу другого человека. Правило гласит, что альтруистическое поведение имеет место, когда гВ > С, где:
В = выгода реципиента от альтруистического поступка;
С = затраты человека, который совершил альтруистический поступок;
г = соотношение генов совершающего альтруистический акт и его реципиента.
На самом простом примере эгоизма рассмотрим действие, которое кто-либо совершает для своей же пользы. В этом случае г = 1,00; то есть мы разделяем 100% генов на самих себя. Согласно правилу Гамильтона, мы склонны поступать подобным образом для своей выгоды, даже когда выгода едва превышает затраты. Поэтому возможность получить выгоду всегда довольно высока. Половина генов ребенка родительские, т. е. г = 0,50. Это означает, что выгода от альтруистического поступка должна более чем в два раза превышать затраты на него. Поэтому мы скорее склонны к альтруизму по отношению к себе, чем к нашим детям. На самом деле до тех пор пока отношение выгоды к затратам не будет велико, а так произойдет в случае, если здоровье или жизнь ребенка окажутся под угрозой, правило Гамильтона говорит о том, что мы будем поступать эгоистично. Если мы включим других родственников в уравнение, мы начнем понимать смысл правила Гамильтона. Наши братья, сестры и родители также разделяют 50% наших генов, но племянники, дяди, тети и бабушки с дедушками разделяют только 25% наших генов. И в соответствии с правилом, чтобы совершить альтруистический акт ради этих родственников, выгода должна превышать затраты в четыре раза. Поэтому мы более всего склонны совершать альтруистические поступки ради нас самих, затем ради отпрысков и нуклеарной семьи и в последнюю очередь — для более отдаленных родственников. Чем меньше генов мы разделяем с другим индивидуумом, тем меньше вероятность неэгоистичного поведения, и вероятность совершения альтруистического поступка ради неродственника является нулевой.
В этом смысле эволюционные психологи смотрят на родительство как на действия, подчиняющиеся принципам выбора по родству. Нашим отпрыскам достается выгода, поскольку они среди тех, кто скорее всего будет реципиентом наших неэгоистичных поступков. Как считает Кребс (Krebs, 1998, р. 353), родители «просто делают то, что должны для того, чтобы распространить свои гены. Говоря на языке генетики, они помогают сами себе».
Половые различия. Бараш (Barash, 1979) отмечает, что родительство было и остается теперь в основном женским делом.
Не существует человеческого общества, в анналах истории или в современности, в котором женщина не несла бы главной ответственности за заботу о ребенке. Родительство во многом связано с полом. Во всех культурах мужчины выполняют мужскую работу, удел женщины — уход за ребенком. Но почему так происходит? Поскольку по половине генов индивидуума закладывается обоими родителями, каждый из родителей должен быть одинаково заинтересован в каждом ребенке. Так ли это на самом деле? Нет.
По мнению эволюционных психологов, есть две основные причины, почему женщины занимаются воспитанием детей больше, чем мужчины. Во-первых, женщины сильнее, чем мужчины, заинтересованы в своем потомстве. Бараш (Barash, 1979) объясняет:
Яйцеклетка оплодотворяется спермой, а не наоборот. Беременеет женщина, не мужчина. Именно женщина должна вырабатывать плаценту и кормить плод; она вынашивает эмбрион, который растет и прибавляет в весе, делая женщину все более неуклюжей по мере развития беременности; и когда ребенок рождается, она должна ухаживать за ним. Поскольку женщины беременеют, они просто не могут воспроизвести столько детей, сколько способны зачать мужчины. Мы можем сожалеть об этом факте, гордиться им или просто принять, но, несмотря ни на что, он остается неотъемлемой частью нашей биологии.
Во-вторых, для того чтобы поведение самопожертвования следовало правилу Гамильтона, должны существовать механизмы, с помощью которых мы узнаем родственников, включая наших собственных детей как носителей наших генов. Полагают, что половые различия в воспитании детей отчасти возникают потому, что мужчины и женщины должны полагаться на разные признаки для решения проблемы определения отцовства (или материнства). Для матерей это простой вопрос. Басе (Buss, 1988, р. 415-416) отмечает: «Нет женщины, которая, дав жизнь ребенку, сомневается в том, что ребенок, появившийся из ее тела, ее ребенок. И напротив, мужчина никогда не может быть в этом уверен». Поэтому женщины более склонны к выполнению обязанностей родителя, нежели мужчины. Это не значит, что мужчины не способны идти на жертвы ради своих детей. Однако они должны полагаться на такие признаки, как фенотипическая (физическая) внешность, менее очевидные, чем те, на которые опирается мать (Krebs, 1988).
Насилие в семье. Важным заключением из правила Гамильтона и выбора по родству является тот факт, что жестокое поведение вряд ли будет направлено против тех, кто имеет общие с нами гены. Следовательно, насилие в семьях должно происходить редко. Тем не менее широко известно, что насилие в семьях происходит ежедневно. Геллз и Страус (Gelles & Straus, 1985, p. 88), к примеру, предупреждали, что если не рассматривать военных и защитников правопорядка, индивидуум «скорее всего будет убит или ранен в своем собственном доме другим членом семьи, чем кем-либо или где-либо еще».
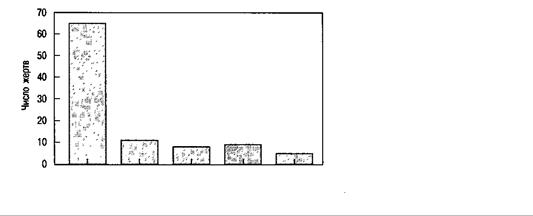
Дейли и Уилсон (Daly & Wilson, 1982, 1998) утверждают, что, несмотря на ежевечерние выпуски новостей и заявления таких исследователей, как Геллз и Страус, выбор по родству оказывает сильное влияние на насилие в семье. В частности, выбор по родству склоняет к жестокому поведению против членов семьи, которые не являются родственными генетически. Например, в компиляции статистики убийств в Детройте, показанной на рис. 15.2, Дейли и Уилсон обнаружили, что убийства более чем в 20 раз чаще совершаются в отношении супруга (не генетического родственника) или неродственного индивидуума, чем в отношении ребенка, родителя или других генетических родственников. Объединив данные различных культур, они пишут (Daly & Wilson, 1998):
Близкие генетические взаимоотношения гораздо более превалируют среди соучастников насилия, чем между жертвой и убийцей... Даже для патриархальных обществ, где братья поочередно наследуют семейные земли и титулы, характерно, что близкие ге-неологические связи смягчают конфликты и снижают количество прецедентов жестокого обращения... Единство семьи не может быть просто следствием родства и близкого знакомства (р. 440).
Представляется затруднительным трактовать отношения между детьми и приемными родителями с позиций правила Гамильтона. Пинкер (Pinker, 1997, р. 433) пишет: «Отчим сделал покупку для супруги, но не для ребенка; ребенок — это цена, которая является частью сделки». Дэйли и Уилсон (Daly & Wilson, 1998) представляют проблему следующим образом.
Адаптивно и нормально для биологических родителей подвергать риску собственные жизни, заботясь о ребенке, но селекция, вероятно, предполагает в воспитании приемных детей гораздо более низкие пороги допустимых затрат... В таком случае неудивительно, что эксплуатация и плохое обращение с приемными детьми является фольклорной тематикой во всем мире (р. 441).
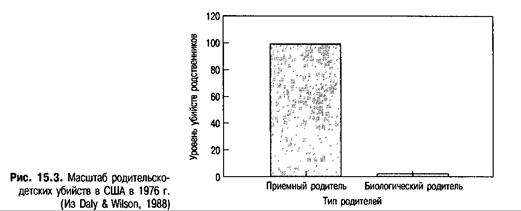
Но только ли это сказки? Действительно ли приемные дети скорее, чем биологические, выбираются в качестве мишени для насилия? Ответ: «Да». При исследовании убийств в Канаде, совершенных между 1974 и 1990 гг., Дэйли и Уилсон (Daly & Wilson, 1988, 1994) обнаружили, что приемные родители убивали детей, особенно от рождения до пяти лет, в 50-100 раз чаще, чем биологические. Эти драматичные факты отображены на рис. 15.3.
Альтруизм и нравственное поведение
Вид альтруизма, который мы рассматривали выше, называется родственный альтруизм; его проявления определены правилом Гамильтона. Эволюционные психологи также рассматривают обоюдный альтруизм, который представляет собой помощь со стороны генетически не связанных между собой индивидуумов. Обоюдный альтруизм основывается на факте того, что люди, которые сотрудничают, выживут с большей вероятностью, чем одиночки (например, на охоте или войне). Обоюдный альтруизм базируется на предположении о том, что если один член сообщества поможет другому, когда-нибудь в будущем тот или другой соплеменник ответит на доброту. Подобный альтруизм следует принципу: «Поступай с другими так, как хотел бы, чтобы поступали с тобой».
Почему при том, что успех в конечном счете определяется с точки зрения выживания наших генов и генов родственников, мы должны развивать стратегии сотрудничества, способствующие благополучию людей, с которыми у нас нет родственных связей? Почему поведение, на котором базируется «нравственность» — делиться с другими, помогать, сотрудничать, сострадать, — должно развиваться в первую очередь? Кребс (Krebs, 1998) дает следующий ответ.
Следование собственным интересам и эксплуатация других не являются эффективными межличностными стратегиями поведения по трем основным причинам. Во-первых, некоторые ресурсы находятся вне досягаемости человека, действующего в одиночку. Во-вторых, несдерживаемый эгоизм может разрушить систему кооперации, за счет которой он снабжается. В третьих, другие люди в процессе эволюции научились сопротивляться эксплуатации. В-сущности, индивидуумы в группах сотрудничества соглашаются, хотя не всегда осознанно и отчетливо, принять нравственные принципы взаимодействия для максимизации общей выгоды. Правила морали придерживаются этих стратегий, определяя вложения (т. е. обязанности), которые должен выполнять каждый человек, чтобы получить нечто взамен (т. е. права).
Среди стратегий нравственного поведения, принятых людьми для максимизации общей выгоды и сопротивления индивидуальной эксплуатации, можно выделить почитание власти, разработку системы правосудия и механизмы отслеживания «жульничества» (Krebs, 1998).
Важно отметить, что как родственный, так и обоюдный альтруизм не является подлинным альтруизмом. В обоих случаях «альтруистическое» поведение в конечном счете относится к увековечению копий чьих-либо генов и потому эгоистично. Бараш (1979, р. 135) говорит: «Настоящего искреннего перед Богом альтруизма просто не существует в природе».
Речь
Как мы видели в главе 5, Скиннер предполагал, что развитие речи можно объяснить с помощью принципов оперантного обусловливания. В главе 7 упоминалось, что Павлов подходил к проблеме речи, привлекая принципы классического обусловливания выше первого порядка. Хотя мы и не обращались к этой проблеме в главе 13, социально-когнитивную теорию научения Бандуры можно применить и к вопросу развития речи, предположив, что оно происходит за счет научения через наблюдение. Однако эволюционные психологи считают эти теории научения неполными, поскольку они не обращаются к различным данным, свидетельствующим о роли физической подготовленности организма в развитии речи. На самом деле обучение речи демонстрирует значение биологической подготовленности в человеческом научении, возможно, наиболее ярко, чем любое другое явление, которое мы рассматривали.
Высказываниям против принятых в классической теории научения объяснений речи положили начало два весомых контраргумента, выдвинутых Ноамом Хомским (Chomsky, 1959,1975), хотя важно отметить, что сам Хомский скептически относился к эволюционным объяснениям феномена речи (Chomsky, 1972, 1988). Во-первых, Хомский указывает на то, что дети скорее составляют собственные уникальные фразы и предложения, чем просто повторяют те слова и предложения, за которые они получили подкрепление. Также дети говорят такие слова, которые не были и в некоторых случаях не могли быть смоделированы в их среде. Например, было бы несвойственно ребенку выучить признание: «Да, я съел леденец. Я сделал это!» Во-вторых, Хомский утверждает, что у детей развивается понимание грамматики без какой-либо формальной инструкции. То есть они распознают и обобщают основные правила языка и способны понять как стандартные, так и нестандартные предложения и фразы. С позиции Хомского мозг отчасти является органом, созданным природой для использования и понимания языка. Поэтому в разговорах о научении речи не больше смысла, чем о необходимости научения для пользования руками, сердцем или любым другим биологическим органом.
Стивен Пинкер в своей книге «Инстинкт языка» (The language instinct, Pinker, 1995), оказавшей значительное влияние, переходит на следующую логическую ступеньку и приводит весомые доказательства отнесения «органа языка» Хомского к сфере эволюционной психологии. Во-первых, Пинкер утверждает, что существуют универсалии, характеристики, типичные для всех известных языков. Примем на веру то, что касается других языков; во всех языках есть разделение на прошлое, настоящее и будущее время, все имеют падежи и конструкции субъект/действие (причина-следствие). Также во всех языках присутствуют простые правила для образования множественного числа, даже при наличии исключений из этих правил. Во всех языках существуют относительно простые правила для преобразования утверждений в вопросы, но ни в одном известном языке вопросы не образуются путем изменения порядка слов в утверждении. Ранние исследователи в области лингвистики обнаружили целых 45 грамматических универсалий в 30 различных языках (Greenberg, 1963), и последующие исследования выявили гораздо больше (Hawkins, 1988). Это именно те данные, которые подтверждают заявление Хомского о том, что мозг снабжен врожденной «универсальной грамматикой».
Во-вторых, Пинкер утверждает, что дети биологически подготовлены к созданию грамматических структур даже при отсутствии моделей или руководства. Креольские языки демонстрируют подобную врожденную изобретательность (см. Bickerton, 1981, 1984, 1998). Когда рабочие, принадлежащие к нескольким различным культурам и грамматическим группам, живут вместе, например как работники фермы, они разрабатывают функциональный стенографический язык под названием пиджин. Язык пиджин обычно содержит существительные и глаголы из представленных языковых групп, но в нем не будет «согласующегося порядка слов, приставок или суффиксов, времени или других указателей на время или логику, структуры сложнее, чем простое предложение, и никакого способа обозначения кто, что и кому сделал» (Pinker, 1994, р. 34). Дети, растущие в языковой среде пиджин, не используют его для общения друг с другом. Скорее они создают сложные грамматические правила, изобретая несуществующую в языке родителей пиджин структуру, таким образом создавая новую языковую форму под названием креол. Что самое главное, грамматика, превращающая креол в настоящий язык, разрабатывается детьми, а не предлагается родителями или педагогами. Таким образом, дети биологически подготовлены к созданию грамматических систем, имея под рукой фундаментальные лингвистические инструменты, даже если грамматическая структура не входит в эти механизмы. Недавно были проведены наблюдения за преобразованием пиджина в креол в языке жестов глухих детей Никарагуа. В 1979 г. были созданы первые школы для глухих детей в Никарагуа, и дети, посещавшие их, разработали форму жестового языка пиджин (несмотря на то, что учебные планы делали акцент на чтение по губам и речевую деятельность). Второе поколение глухих учеников сделало скачок от пиджина до более грамматически сложного креольского языка жестов, и этот скачок был гораздо более быстрым по сравнению с тем, какой сделали люди, говорящие на языке креол и использующие только разговорную речь (Kegl & Iwata, 1989). Откуда берутся правила грамматики при отсутствии формального грамматического руководства и взрослых для моделирования сложных структур предложения, изменения времени глагола, местоимений и так далее? Пинкер согласен в этом вопросе с Хомским, что грамматическое научение биологически подготовлено.
И наконец, весомый аргумент за врожденную предрасположенность к речи будет включать в себя факт наследственного генетического механизма. Конечно, сложность понимания и использования речи делает невозможным предположение о том, что один ген или даже небольшое скопление генов лежат в основании феномена речи. Однако Пинкер предоставляет доказательства следующей замечательной концепции — генетической неспособности выучить грамматические структуры. Специфическое нарушение речи (СНР) (Gopnik, 1990; Gopnik & Crago, 1991) — это наследственное расстройство, которое проявляется в замедленном развитии речи, неразвитой артикуляции и грамматических ошибках при отсутствии указаний на общую умственную дисфункцию. Например, четырехлетние дети в норме успешно проходят Wug-mecm. Он заключается в том, что детям показывают линейный рисунок несуществующего животного и сообщают, что оно называется Wug. Затем детям показывают изображения двух (или более) таких существ. Следовательно, там находятся два...? Четырехлетние дети без отклонений обычно пишут в бланке слово Wugs. Дети и взрослые с СНР не могут решить подобного рода задачу правильно, но они не испытывают затруднений при решении математических или неязыковых задач.
Хотя Пинкер (Pinker, 1994) признает, что многое еще предстоит узнать об эволюции языка, развитии речи и роли человеческого мозга в данном явлении, он с энтузиазмом защищает перспективность эволюционной психологии:
Итак, мы знаем, какие биологические характеристики участвуют в естественном отборе, а какие относятся к другим эволюционным процессам. Что можно сказать о языке? По моему мнению, вывод напрашивается сам собой. Каждая дискуссия... подчеркивала приспособительную сложность речевого инстинкта. Он состоит из многих частей: синтаксис, с его комбинаторной системой построения структуры фразы; морфология, вторая комбинаторная система построения слов; большой словарный запас; голосовой аппарат; фонологические правила и структуры; восприятие речи; алгоритмы грамматического разбора; алгоритмы научения. Физиологическим механизмом реализации данных составляющих являются нервные контуры, образованные рядом определенно следующих во времени событий. Благодаря нервным контурам открываются немыслимые возможности: способность передавать бесконечное число четко структурированных мыслей из головы в голову, модулируя выдыхание воздуха.
Оценка эволюционной психологииж. Вклад в развитие науки
Эволюционные психологи провели различие между предварительными объяснениями и окончательными объяснениями поведения. Предварительные объяснения включают условия депривации, наблюдаемый стимул среды, вероятность подкрепления и непосредственную историю научения организма. Окончательные объяснения делают акцент на характеристиках и стереотипах поведения организмов, которые сформировались в процессе естественного отбора. Большинство теорий научения, рассмотренных в данном издании, подчеркивают первое и не обращают внимания или полностью игнорируют последнее. Возможно, самым важным вкладом Боллса и других эволюционных психологов является то, что они настаивали на равном учете как примерных, так и окончательных объяснений. Отдавая дань Боллсу, Гарсиа (Garcia, 1997, р. 13) замечает: «Боллс сделал шаг вперед по сравнению с Толменом. Контекстуальная карта и последующие чувственные впечатления должны соответствовать эволюции конкретного вида под наблюдением». Боутон и Фэнслоу (Bouton & Fanselow, 1997) расширили представление о вкладе Боллса в науку.
Его подход был скорее молярным, чем атомистичным или редукционистским. Боллс расширил ориентированный на цель подход Толмена, рассматривая поведение с точки зрения как его непосредственной или ближайшей цели, так и конечной или эволюционной. Он всегда помещал поведение в контекст его функции... Невозможно достигнуть полного понимания поведения вне его функционального контекста.
Это не значит, что эволюционная психология облегчает задачу психологов. Плоткин (Plotkin, 1998) пишет:
Итак, когда к каузальному объяснению примешивается естественный отбор, подобное толкование становится гораздо более сложным, чем было, поскольку непосредственные причины не исчезают. Вместо этого к ним прилагается длинный ряд других причин... Это не просто более значительная история причин, которую необходимо рассказать. Она еще и более законченная.
Польза от этой более полной истории должна быть очевидна. Мы видели, как результаты исследований, нарушающие известные принципы научения, интерпретируются за счет привлечения эволюционного объяснения. Озадачивающие «исключения из правил» оборачиваются и не исключениями вовсе. Также эволюционная психология выполнила важную эвристическую функцию. Она стимулировала возникновение новых вопросов для изучения, многие из которых фокусируются на феномене человеческого научения, приближая нас к более полному пониманию научения и человека и животных.
Критика
Возможно, самой типичной критикой в адрес эволюционной психологии и теории эволюции как таковой будет утверждение о том, что эволюционные аргументы представляют собой замкнутый круг. То есть критики заявляют, что успешная адаптация определяется по тем физическим или поведенческим характеристикам, которые отсеиваются в процессе естественного отбора (и передаются по наследству); поэтому если поведение имеет место в текущем поколении, оно было отобрано и, следовательно, способствует успешной адаптации. Дискуссия, упомянутая ранее (о надстройках и коадаптации), показывает, что эволюционные психологи избегали ловушки адаптации и проблемы замкнутого круга.
Второе критическое положение касается того, что эволюционное объяснение поведения избирает доктрину о генетическом детерминизме. То есть если мы представляем собой результат генетического отбора, мы обречены быть продуктами наших эгоистичных и жадных генов. Однако, как мы уже видели, эволюционная психология не принимает принцип «ничего кроме». Петрайнович (Petrino-vich, 1997, p. 23) указывает на то, что эволюционная психология «не постулирует генетическую обусловленность, поскольку эволюционно детерминированные черты могут измениться, если среда, в которой развивается индивидуум, модифицируется. Сердцевиной современной социобиологии и эволюционной психологии является их широкое взаимодействие».
В-третьих, критики опасаются, что эволюционная психология представляет возврат к социальному дарвинизму, учению, которое оправдывает семейственность, расизм и, возможно, даже селекцию для людей. Как мы отмечали ранее, нравственное поведение, включающее в себя доброту к посторонним людям и увеличение помощи людям, не связанным с нами родством, эволюционировало, поскольку в наших интересах совершать подобные действия. Вновь Петрайнович (Petrinovich, 1997) выступает в защиту эволюционной психологии:
Так или иначе важность совокупной приспособленности не означает, что люди предназначены для поиска выгоды своему роду и друзьям в ущерб всем другим людям, обрекая человечество на среду, состоящую из «нас» и «их». Она всего лишь предполагает наличие склонности к общению и сотрудничеству преимущественно со знакомыми, чем с незнакомцами. Сам факт существования пристрастий не означает, что люди безнадежно обречены погрузиться в глубины ксенофобии.
В-четвертых, критики убеждены, что генетическая предрасположенность исключает научение. В сущности, говорят они, если поведение является результатом генетических процессов, ему не обучаются. Ситуации просто вызывают поведение; таким образом, поведение описывается как кластеры безусловных рефлексов. Однако, как мы видели, эволюционная психология утверждает, что эволюционные влияния направляют научение. Отклоняя эмпирический принцип эквипотенциальности, эволюционная психология придерживается мнения, что научение сдерживается врожденными факторами, но это не значит, что оно вообще не происходит. Как говорит Пинкер (Pinker, 1994, р. 410): «Эволюционная психология не пренебрегает научением, но пытается объяснить его... не существует научения без какого-либо врожденного механизма, благодаря которому научение состоится».
Вопросы для обсуждения
1. Обсудите главные положения эволюционной психологии.
2. Кратко изложите теорию эволюции Дарвина.
3. Объясните, почему коадаптации и надстройки свидетельствуют против мнения, что все приспособления были обретены в результате естественного отбора.
4. Проведите различие между совокупной приспособленностью и более узким определением приспособленности Дарвина.
5. Каким образом Боллс расширил теорию научения ожиданиям Толмена, включив в нее принципы эволюции?
6. В чем заключается эмпирический принцип эквипотенциальности? Почему приверженцы эволюционной теории не согласны с ним?
7. Кратко опишите аргумент ниши Боллса. Включите в ваше описание понятие о среде эволюционной адаптации (СЭА).
8. Приведите примеры того, как биология ограничивает инструментальное, оперантное и классическое обусловливание. Включите в свой ответ обсуждение понятия видоспецифических защитных реакций (ВСЗР), введенного Боллсом.
9. Объясните автоформирование с позиции эволюционной психологии.
10. Обсудите следующие категории человеческого поведения в контексте подготовленного научения: развитие фобий, выбор партнера, воспитание детей, насилие в семье, альтруизм и нравственное поведение.
11. Объясните различие между альтруистическим поведением ради родственников и взаимным альтруизмом и приведите пример к каждому.
12. Обсудите развитие речи с позиции эволюционной психологии.
13. Каков вклад эволюционной психологии в понимание процесса научения? За что критиковали данный подход?
Часть VII
Дата: 2019-04-23, просмотров: 625.