Г лава 14
Дональд Олдинг Хебб
Хебб родился 22 июня 1904 г. в городе Честер канадской провинции Новая Шотландия. Мать и отец Дональда были врачами. Его мать получила медицинскую степень в 1896 г. в университете Далхаузи в Галифаксе, Новая Шотландия, став третьей женщиной-врачом в этой провинции.
В 1925 г. в Университете Далхаузи Хебб получил степень бакалавра искусств. При этом его оценки были самыми низкими на курсе, и, по сути дела, он балансировал на грани между завершением обучения и исключением. Впоследствии Хебб стал одним из выдающихся исследователей и теоретиков психологии. Как мы видим, в его случае низкая успеваемость во время обучения в университете не сыграла решающей роли. По окончании университета Хебб работал учителем в своем родном городе. В возрасте 23 лет он познакомился с работами 3. Фрейда и понял, что в психологии существует множество областей для исследования. Заведующий кафедрой психологии университета Мак-Гилл в Монреале был другом матери Хебба, поэтому он согласился принять молодого человека в качестве студента-заочника, несмотря на низкие оценки в дипломе. Параллельно с обучением на кафедре психологии Хебб продолжал преподавать в начальной школе. Пытаясь видоизменить традиционные способы обучения, он с переменным успехом провел серию психологических экспериментов. В одном из них Хебб решил отказаться от использования в качестве наказаний дополнительных домашних заданий, поскольку это могло сформировать у учеников отрицательное отношение к учебе.
В то же время он считал, что для поддержания порядка в классе какая-то форма наказания все же необходима. В качестве меры воздействия он решил использовать ремешок, которым несильно ударял по рукам ученика при нарушении дисциплины. Хебб (Hebb, 1980, р. 282) описывает случай, когда такой способ дисциплинарного воздействия обернулся против него самого: «Однажды я принялся наказывать своего ученика, но мальчишка увернулся, и конец ремешка скользнул мимо его рук и ударил меня по брюкам на уровне головки пениса. Было чертовски больно, и я сказал мальчику: "Я страдаю сильнее, чем ты". Но не думаю, что он понял шутку».
Наряду с попытками реформировать систему образования Хебб в молодости пробовал писать рома-Дональд 0. Хебб (фотография ны и зарабатывать этим на жизнь. Однако, как и Криса Пейна) Б. Ф. Скиннер, потерпел на этом поприще поражение.
В университете Мак-Гилл обучение основывалось на традициях школы И. П. Павлова. В 1932 г. Хебб защитил диссертацию, основанную на представлениях павловской школы, и получил степень магистра гуманитарных наук. Будучи воспитанным на теории Павлова, Хебб все же видел ее ограничивающие рамки и ставил под сомнение ее значимость. Еще во время обучения в университете Мак-Гилл Хебб прочел «Гештальт-психологию» Вольфганга Кёлера и познакомился с работами Карла Лэшли, посвященными физиологии мозга (которые мы кратко рассмотрим ниже). В 1934 г. Хебб принимает решение продолжить обучение в Чикагском университете. Здесь он работает с К. Лэшли и посещает семинары Кёлера. Результаты экспериментов Лэшли поставили под сомнение общепринятое на тот момент времени представление о мозге как сложном коммутаторе. Синтетической концепции мозга придерживались в основном бихевиористы, например Торндайк, Халл и Уотсон, и ассоцианисты, например Павлов и Газри. Сторонники целостной концепции полагали, что определенные сенсорные события стимулируют определенные области мозга, вызывая специфические реакции. Научение приводит к изменению нервных импульсов, и сенсорные события стимулируют появление реакций, отличных от наблюдаемых ранее. Исследования Лэшли на крысах поставили под вопрос целый ряд положений синтетической концепции мозга. Самым поразительным результатом его исследований стал вывод о том, что локализация разрушенного участка мозга менее важна, чем объем разрушений. Этот вывод лег в основу предложенного Лэшли принципа общей активности. Согласно этому принципу распад научения и памяти усиливается по мере увеличения объема разрушений в коре мозга вне зависимости от местоположения разрушенных структур. Лэшли делает вывод, что кора мозга в процессе научения функционирует как единое целое. Если разрушается один участок коры мозга, другие части могут взять на себя функции разрушенного участка. Такую способность одних частей мозга принимать на себя функции другого участка Лэшли назвал эквипотенциальностью. Таким образом, принцип общей активности гласит, что объем нарушений научения и памяти является функцией количества нарушений в коре мозга. А эквипотенциальность означает, что местоположение разрушенного участка коры не имеет значения.
Без сомнения, выводы Лэшли вступали в противоречие со знаниями, полученными Хеббом в университете МакГилл. Со временем неприятие отдельных постулатов теории Павлова переросло у Хебба в полное отрицание. «Мое рвение было подобно усердию перевоспитавшегося алкоголика, посещающего собрания в обществе трезвости. Будучи ранее убежденным сторонником Павлова, я стал не менее рьяным приверженцем гештальта и Лэшли» (Hebb, 1959, р. 625). И вновь мы сталкиваемся с важнейшей характеристикой хорошего ученого: с желанием изменить собственное мнение.
В 1935 г. Лэшли стал профессором Гарвардского университета и пригласил Хебба присоединиться к нему. В 1936 г. Хебб получил в Гарварде степень доктора философии и остался в университете еще на год в качестве преподавателя и научного сотрудника.
В 1937 г. Хебб становится сотрудником Монреальского неврологического института и начинает работать вместе с известным нейрохирургом Уайлдером Пен-филдом. Задачей Хебба было исследование психического состояния пациентов Пенфилда, подвергшихся нейрохирургическому вмешательству. К великому своему изумлению, Хебб обнаружил, что даже после потери значительной части ткани лобных долей мозга умственные способности пациентов сохранялись на прежнем уровне, а в некоторых случаях даже немного улучшались. Иногда потеря мозговой ткани достигала 20% массы мозга. Эти наблюдения еще больше укрепили его сомнения относительно справедливости целостной концепции мозга и убедили его в том, что мозг функционирует как единое целое. По словам Хебба (Hebb, 1980, р. 290), вопросы, порожденные этими наблюдениями, послужили толчком для его последующих исследований: «Я не мог найти ни одного признака нарушения интеллекта после удаления значительной части ткани лобных долей головного мозга... Именно эта проблема определила направление всей моей дальнейшей деятельности».
После пяти лет исследований пациентов доктора Пенфилда (1937-1942) Хебб ( Hebb , 1980, р. 292) пришел к выводу, который позднее стал важной частью его теории: «Детский опыт в норме приводит к развитию общих представлений, образа мыслей и способов восприятия, которые вместе образуют интеллект. Повреждения мозга в раннем детстве оказывают существенное влияние на становление интеллекта, однако во взрослом состоянии точно такое же повреждение не влечет за собой отрицательных последствий».
На том этапе своей деятельности Хебб сделал три предположения, которые позднее попытался объяснить с помощью своей теории.
1. Мозг не является простым синтезатором, как предполагают бихевиористы и ассоцианисты. Если бы это было так, разрушение значительного количества нервной ткани лобных долей мозга приводило бы к более значительным нарушениям интеллекта.
2. Интеллект является следствием опыта, и поэтому он генетически недетер-минирован.
3. Опыт, полученный в детском возрасте, более важен для развития интеллекта, нежели опыт, полученный в зрелом возрасте.

В 1942 г. Лэшли был назначен директором Лаборатории биологии приматов Роберта Йеркса в Оранж Парке, штат Флорида, и вновь позвал Хебба за собой. Во время работы в лаборатории Йеркса (1942-1947) Хебб исследовал эмоции и личностные особенности шимпанзе. Он также провел наблюдения, которые впоследствии легли в основу его нейрофизиологической теории научения и восприятия. Так, например, «[шимпанзе] пугались, а точнее сказать, испытывали ужас, при виде глиняной модели головы шимпанзе или... какой либо узнаваемой части тела шимпанзе или человека (например, головы или руки манекена)» (Hebb, 1980, р. 294). На основании этого и других наблюдений Хебб предложил собственное уникальное объяснение страха, которое мы рассмотрим далее в этой главе.
В 1948 г. Хеббу предложили место профессора на кафедре психологии университета Мак-Гилл, где он проработал до выхода на пенсию. В те годы физиологичеекая психология была не слишком популярна, и Хебб считал, что ему очень повезло с работой. По его мнению, нейрофизиологические объяснения научения были в то время непопулярны по двум причинам. Во-первых, по мере развития научной философии и накопления знаний о ее логике многие исследователи пришли к убеждению, что объяснение поведения в терминах физиологических событий подобно смешению мух с котлетами. Привлечение физиологических механизмов для объяснения внешних форм поведения требовало изменения уровня рассуждений, а выводы, сделанные на основании перехода от одного уровня к другому, нарушали каноны научной логики. Другими словами, предполагалось, что физиологические события, как и внешние проявления поведения, образуют самодостаточную систему, а связь между двумя системами оставалась неопределенной.
Во-вторых, само бихевиористское движение было, по сути, выступлением против интроспекции. Для бихевиористов единственным легитимным предметом обсуждения психологии было нечто осязаемое, то, что можно было потрогать и изучить. Ощущения, описываемые в ходе интроспекции, и физиологические события не удовлетворяли этому критерию. Концентрируя внимание на внешних измеряемых проявлениях поведения, бихевиористы исключали из своих исследований нейрофизиологию.
Во многом именно Хебб заставил научный мир считаться с нейрофизиологическими объяснениями научения. Он предположил (Hebb, 1949), что бихевиористское движение представляет собой всего лишь первый шаг на пути революционных изменений в психологии, ниспровергающих представления устаревших субъективных философских школ. По мнению Хебба, бихевиоризм настаивал на объективном исследовании внешних проявлений поведения, и в этом заключалась его сила. Но, ограничиваясь исследованием поведения, бихевиористы, образно говоря, выплеснули младенца вместе с водой, в которой его купали. Сегодня, по мнению Хебба, психология готова ко второй фазе революционных изменений, а именно к объективному исследованию когнитивных процессов. Как мы увидим далее, Хебб применил к исследованию этих процессов нейрофизиологический подход, и в то время он был единственным, кто сделал это. По мнению Хебба ( Hebb , 1959), очень важно, чтобы в дальнейшем психологическая наука не отказывалась от исследования когнитивных процессов.
Сегодня нам нужно больше узнать о процессах формирования или передачи идей, и о том, какую роль эти процессы играют в поведении. Необходимо не только сформировать максимально точные в разумных пределах гипотезы, но и проследить затем, каковы их следствия для поведения и не возникли ли эти следствия в процессе эксперимента. Конечно, если угодно, вы можете называть эти события процессами передачи, а не идеями или клеточными ансамблями, но позвольте нам заниматься делом и изучать их.
Среди многочисленных наград Хебба можно упомянуть 8 почетных докторских степеней, пост президента Канадской психологической ассоциации (1952), председательство в Американской психологической ассоциации (1959), медаль Уоррена (1958) и награду Американской психологической ассоциации за выдающийся научный вклад в психологию (1961).
Отказавшись от бихевиоризма, основанного на теории Павлова, Хебб начал атаку на это научное течение и не отступал с этих позиций уже до конца жизни. Его первой крупной работой стала книга «Организация поведения» (Organization of behavior, 1949). По странной случайности начальные буквы заглавия его книги (ОП) совпали с первыми буквами крупнейшей работы Б. Скиннера «Поведение организмов» (Behavior of organisms, Skinner, 1938), широко известной читателям под аббревиатурой ПО. Более поздняя работа «Потребности и С. Н. С. ((сенсорная нервная система)» (Drives and С. N. S. (conceptual nervous system), Hebb, 1955) продемонстрировала стремление Хебба «физиологизировать» психологические процессы. В прекрасно написанном «Учебнике психологии» (Textbook of psychology, Hebb, 1972) можно найти описание общих положений его теории. Книга «Психология: исследование науки» (Psychology: a study of a science, Hebb, 1959) содержит более подробное изложение теории Хебба. Подход Хебба диаметрально противоположен методам функционального анализа Скиннера, согласно которым С-Р связь устанавливается вне зависимости от внутренних событий в организме.
После выхода на пенсию в 1974 г. Хебб переехал на небольшую ферму около Честера в провинции Новая Шотландия, где он родился. Он сохранил душевное и физическое здоровье вплоть до самой смерти. Умер он 20 августа 1985 г. в больнице во время плановой хирургической операции на бедре (Beach, 1987, р. 187). Наиболее важные теоретические построения Хебба мы рассмотрим ниже.
Ограниченная и обогащенная среда Ограниченная среда
Мы уже упоминали о том, что, работая с Пенфилдом, Хебб пришел к выводу, что детский опыт для умственного развития более важен, чем опыт, полученный во взрослом возрасте. С помощью ряда экспериментов удалось продемонстрировать потенциально опасное действие сенсорно ограниченной среды на научение и развитие нервной системы в раннем возрасте. Немецкий офтальмолог фон Зенден (von Senden, 1932) исследовал взрослых людей, страдавших врожденной катарактой, но затем, после хирургической операции получивших возможность видеть. Оказалось, что участники исследования могли сразу же определить присутствие объекта, но были не способны идентифицировать его, пользуясь одним лишь зрением. Так, например, можно было ожидать, что пациент, получивший возможность видеть, с легкостью отличит круг от треугольника, сравнивая непрерывный плавный контур круга с прямыми сторонами и углами треугольника. Тем не менее для пациентов фон Зендена эта задача была очень сложна, а иногда и невыполнима. Кроме того, участники исследования испытывали большие трудности с запоминанием отличительных признаков различных геометрических фигур. Все эти факты указывали на то, что восприятие форм до некоторой степени является врожденным, но для того чтобы научиться отличать один объект от другого, человеку требуется зрительный опыт. Постепенно, после интенсивного обучения, ранее незрячие пациенты научились узнавать объекты окружающей их среды, и их процессы восприятия пришли в норму.
Остин Рейзен (Reisen, 1947) выращивал маленьких шимпанзе в полной темноте примерно до двухлетнего возраста. Когда наконец животных вывели на свет, они вели себя так, как будто были абсолютно слепы. Тем не менее через несколько недель они начали видеть, и их поведение постепенно стало таким же, как и у обезьян, воспитывавшихся в нормальных условиях. Хебб пришел к выводу, что и взрослые люди, участвовавшие в исследовании фон Зендена, и молодые шимпанзе Рейзена должны были научиться видеть.
Вывод о том, что ограничение раннего опыта влияет на развитие интеллекта и восприятия, подтверждается данными множества других исследований. Раннее научение необходимо даже для становления восприятия боли — явления, которое как полагали многие, исходя из его жизненной значимости, могло быть врожденным. В одном из исследований, проводившихся в лаборатории Хебба (Melzack & Thompson, 1956), было показано, что скотч-терьеры, выращенные в частичной изоляции, были нечувствительны к боли и менее агрессивны, чем их собратья, воспитывавшиеся в нормальных условиях.
Обогащенная среда
Если сильно ограниченная среда приводит к нарушениям развития и нормального функционирования, можно предположить, что среда, богатая сенсорными стимулами, напротив, стимулирует развитие. По-видимому, так оно и есть. Хебб провел один из первых экспериментов, изучающих влияние различных условий воспитания на умственное развитие подопытных животных (Hebb, 1949, р. 298-299). Опыт проводился на крысах, которых разделили на две группы. Одну группу животных выращивали в клетках в лаборатории Хебба, животные второй группы росли у Хебба дома под присмотром двух его дочерей. Эти крысы проводили значительную часть времени, передвигаясь по дому и играя с девочками. Через несколько недель «домашних» крыс вернули в лабораторию и сравнили с животными, выросшими в клетке. Оказалось, что домашние крысы значительно лучше справлялись с заданиями, связанными с поиском обходных путей и прохождением лабиринта, нежели грызуны, выросшие в лаборатории.
Результаты этой ранней работы Хебба были подтверждены данными многих других исследований. Так, например, в серии экспериментов, проводившихся в Калифорнийском университете Беннеттом, Даймондом, Кречем и Розенцвейгом (Bennett, Diamond, Krech & Rosenzweig, 1964), подтвердился вывод о том, что крысы, выросшие в обогащенной среде, обучались быстрее, чем их собратья, росшие в относительной изоляции. В этих экспериментах в качестве обогащенной среды использовалась просторная клетка с различными игровыми приспособлениями, в которой содержалась группа животных (см. рис. 14.1).
Сохраняется ли действие ограниченной среды на молодых животных в дальнейшем? По данным исследований Розенцвейга и его коллег, по-видимому, нет. Негативное воздействие бедной сенсорными стимулами среды в дальнейшем можно достаточно легко устранить, помещая животных в обогащенную среду на несколько часов в день. Таким образом, ущерб, нанесенный молодому животному ограниченными условиями среды, можно исправить, изменив условия содержания к лучшему. Другими словами, данная стадия развития не является критической, и ущерб, нанесенный ограниченной окружающей средой в раннем возрасте, может быть возмещен.
Хебб предложил простое объяснение этого явления. В обогащенной среде большее сенсорное разнообразие давало животным возможность создать большее количество более сложных по структуре нервных контуров или сети. Однажды сформированные нервные контуры в дальнейшем использовались при научении. Недостаточный сенсорный опыт в депривированной среде ограничивает количество нервных связей или вообще откладывает их формирование. Поэтому животные, выросшие в малостимулирующей среде, хуже справляются с решением поставленных перед ними задач. На основании этих сведений можно сделать очевидный вывод: чем сложнее сенсорное окружение маленького ребенка, тем успешнее он в дальнейшем будет справляться с решением встающих перед ним проблем.

Описанные выше наблюдения укрепили эмпирическую позицию Хебба. Интеллект, восприятие и даже эмоции являются результатом научения и опыта, а поэтому не наследуются, как полагали нативисты. Согласно предложенной Хеббом теории, у новорожденных детей в нейронных сетях количество связей между клетками невелико. Сенсорный опыт повышает уровень организации нейронных сетей и создает средства для эффективного взаимодействия с окружающей средой. Ключевыми понятиями теории Хебба и основными элементами предложенных им нервных контуров являются констелляции клеток и фазовые последовательности.
Констелляции клеток и фазовые последовательности. Констелляции клеток
Согласно Хеббу, каждый объект окружающей среды, с которым мы сталкиваемся, активизирует в мозге сложную группу нейронов, называемую констелляцией клеток. Так, например, когда мы смотрим на карандаш, мы переводим взгляд с острия на резинку и на деревянную рубашку карандаша. По мере смещения внимания активизируются различные нейроны. В то же время полная группа нейронов, задействованная в данный момент времени, соответствует одному объекту окружающей среды — карандашу. Поначалу все характеристики этой сложной группы нейронов независимы. Так, например, когда мы смотрим на острие карандаша, активизируется одна группа нейронов. Вначале нейроны этой группы не оказывают влияния на нервные клетки, активизирующиеся при переключении внимания на резинку или тело карандаша. Тем не менее в конце концов из-за небольшой разницы во времени между активизацией нейронов, соответствующих острию карандаша и других его частей, различные части активной группы нейронов оказываются взаимосвязанными. Сформулировав свои «нейрофизиологические постулаты», Хебб (Hebb, 1949, р. 62) предложил механизм, при помощи которого ранее независимые группы нейронов оказываются объединенными в стабильную констелляцию клеток: «Когда аксон клетки А готов возбудить клетку В и неоднократно (или постоянно) стимулирует ее, в одной или обеих клетках происходит некоторый рост или изменение метаболизма. В результате эффективность клетки А у как одной из клеток, активизирующих В, возрастает».
Хебб (Hebb, 1949) рассматривал констелляции клеток скорее как динамичные, нежели фиксированные или статичные системы нейронов. Он предложил механизмы, при помощи которых нервные клетки могут покидать констелляцию клеток или присоединяться к ней, позволяя этой структуре совершенствоваться в процессе научения и развития.
В предполагаемом процессе интеграции... обязательно должно происходить постепенное изменение частотных характеристик системы. В результате в системе возникает некоторая дробность и в то же время пополняется число клеток. Изменяются также и характеристики составляющих систему нейронов. Это означает, что некоторые клетки, синхронизированные с другими единицами системы, «выпадают» из нее и возникает «дробность». Другие нейроны, поначалу не входившие в систему, напротив, присоединяются к ней. Таким образом, по мере перцептивного развития наблюдается медленный рост констелляции клеток. При этом под «ростом» не обязательно подразумевается увеличение количества входящих в констелляцию клеток, но и любое изменение системы.
В зависимости от того, каков объект или событие окружающей среды, констелляция клеток может быть большой или малой. Так, например, констелляция клеток, связанная с дверной ручкой, будет состоять из относительно небольшого количества нейронов; констелляция, связанная с домом, будет включать в себя большое количество клеток. Констелляция клеток — это взаимосвязанная неврологическая система, которая может активизироваться посредством внешней или внутренней стимуляции, а также комбинации двух последних. Когда констелляция клеток возбуждается, у нас возникает мысль о событии, которое представляет это сообщество нейронов. Для Хебба констелляция клеток — это неврологическая основа идеи или мысли. Таким образом, Хебб объясняет, почему дома, коровы или наши любимые не обязательно должны находиться перед нашими глазами для того, чтобы мы думали о них.
Фазовые последовательности
Различные признаки одного и того же объекта оказываются неврологически связанными и формируют констелляцию клеток. Точно так же неврологическая связь может возникать между отдельными констелляциями, которые вместе образуют фазовые последовательности. Фазовая последовательность — это «временно связанная между собой серия форм деятельности констелляции клеток; она эквивалентна одной "струе" в потоке мыслей» (Hebb, 1959, р. 629). Однажды возникшая фазовая последовательность, как и констелляция клеток, может активизироваться изнутри, извне или посредством комбинации этих двух видов стимуляции. Когда возбуждается одна констелляция клеток или комбинация констелляций в составе фазовой последовательности, как правило, активизируется вся фазовая последовательность. При активизации фазовой последовательности у человека возникает поток мыслей — серия идей, организованных в некотором логическом порядке. Этот процесс объясняет, каким образом запах духов или несколько аккордов любимой песни могут пробудить воспоминания о любимом человеке. Хебб (Hebb, 1972) следующим образом описывает развитие фазовой последовательности.
Если констелляции активизируются одновременно, между ними устанавливается связь. Повседневные события в жизни ребенка приводят к возникновению констелляций. Когда эти события происходят одновременно, между констелляциями устанавливается связь (поскольку все они активны). Например, если ребенок слышит шаги, возбуждается определенная констелляция клеток. Пока эта группа нейронов активна, малыш видит лицо и чувствует, как чьи-то руки поднимают его вверх. Это приводит к активизации других констелляций. Таким образом «констелляция шагов» оказывается связанной с «констелляций лица» и «констелляцией рук». Впоследствии, когда ребенок слышит шаги, возбуждаются все три констелляции нейронов. Малыш как бы воспринимает лицо матери и прикосновение ее рук до того, как она появляется в поле зрения. Поскольку сенсорной стимуляции еще не было, то, что испытывает ребенок, можно назвать формированием идеи или образа, а не восприятием.
Согласно представлениям Хебба, существует два типа научения. Один из них связан с медленным формированием констелляций клеток в раннем возрасте. Его можно объяснить с помощью теорий научения С-Р, например теории Газри. Научение этого типа представляет собой прямой ассоцианизм. Аналогично формирование фазовых последовательностей можно объяснить в терминах ассоцианизма. Так, объекты и события, связанные в окружающей среде, оказываются связанными на неврологическом уровне. После формирования констелляций клеток и фазовых последовательностей научение приобретает когнитивный характер и происходит значительно быстрее. Научение в зрелом возрасте, например, зачастую характеризуется инсайтом и креативностью и, возможно, связано с перегруппировкой фазовых последовательностей. Так, Хебб утверждает, что переменные, влияющие на научение ребенка и взрослого, неидентичны. Научение в детском возрасте закладывает основы для дальнейшего обучения. Например, развитие речи — это медленный, сложный процесс, связанный, возможно, с построением миллионов констелляций клеток и фазовых последовательностей. Тем не менее, развив речь, человек может творчески преобразовывать его различными способами и создать, например, поэму или роман. Тем не менее, утверждает Хебб, вначале закладываются основы, и только затем приходит черед инсайта и креативности, характерных для научения в зрелом возрасте.
Теория активации
Все мы попадали в ситуации, когда из-за шума или суеты не могли ясно думать. С другой стороны, бывает, когда нужно сосредоточиться и работать на должном уровне. Подобные реакции предполагают, что не слишком слабая или слишком сильная по интенсивности стимуляция приводит к оптимальному когнитивному функционированию. Хебб исследовал связь между уровнем стимуляции и когнитивной деятельностью в контексте теории активации.
Теория активации связана с работой ретикулярной активирующей системы (РАС) — области размером с палец, расположенной в мозговом стволе над спинным мозгом, непосредственно под таламусом и гипоталамусом. Деятельность РАС связана с процессами сна, внимания и эмоционального поведения.

Согласно Хеббу (Hebb, 1955), нервные импульсы, причиной возникновения которых является стимуляция рецепторов органов чувств, имеют две функции. Одна носит название ключевой функции стимула. Сенсорный стимул порождает нервный импульс, который передается от рецептора органа чувств по чувствительному тракту спинного мозга к различным сенсорным зонам и наконец достигает определенной области коры головного мозга. Эта функция стимула позволяет организмам получать информацию об окружающей их среде. Вторая функция — это активирующая функция стимула. Было обнаружено, что от чувствительного тракта спинного мозга РАС отходят особые коллатеральные пучки. Сенсорная информация по пути к коре головного мозга через коллатеральные пучки воздействует на РАС и приводит к усилению ее активности. Такую способность сенсорного стимула увеличивать активность РАС и принято называть активирующей функцией стимула.
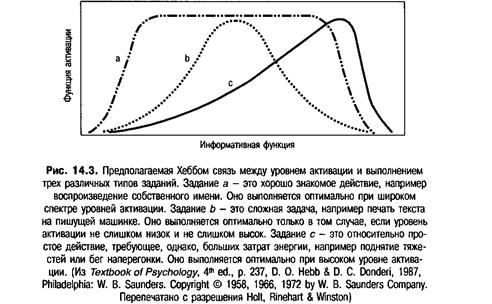
Хебб (Hebb, 1955) предложил теорию, объясняющую природу связи между уровнем активации и поведением. Он предположил, что, для того чтобы ключевая функция стимула была реализована полностью, необходимо, чтобы РАС создала оптимальный уровень активации. Если уровень активации слишком низок, как, например во время сна, сенсорная информация, поступающая в мозг, не может использоваться. Если же уровень активации слишком высок, кора мозга анализирует слишком большой объем информации, и в результате возникают конфликтующие ответные реакции и неадекватное поведение. Таким образом, для оптимального функционирования коры головного мозга и, как следствие, оптимального поведения необходим не слишком низкий и не слишком высокий уровень активации. Предполагаемая связь между оптимальным уровнем активации и поведением представлена на рис. 14.2.
Хебб предположил, что различные типы заданий характеризуются разными уровнями активации, необходимыми для их оптимального выполнения. Так, например, простое, хорошо знакомое действие может быть оптимально выполнено при различных уровнях активации. В то же время для оптимального выполнения сложной задачи допустим очень небольшой спектр уровней активации. Простейшие поведенческие стереотипы обычно проявляются при экстремально высоких уровнях активации. Предполагаемая связь между оптимальным выполнением различных действий и уровнем активации представлена на рис. 14.3.
Теория активации и подкрепление
По мнению Хебба, если уровень активации слишком высок и организм не может оптимально действовать в актуальных условиях, он будет стремиться понизить уровень активации. Так, например, если человек пытается заниматься, а люди вокруг него разговаривают или смотрят телевизор, он может попросить окружающих вести себя потише и выключить телевизор или просто попытаться найти место, более пригодное для занятий. С другой стороны, если вокруг слишком тихо и сенсорных сигналов недостаточно для поддержания оптимального уровня активации, можно включить радио, громко заговорить или выполнить какое-либо мышечное действие, например поерзать. Вообще говоря, когда уровень активации слишком высок, его понижение и является подкреплением. Ели он слишком низок, подкреплением является его повышение. В отличие от теории Халла, где подкрепление ослабляет потребность, теория Хебба в качестве подкрепления в зависимости от обстоятельств рассматривает как уменьшение, так и усиление потребности. По мнению Хебба (Hebb, 1955), стремление к возбуждению является очень важным побудительным мотивом поведения человека.
Если задуматься, диву даешься, в какие неприятности влезают люди, чтобы попасть еще в большие неприятности за карточным столом или на площадке для гольфа; столь же удивительной кажется притягательность детективных историй, триллеров и газетных статей о реальных приключениях и трагических событиях. Когда мы имеем дело с человеческой мотивацией, не следует забывать о стремлении к возбуждению. Оказывается, до определенного момента угроза и загадка имеют положительную мотивацион-ную ценность, после него ценность становится негативной (р. 250).
Эффект разрыва сформировавшихся фазовых последовательностей. Сенсорная депривация
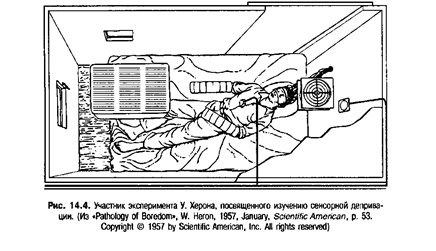
Мы уже говорили о том, что ограничение сенсорного опыта отрицательно сказывается на развитии нейрофизиологических констелляций, представляющих объекты и события окружающей среды. Но что происходит в случае ограничения сенсорного опыта уже после завершения нейрофизиологического развития? Ответ на этот вопрос был получен в серии экспериментов, проводившихся в Университете МакГилл под руководством Хебба. В одном из этих опытов (Heron, 1957) группе студентов колледжа платили $20 в день за то, чтобы они ничего не делали. Им нужно было только лежать на удобной кровати с полупрозрачной повязкой на глазах, позволявшей видеть рассеянный свет, но не дававшей возможности четко видеть объекты. Через наушники участники эксперимента постоянно слышали легкий шум. Чтобы в дальнейшем подавить восприятие звуков, в комнате монотонно жужжал кондиционер. На руки испытуемых надевали хлопчатобумажные перчатки и картонные обшлага, выступавшие за кончики пальцев и сводившие к минимуму тактильную стимуляцию. В таких условиях участники эксперимента находились круглые сутки за исключением перерывов на еду и посещение туалета. Условия проведения эксперимента показаны также на рис. 14.4.
Большинство участников эксперимента смогли выдержать такие условия в течение всего лишь двух-трех суток (самый стойкий участник продержался шесть дней). Как правило, студенты, участвовавшие в опыте, становились раздражительными и вели себя с экспериментатором почти по-детски. К великому удивлению Хебба и его коллег, сенсорная депривация производила эффект гораздо больший, чем просто скука. Хебб и Дондери (Hebb & Donderi, 1987) следующим образом подводят итог эксперимента Херона.
Эксперимент показал, что люди могут скучать, — это мы знали и ранее. Но он также показал, что скука — слишком слабое слово для обозначения воздействия сенсорной депривации. Потребность в нормальной стимуляции со стороны разнообразной окружающей среды — фундаментальная потребность организма. В отсутствие таковой нарушается умственная деятельность и разрушается личность. В условиях изоляции субъекты жаловались на потерю способности связно мыслить, хуже справлялись с простыми проблемами и начинали галлюцинировать. Некоторые видели шеренги маленьких человечков в черных шапочках, другие — белок, марширующих с рюкзаками на плечах, третьи — доисторических животных в джунглях. По описанию участников эксперимента эти видения были похожи на мультфильмы. Еще больше беспокойства доставляли галлюцинации, во время которых участникам эксперимента казалось, что их тело двоится или голова отделяется от тела... Разрушению начинала подвергаться сама личность человека.
Другим исследователям не удалось повторить некоторые наиболее сильные эффекты сенсорной депривации, полученные Хеббом (Suedfield & Coren, 1989; Zubek, 1969). Тем не менее в более поздних исследованиях было показано, что в жестких условиях сенсорной депривации участники исследования чувствуют себя плохо и могут переносить такое неприятное состояние лишь в течение короткого периода времени. Так, например, если участников исследования погружали в воду в полной темноте, позволяя им дышать через трубку, они, как правило, могли продержаться в таких условиях всего лишь несколько часов, а затем заявляли о желании прекратить эксперимент.
Из описанных выше исследований Хебб сделал вывод, что сенсорный опыт необходим не только для нормального нейрофизиологического развития, но и для поддержания нормальной деятельности. Другими словами, если события в жизни человека были нейрофизиологически зафиксированы в виде констелляций и фазовых последовательностей, они и в дальнейшем должны согласовываться с событиями окружающей среды. Если ранее обычные в жизни человека сенсорные события более не происходят, возникает сильное и неприятное возбуждение, которое воспринимается как стресс, страх или дезориентация. Таким образом, события окружающей среды необходимы не только для возникновения определенных нервных контуров. Те же самые события в дальнейшем поддерживают эти нервные связи. Следовательно, по мнению Хебба, к различным потребностям организма, например потребности в пище, воде, размножении и кислороде, следует добавить потребность в стимуляции. Даже если все остальные нужды организма удовлетворяются, отсутствие нормальной стимуляции ведет к сильнейшей когнитивной дезориентации.
Природа страха
Работая в лаборатории биологии приматов Йеркса, Хебб исследовал источники страха у шимпанзе. Он показывал подопытным животным различные тестовые объекты, например гипсовый слепок головы шимпанзе, куклу, изображающую человеческого ребенка, голову манекена, изображающего взрослого человека в натуральную величину, и маленького детеныша шимпанзе, находящегося под анестезией.
По наблюдениям Хебба, шимпанзе не выказывали никаких признаков страха до четырехмесячного возраста. После этого они не пугались хорошо знакомых или совершенно незнакомых им объектов. Страх вызывала лишь демонстрация знакомых объектов в незнакомом виде. Так например, целое тело шимпанзе или человека не вызывало у обезьян никакого страха, а модели отдельных частей тела человека или обезьяны приводили животных в ужас. Примеры объектов, вызывавших у подопытных животных страх, приведены на рис. 14.5.

Хебб полагал, что спонтанность страха исключает возможность объяснения поведения обезьян с точки зрения условно-рефлекторных реакций. В рамках подобного объяснения особое значение имело бы повторение демонстрации нейтральных объектов (например, головы шимпанзе) в сочетании с каким-либо неприятным стимулом. В этом случае страх вырабатывался бы медленно как результат опыта. Хебб наблюдал нечто совсем иное. В его случае шимпанзе сильнее всего пугались при первом появлении объекта. Хебб дал объяснение этого явления, используя понятия клеточных констелляций и фазовых последовательностей. Если организму показывают совершенно незнакомый объект, возникнет соответствующая ему констелляция клеток. При каждом последующем предъявлении объекта констелляция будет постепенно развиваться и страха не возникнет. Аналогично, если организму показывают знакомый объект, будут активированы нервные контуры, сформировавшиеся в результате предыдущего опыта, и поведение также не будет нарушено. Страх возникает лишь в том случае, если объект активизирует существующую констелляцию клеток или фазовую последовательность, но за предъявлением этого объекта не следуют характерные для него события. Так, например, вид находящегося под анестезией шимпанзе активизирует у подопытного шимпанзе фазовую последовательность, ассоциированную с видом нормального, активного животного. Однако события, в норме следующие за подобным восприятием, отсутствуют. Вместо типичных реакций и производимых шимпанзе звуков подопытное животное сталкивается с молчанием и отсутствием движения. Таким образом, фазовая последовательность активизируется, но не поддерживается сенсорными событиями, которые ранее привели к ее развитию. Отсутствие сенсорной поддержки, по мнению Хебба, вызывает страх. Таким же образом Хебб объяснил человеческую реакцию на покалеченные или мертвые тела. В результате Хебб (Hebb, 1946, р. 268) сделал в отношении страха следующий вывод: «Страх возникает, когда одни признаки объекта, попадающего в поле зрения организма, ему знакомы и запускают обычный процесс восприятия, а другие активизируют не сочетающиеся с восприятием процессы».
Предложенное Хеббом объяснение страха помогает объяснить травматичную природу сенсорной депривации. У взрослого человека формируется большое количество констелляций клеток и фазовых последовательностей, которые могут активизироваться за счет внутренней или внешней стимуляции, а также их сочетания. В условиях сенсорной депривации отсутствует сенсорная поддержка любых форм нервной деятельности. Таким образом, активизация многих нервных контуров не поддерживается сенсорными событиями, следующими за ней в норме. Неудивительно, что в подобной ситуации участники исследования дезориентированы и испытывают страх.
Долговременная и кратковременная память
Еще в 1900 г. Мюллер и Пильцэккер предположили, что существуют две различные формы памяти. Гораздо позже к этой идее обратился и Хебб. Он (Hebb, 1949) выделил постоянную память, в основе которой, по его мнению, лежат физико-структурные изменения нейронов, и недолговечную кратковременную память, которая существует за счет сохранения активности клеточных констелляций и фазовых последовательностей. В большинстве своем исследователи сегодня согласны с выделением двух основных форм памяти: долговременной и кратковременной памяти. Кроме того, они высказывают предположения о существовании нескольких разновидностей долговременной памяти. В этом разделе мы рассмотрим кратковременную память и обсудим доказательства существования двух видов долговременной памяти.
Принято считать, что сенсорный опыт приводит к возникновению нервной активности, которая длится дольше, чем вызвавшая ее стимуляция. Хебб называл ее реверберирующей нервной деятельностью. Хотя он признавал, что некоторые формы научения «возникают мгновенно и сохраняются очень долго» (Hebb, 1949, р. 62), он рассматривал реверберирующую нервную деятельность и как основу кратковременной памяти, и как процесс, приводящий к структурным изменениям, лежащим в основе долговременной памяти. Он также был одним из наиболее последовательных сторонников теории консолидации, предполагающей, что кратковременная память некоторым образом переносится в память долговременную.
Современные когнитивные психологи представляют себе кратковременную память примерно так же, как Хебб. По их мнению, кратковременная память — это относительно непродолжительная нервная деятельность, которая запускается сенсорной стимуляцией и продолжается некоторое время после прекращения стимуляции. Хебб предположил, что в случае фазовых последовательностей реверберация может продолжаться от одной до десяти секунд (Hebb, 1949, р. 143), но как долго длится кратковременная нервная деятельность, точно было неизвестно. Петерсон и Петерсон (Peterson & Peterson, 1959) предложили следующий алгоритм исследования кратковременной памяти. Они зачитывали участникам эксперимента триграмму согласных (например, QHJ), а затем просили людей считать в обратном порядке, отнимая по 3 или 4 от названного им трехзначного числа. У разных участников эксперимента счет прерывали через разные промежутки времени и просили воспроизвести зачитанную ранее триграмму. Были выбраны интервалы 3, б, 9,12,15 и 18 с. Оказалось, что лучше всего триграмма сохранялась в памяти, если интервал составлял 3 с, чуть хуже оказался результат при интерва ле в 6 с и т. д. Хуже всего участники эксперимента справлялись с воспроизведением триграммы, если интервал был 18 с. Таким образом, кратковременная память, по-видимому, быстро угасает в зависимости от времени.
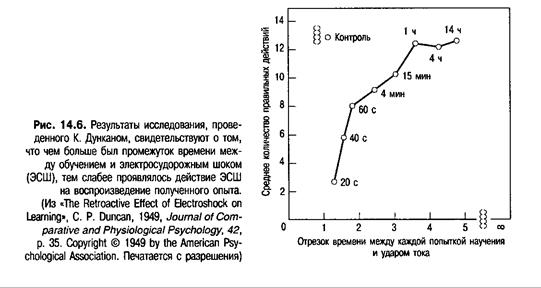
Долговременная память, как предполагается, зависит от консолидации (укрепления) кратковременной памяти. Поэтому любые нарушения последней будут отрицательно сказываться на способности организма длительное время сохранять накопленный опыт. Основываясь на этом утверждении, Дункан (Duncan, 1949) провел опыт, где учил крыс перепрыгивать через барьер, чтобы избежать удара током. Если они перепрыгивали с одной стороны экспериментальной клетки на другую в течение 10 с после помещения в испытательный стенд, они избегали удара током. Если же они не перебирались на «безопасную» сторону, они получали удары током до тех пор, пока не делали это. Животные совершали одну попытку научения в день. После ежедневного предварительного обучения каждого грызуна подвергали электросудорожному шоку (ЭСШ) через пару электродов, прикрепленных к ушам животного. ЭСШ приводит к судорогам, очень похожим на конвульсии во время эпилептического припадка. В зависимости от принадлежности к определенной группе животных подвергали ЭСШ через 20,40, 60 с, 4, 15 мин, 1, 4 или 14 ч в после каждого этапа предварительного обучения. Животных контрольной группы вообще не подвергали ЭСШ. Обучение длилось 18 дней. На рис. 14.6 показано среднее количество правильных действий, а именно перемещений с одной стороны клетки на другую после помещения в испытательный стенд.
Из графика видно, что чем меньше был промежуток времени между предварительным обучением и ЭСШ, тем сильнее нарушалась память о полученном опыте. Так, например, животные, подвергавшиеся действию ЭСШ через 20 с после предварительного обучения, вообще не могли воспроизвести реакцию избегания при следующем помещении в испытательный стенд. Если ЭСШ применялся в течение часа после попытки научения, он воздействовал на память животного. Если промежуток времени был более часа, то ЭСШ практически не оказывал влияния на память. Животные, подвергавшиеся действию ЭСШ через час или более после предварительного обучения, в последующих опытах справлялись с заданием так же хорошо, как животные контрольной группы, не испытывавшие ЭСШ. Результаты эксперимента Дункана поддерживают теорию консолидации и позволяют предположить, что период консолидации длится около часа. Тем не менее первые минуты сразу после научения, по-видимому, более важны для процесса консолидации, нежели более поздний промежуток времени.
Теорию консолидации также подтверждает явление ретроградной амнезии — исчезновение из памяти событий, предшествовавших травматичному опыту (например, аварии или ранению). Потеря памяти может затрагивать несколько часов, дней и даже месяцев перед травмой. Как правило, память об этих событиях медленно возвращается, за исключением нескольких моментов, непосредственно предшествовавших травме. Таким образом, действие травмы подобно действию ЭСШ, использованного в экспериментах Дункана.
Нарушают ли ЭСШ и другие травмы мозга консолидацию долговременной памяти из-за вмешательства в процессы нервной реверберации (в констелляциях клеток и фазовых последовательностях) или же воздействуют на нервные процессы, необходимые для консолидации, но не связанные с реверберацией? Этот вопрос нетривиален и особенно интересен в связи со случаем Г. М. — пациента, перенесшего хирургическую операцию и столкнувшегося с весьма специфичной проблемой консолидации.
Консолидация и мозг
За выражение различных эмоций в головном мозге ответственна группа взаимосвязанных структур, объединенных под названием лимбической системы. Одной из структур лимбической системы является гиппокамп. Бренда Милнер, одна из студенток Хебба, занималась случаем пациента, вошедшего в психологическую литературу под инициалами Г. М. (Milner, 1959, 1965; Scoville & Milner, 1957). Г. М. перенес хирургическую операцию на мозге по поводу мучившей его эпилепсии. Во время операции были повреждены структуры левой и правой частей гип-покампа и связанные с ними структуры лимбической системы. После операции у пациента развилась ярко выраженная антероградная амнезия. Он не испытывал затруднений при воссоздании событий, происходивших с ним до операции, но с большим трудом мог вспомнить что-либо, случившееся после нее. Пациенты со сходными с Г. М. симптомами хорошо справляются с тестами на интеллект; у них не нарушаются моторные навыки, приобретенные до повреждения гиппокампа. Милнер указывала также, что повреждение мозга не приводит к видимым изменениям личности. Такие пациенты ведут себя так, как будто их кратковременная память функционирует нормально. Но как только их внимание отвлекается от сиюминутного задания, память о нем исчезает. Пример Г. М. и подобных ему больных показывает, что для возникновения долговременной памяти недостаточно одной лишь реверберационной активности, в том числе и реверберации, порожденной простым повторением информации. Таким образом, считается, за консолидацию ответствен гиппокамп и, возможно, некоторые другие структуры мозга.
Проблемы Г. М. и других пациентов с нарушениями гиппокампа даже более сложны, нежели поначалу представляли себе исследователи. Пациенты с подобными мозговыми нарушениями способны научиться некоторым сложным действиям, но, по всей видимости, они пребывают в неведении относительно того, что научение имело место. Так, например, их навыки в области составления сложных мозаик или рисования в зеркальном отображении после тренировок улучшаются, что указывает на долгосрочное научение. В то же время сами пациенты могут утверждать, что они никогда не видели предлагаемых им заданий и не пытались выполнять их. Кроме того, они испытывают большие трудности с выполнением заданий, связанных с запоминанием списков и воспроизведением новых событий и фактов (Cohen & Eichenbaum, 1993; Cohen, Ryan, Hunt, Romine, Wszalek & Nash, 1999; Cohen & Squire, 1980; Squire, 1992). Для обозначения памяти, нарушенной у пациентов, подобных Г. М., исследователи используют термин декларативная память. Декларативная память (память о фактах) является составляющей памяти высшего порядка, а другая ее составляющая — процедурная память — отвечает за приобретение и удержание различных навыков деятельности. Повреждения гиппокампа и других структур серединной височной доли препятствуют консолидации декларативной памяти, но, как мы отмечали ранее, не нарушают долгосрочную память другого типа.
Считается, что базальные ганглии, группа нервных структур, принимают участие в контроле мышечных движений. Эта функция базальных ганглиев ярко видна на примере пациентов с ранними проявлениями болезней Гентингтона и Паркинсона, связанных с различными нарушениями данных мозговых структур. Мишкин с коллегами (Mishkin, Malamut & Bachevalier, 1984; Petri & Mishkin, 1994) указывают, что у пациентов с подобными нарушениями декларативная память не изменяется. Изменения затрагивают двигательную память, связанную со сложными моторными действиями, например составлением мозаик или рисованием в зеркальном отображении. Исследования последнего времени подтверждают эти выводы, обращая внимание на то, что двигательная память сильнее всего страдает у пациентов с болезнью Паркинсона (Thomas-Olliver, Reymann, LeMoal, Schuck, Lieury & Allain, 1999; Vakil & Herishanu-Naaman, 1998). В отличие от пациентов с повреждениями гиппокампа (как в случае с Г. М.) у этих больных навыки составления мозаик не улучшаются даже после длительной практики. И в то же время они осознают свои неудачи в научении.
Таким образом, для формирования долговременной памяти необходима консолидация, которая, однако, не является единым процессом. Результаты исследований говорят о том, что существует по крайней мере два типа долговременной памяти: декларативная и двигательная память, каждая из них имеет собственные нервные механизмы консолидации. Более того, для того чтобы перевести относительно нестойкую кратковременную память в постоянную долговременную, необходима не реверберация как таковая, а деятельность лимбической системы (в случае декларативной памяти) или базальных ганглиев (в случае памяти процедурной).
Здесь мы завершаем рассмотрение теоретических построений Хебба. Надеемся, читатель согласится с нами в том, что Хебб указал психологической науке ранее не существовавшие или обойденные вниманием пути исследования. Он одним из первых начал поиск нейрофизиологических коррелятов психологических явлений, например научения. Во многом благодаря его усилиям нейропсихология стала сегодня столь популярна и проникла во многие области психологии за пределами направлений, в которых работал Хебб и его студенты. Было бы неуместно в этой книге подробно рассматривать все плодотворные направления, развивающиеся сегодня в рамках нейрофизиологической парадигмы. Далее мы хотим представить вам всего лишь несколько примеров подобных исследований. Первая тема, посвященная центрам подкрепления в мозге, косвенно связана с Хеб-бом, так как родилась из случайного открытия, сделанного в его лаборатории во время изучения ретикулярной активирующей системы (РАС). Следующий раздел, посвященный асимметрии мозга и вопросам лево- и правополушарности, не связан напрямую с теорией Хебба, хотя один из его студентов сделал большой вклад в данное направление нейрофизиологии. Завершающая тема, связанная с научением на клеточном уровне, вновь возвращает нас к фундаментальным представлениям Хебба о клеточных констелляциях.
Центры подкрепления в мозге
В главе, посвященной работам И. П. Павлова, мы отмечали, что открытие условного рефлекса произошло достаточно случайно. Интуитивная прозорливость — обнаружение одного явления во время поисков другого — в некоторых случаях ведет к открытию важнейших феноменов, итогом исследования которых становится прорыв в науке. Еще одним примером интуитивной прозорливости в науке является открытие центров подкрепления в мозге, сделанное Олдсом и Милнер в лаборатории Хебба в Университете Мак-Гилл (Olds & Milner, 1954). Олдс (Olds, 1955) так описывает это событие:
Осенью 1953 г. мы собирали информацию о ретикулярной активирующей системе. Для этого использовались электроды, на длительное время вживленные в мозг нормально ведущей себя крысы... По чистой случайности электрод был вживлен в область передней перегородки.
Полученный результат удивил всех. Когда крысу стимулировали в определенном месте на открытом пространстве, в некоторых случаях она покидала его, но затем возвращалась и начинала обнюхивать поверхность. Более сильная стимуляция в данном месте заставляла животное проводить на нем больше времени.
Позже мы обнаружили, что то же самое животное можно «привести» к любому месту лабиринта, давая небольшой электрический импульс после каждого поворота в правильном направлении. Все это очень напоминало детскую игру в «горячо-холодно». За каждой правильной реакцией следовала электрическая пульсация, которая, по-видимому, указывала животному, что оно находится на верном пути.
После этого случайного открытия центров подкрепления у крыс подобные структуры были найдены у кошек, собак, золотых рыбок, обезьян, дельфинов, пингвинов и людей. У людей стимуляция центра подкрепления вызывает эротические ощущения или просто удовольствие.
Олдс и Милнер (Olds & Milner, 1954) поначалу полагали, что в большинстве своем центры подкрепления находятся в области перегородки, тем не менее в дальнейшем выяснилось, что центры подкрепления рассеяны по всей лимбической системе мозга. Лимбическая система, участвующая в мотивированном и эмоциональном поведении, включает в себя нижние слои коры мозга, гиппокамп, миндалину, перегородку и отдельные структуры таламуса и гипоталамуса.
Данные области мозга принято называть центрами подкрепления, поскольку при их стимуляции животное, как правило, повторяет действия, которые оно производило до подачи стимула. Таким образом, животное с электродом, вживленным в центр подкрепления, можно научить проходить лабиринт или нажимать на рычаг в ящике Скиннера, всего лишь стимулируя данную область мозга слабыми электрическими импульсами при правильном выполнении действия.
Тем не менее было показано, что подкрепление посредством прямой стимуляции мозга имеет некоторые особенности и поэтому влияет на животное несколько иначе, чем традиционное подкрепление с помощью пищи и воды. Необычные характеристики подкрепления с помощью прямой стимуляции мозга кратко перечислены ниже.
1. Перед предварительным обучением не требуется депривация. В отличие от предварительного обучения с использованием в качестве подкрепления воды и пищи, при использовании прямой стимуляции мозга, как правило, не требуется соблюдения условий депривации перед началом обучения. Животное не должно
испытывать побуждения или потребности. Тем не менее из этого правила существуют исключения: обнаружены центры подкрепления, по всей видимости, зависящие от того, испытывает ли организм потребность.
2. Насыщение не наступает. При использовании в качестве подкрепления воды или пищи животное в конце концов насыщается. Потребность в пище или воде удовлетворяется, и животное перестает отвечать на стимулы. Тем не менее при прямой стимуляции мозга животное продолжает реагировать на стимуляцию с очень высокой частотой (так, например, существуют сведения о том, что крысы отвечали на стимуляцию нажатием на рычаг с частотой 7000 раз в час) вплоть до полного физического истощения.
3. Прямая стимуляция мозга отодвигает на второй план остальные потребности организма. Животные продолжают нажимать на рычаг, чтобы получить прямую стимуляцию мозга, даже если перед ними находится пища и они не ели в течение продолжительного времени. Точно так же животные, как правило, предпочитают выдержать более сильный удар током, чтобы получить подкрепление в виде прямой стимуляции мозга, нежели получить пищу даже в том случае, если они не ели в течение суток.
4. Угасание происходит стремительно. В отличие от постепенного угасания реакции, наблюдающегося при использовании в качестве подкрепления воды или пищи, при прекращении прямой стимуляции центров подкрепления мозга угасание происходит сразу же. Тем не менее, хотя угасание и происходит очень быстро, скорость воспроизведения реакции полностью восстанавливается, как только животное получает новое подкрепление.
5. Большинство режимов подкрепления не работают. Поскольку при прекращении стимуляции мозга угасание происходит очень быстро, любой режим подкрепления, подразумевающий задержку между реакцией и подкреплением, приводит к исчезновению реакции. Таким образом, при таком способе подкрепления могут использоваться только те режимы, в которых подкрепление происходит быстро и часто.
Сегодня исследования центров подкрепления сосредоточены на небольшом участке мозга, носящем название nucleus accumbens. При стимуляции эта область лимбической системы высвобождает химический нейротрансмиттер дофамин. Если стимулирующий электрод заставляет клетки nucleus accumbens высвобождать дофамин, стимуляция мозга через электрод будет восприниматься организмом как подкрепление. Если же стимулирующий электрод не вызывает высвобождения дофамина, эффект подкрепления не появится (Garris, Kilpatrick, Bunin, Michael, Walker & Wightman, 1999).

Берридж и Робинсон (Berridge & Robinson, 1995,1998), а также Каливас и Накамура (Kalivas & Nacamura, 1999) полагают, что nucleus accumbens ответственно за предвкушение удовольствия — тягу и стремление к нему, — а не ощущение удовольствия как такового. Столь большую популярность эта гипотеза приобрела в силу целого ряда причин. Во-первых, она помогает объяснить некоторые необычные характеристики подкрепления посредством прямой стимуляции мозга, например отсутствие насыщения и быстрое угасание реакции. Во-вторых, она предлагает новую интерпретацию пристрастия к наркотикам и поведения наркоманов. Хотя никотин, алкоголь, кокаин и героин очень сильно различаются по своему первичному химическому воздействию на мозг, они (и другие вызывающие привыкание вещества), по-видимому, одинаково стимулируют nucleus accumbens в лимбической системе (Leshner & Koob, 1999; Ranaldi, Pocock, Zereik & Wise, 1999). Как отмечают Берридж и Робинсон (Berridge & Robinson, 1995), даже после того как наркотические вещества теряют способность вызывать сильное ощущение удовольствия, они продолжают стимулировать тягу и стремление к удовольствию. Эти исследователи полагают, что именно долгосрочное воздействие наркотиков на nucleus accumbens в лимбической системе порождает навязчивые состояния у наркоманов, а также тягу к наркотикам, сохраняющуюся даже после того, как человек прекращает их употреблять.
Исследования расщепленного мозга
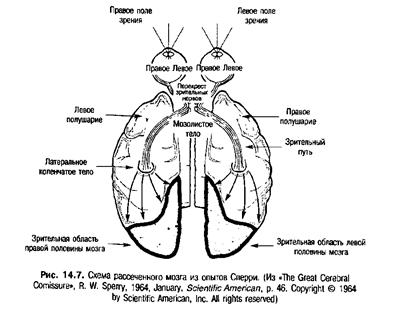
Мозолистое тело — это скопление волокон, соединяющих два полушария мозга. Многие годы функция мозолистого тела оставалась неясной, и лишь в 1960 г. было доказано, что эта мозговая структура принимает участие в передаче информации от одного полушария мозга к другому. В серии экспериментов Роджер Спер-ри (1913-1994) отметил, что существует два возможных пути для такого переноса информации: мозолистое тело и перекрест зрительных нервов (Sperry, 1961). Перекрест зрительных нервов — это участок зрительного нерва, через который информация, передающаяся от одного глаза, проецируется на противоположный этому глазу участок коры мозга. Сперри надевал повязку на один глаз здоровым кошкам, создавая таким образом условия зрительной дискриминации на один глаз. После дискриминативного обучения он исследовал перенос информации, перемещая повязку с одного глаза на другой. Как оказалось, животное справлялось с заданием столь же успешно, как и ранее. Другими словами, он обнаружил полноценный перенос информации между обоими глазами.
Далее Сперри задался целью найти механизм переноса информации от одного полушария мозга к другому. Сначала он провел эксперимент с рассечением зрительного перекреста перед обучением и после него. И вновь он столкнулся с полным переносом научения приобретенным навыкам от одного глаза к другому. Следующим этапом стало расщепление мозолистого тела после дискриминативного обучения. И вновь перенос научения был полным. Далее он рассек и зрительный перекрест, и мозолистое тело перед обучением, и обнаружил, что эти манипуляции препятствуют переносу научения от одного глаза к другому. Расщепление зрительного перекреста и мозолистого тела создало два независимых мозга, имеющих по одному глазу, но не способных обмениваться информацией. Схема рассеченного мозга, использовавшегося в опытах Сперри, представлена на рис. 14.7.
Кошки с расщепленным мозгом, обученные распознавать изображения с повязкой на одном глазу, не могли повторить свои действия, если повязку перемещали на другой глаз. Казалось, что оба полушария мозга обучались независимо друг от друга. Действительно, животное с повязкой на одном глазу можно было научить подходить к двери, на которой был нарисован крестик. Затем то же самое животное, но с повязкой на другом глазу, научалось подходить к соседней двери, на которой был нарисован круг. Таким образом, два мозга, находящиеся в одном черепе, приобретали противоположные навыки. Также можно было научить животное с повязкой на одном глазу подходить к стимулу (например, кругу), а передвинув повязку на другой глаз — избегать того же самого стимула.

В силу различных медицинских причин операция по расщеплению мозга проводилась и на людях. Помимо эффективного лечения различных мозговых отклонений, процедура рассечения мозга человека позволила получить ценные сведения о различиях обработки информации правым и левым полушариями мозга. Далее мы приступим к рассмотрению различий лево- и правополушарной деятельности мозга человека.
Левополушарное, правополушарное научение и обработка информации
Между правым и левым полушариями существуют небольшие анатомические различия, однако они не настолько велики, чтобы полушария работали совершенно по-разному. Более того, контроль над движениями тела и ощущениями разделен между полушариями мозга поровну, хотя и перекрестно. Левое полушарие контролирует правую сторону тела, правое полушарие — левую. Поскольку общее сходство полушарий очень велико, возникает искушение предположить, что полушария мозга научаются, воспринимают и обрабатывают информацию одинаково. Но так ли это на самом деле? Ответ на вопрос о существовании функциональной симметрии правого и левого полушарий мозга найти не так-то просто.
В 1836 г. Марк Дэкс сообщил о том, что потеря речи является следствием повреждения левого полушария мозга и не наблюдается при поражениях правого полушария. О наблюдениях Дэкса никто не вспомнил даже после того, как известный физиолог Поль Брока в 1861 г. пришел точно к таким же выводам, и сегодня мы называем одну из речевых зон левого полушария мозга речевым полем Брока. Сведения о том, что у большинства людей речевая зона находится в левом полушарии и отсутствует в правом, стали первым научным подтверждением функциональной асимметрии полушарий головного мозга.
Многие годы считалось, что правое полушарие находится в подчиненном положении по отношению к левому. Поэтому левое полушарие было названо доминантным или старшим. Однако в конце концов данные исследований привели к кардинальному изменению этой точки зрения. В 1962 г. серия экспериментов Ма-унткасла, Гешвинда, Каплана, Сперри и Газзанига показала, что правое полушарие не только равно по своей значимости левому, но по ряду неречевых функций превосходит его. Подтверждения функциональной асимметрии полушарий мозга были получены в ходе наблюдений за людьми с повреждениями мозга и пациентами, мозг которых был расщеплен по медицинским показаниям, а также в результате изобретательных экспериментов с участием здоровых людей с неповрежденным мозгом.
Так, например, оказалось, что люди с повреждениями правого полушария часто испытывают трудности с концентрацией внимания и восприятия. Они могут быть дезориентированы в знакомом им окружении, с трудом узнают знакомые им лица и объекты. Они чаще, чем пациенты с нарушениями левого полушария, демонстрируют синдром игнорирования — состояние, при котором больной не видит объекты в левом зрительном поле или же не может следить за левой половиной собственного тела. Пациенты с синдромом игнорирования бреют только правую сторону лица или едят пищу только с правой стороны тарелки. Когда пациентов с повреждениями правого полушария просят воспроизвести лежащее перед ними изображение, их рисунки чаще всего напоминают картинку, приведенную на рис. 14.8. Тот факт, что подобные трудности чаще возникают при повреждении правого полушария, стал еще одним доказательством функциональной асимметрии полушарий мозга.
Интерес к функционированию полушарий увеличился еще больше после появления пациентов, у которых по различным медицинским показаниям были рассечены подкорковые пути, соединявшие оба полушария мозга. У пациентов, страдающих эпилепсией, неподдающейся другим формам лечения, после расщепления мозолистого тела нездоровая электрическая активность затрагивает только одно полушарие, что уменьшает частоту и силу припадков. В то же время на примере таких пациентов можно изучать деятельность каждого из полушарий по отдельности. Сегодня разработан целый ряд техник представления сенсорной информации только одному полушарию мозга пациента.
Результаты исследований мозга таких пациентов очень сложны. Тем не менее некоторые исследователи полагают, что на их основе можно выделить правополу-шарные и левополушарные характеристики. Популярную, но несколько преждевременную интерпретацию полученных результатов дали Спрингер и Дойч (Springer & Deutch, 1985, p. 4-5): «Левое полушарие мозга участвует в основном в аналитических процессах, в частности — в воспроизведении и понимании речи. По всей видимости, оно обрабатывает информацию, поступающую извне, последовательно. Правое полушарие отвечает за определенные пространственные навыки и музыкальные способности; оно обрабатывает информацию одновременно и целиком».
Деятельность полушарий в здоровом мозге
Исследования с участием пациентов, мозг которых был рассечен по медицинским показаниям или поврежден, показали, что каждое полушарие может воспринимать, учиться, запоминать и чувствовать независимо от другого полушария. Деятельность обоих их полушарий здорового мозга можно исследовать с помощью метода, получившего название дихотомического прослушивания. Первым дихотомическое прослушивание применил Броадбент для исследования избирательного внимания. Но именно Дорин Кимура, студентка Хебба и Милнер, использовала его как безопасный и надежный метод исследования полушарной асимметрии у людей со здоровым мозгом (Kimura, 1961, 1964, 1967). Во время дихотомического прослушивания через стереонаушники в правое и левое ухо участника эксперимента одновременно подается разная информация, например различные слоги или цифры. Так, например, человек одновременно левым ухом слышит слог «ба», а правым «га». Известно, что информация, попадающая в левое ухо, поступает в основном в правое полушарие мозга, а информация, попадающая в правое ухо, проецируется в основном в левое полушарие. Таким образом, основной вопрос заключается в том, какой из двух одновременно услышанных стимулов участник эксперимента сможет воспроизвести правильно. В описанных выше условиях почти все правши и большинство левшей чаще правильно называют цифру или слог, услышанный правым ухом, чем левым. Таким образом, предположение о том, что левое полушарие мозга отвечает за обработку вербальной информации, подтверждается еще раз.

Некоторые исследователи полагают, что некорректно делать вывод о том, что левое полушарие специализируется на восприятии речи, исходя из результатов дихотомического прослушивания. Было бы правильнее утверждать, что оно специализируется на слуховом восприятии или внимании в целом. Тем не менее тот факт, что большинство нормальных правшей воспринимают мелодии (Kimura, 1964) и звуки окружающей среды, например лай собак или шум моторов (Curry, 1967), левым ухом (правым полушарием) лучше, чем правым, не поддерживает данный вывод.
Предположения
Исследование межполушарных различий привело к появлению на свет множества гипотез, касающихся роли асимметрии мозга в повседневной жизни. Некоторые из этих предположений приводят в своей работе Спрингер и Дойч (Springer & Deutch, 1985):
Говорилось о том, что эти различия отчетливо демонстрируют традиционный дуализм интеллекта и интуиции, науки и искусства, логичного и загадочного... Высказывалось предположение, что юристы и художники пользуются в своей работе различными полушариями мозга и что различия между полушариями проявляются в ходе деятельности, не связанной с их профессиональными занятиями. Некоторые авторы развивали эту мысль дальше и утверждали, что любого человека можно отнести к категории правополушарных или левополушарных людей в зависимости от того, какое полушарие по большей части руководит его поведением.
Боген (Bogen, 1977, р. 135) полагает, что дихотомичность, с которой зачастую описывают мир и мыслительный процесс, является отражением двух различных типов интеллекта, характерных для полушарий мозга. Согласно Богену, приведенные ниже противопоставления являются всего лишь проявлением различий в процессах обработки информации, характерных для правого и левого полушарий.
Левое полушарие Правое полушарие
Интеллект Интуиция
Конвергентный Дивергентный
Реалистичный Импульсивный
Интеллектуальный Чувственный
Дискретный Непрерывный
Управляемый Свободный
Рациональный Интуитивный
Исторический Вневременной
Аналитический Целостный
Последовательный Одновременный
Объективный Субъективный
Атомистический Макроскопический
Попытки отыскать дихотомию, подобную приведенной выше, а затем объяснить ее существование с точки зрения различий в обработке информации правым и левым полушариями мозга получили название дихотомании. Изучив работы, посвященные латеральное™, Битон (Beaton, 1985) делает вывод, что нельзя описывать деятельность полушарий мозга в терминах какой бы то ни было дихотомии.
Попытки обобщить некоторые «фундаментальные» различия полушарий мозга в терминах... дихотомии сопряжены с некоторыми проблемами. Во первых, практически все исследователи согласны, что асимметрия мозга не абсолютна, а выражена лишь в той или иной степени. Так, например, не было обнаружено полное отстутствие у одного полушария функций, в норме присущих другому полушарию. Даже если мы рассматриваем речь, в случае которой левосторонняя асимметрия наиболее очевидна, оказывается, что правое полушарие играет большую роль в понимании языка и может в определенных условиях проявлять свою компетенцию достаточно ярко...
Другие дихотомии выявляются еще менее надежно... Кроме того, нет причин, в силу которых мозг должен был бы развиваться столь удобно... Тем не менее ни одна из дихотомий мозговых функций по сути не является чем-то невероятным... Таким образом, было бы неправильно полагать, что связь между полушариями можно описать с помощью одного-единственного принципа.
Джерре Леви, долгое время занимавшаяся исследованием лево- и правополу-шарной деятельности, полагает, что хотя в особых условиях появляется возможность показать различия в работе полушарий, в нормальном здоровом мозге разделить эти функции невозможно. В своей статье «Правое полушарие, левое полушарие: правда и вымысел» (Right brain, left brain: facts and fiction) Леви (Levy, 1985) пишет:
Миф о существовании двух независимых частей мозга вырос из ошибочной предпосылки: поскольку полушария специализированы, каждое из них должно функционировать как независимый мозг. На самом же деле все как раз наоборот. Поскольку отдельные области мозга дифференцированы, они должны работать совместно. Действительно, интеграция приводит к появлению форм поведения и ментальных процессов, которые выражены сильнее и проявляются иначе, нежели вклад каждой отдельной зоны. Таким образом, поскольку неверна центральная предпосылка творцов мифов, то и выведенные из нее следствия опіибочньї... Мифами становятся неверные интерпретации или ожидания людей, а не наблюдения ученых. У нормальных людей в черепе находятся не половинки мозга и не два мозга, а один удивительным образом дифференцированный мозг, в котором каждое полушарие имеет собственные специализированные функции... Мозг един, и он создает единое умственное «я».
Газзанига и Ле До (Gazzaniga & LeDoux, 1978) в своем анализе менее великодушны. Проведя обширные исследования пациентов с расщепленным мозгом, они пришли к выводу, что популярная дихотомическая концепция мозга является следствием плохой продуманности экспериментов, в которых результаты определялись «влиянием реакции», а не реальными различиями полушарий. Вид реакции, ожидавшийся от участников исследования, влиял на экспериментальные данные сильнее, нежели процессы восприятия и познания, предшествовавшие ему. Из-за левополушарного речевого доминирования у большинства пациентов в заданиях, связанных с устной речью или письмом, левое полушарие работало лучше. Правое полушарие доминировало в тех случаях, когда пациентов просили отвечать с использованием движений рук в трехмерном пространстве, например при помощи рисования, строительства, прикосновений/ощущений и тому подобного. По мнению Газзанига и Ле До, при оценке специализации полушарий мозга с использованием заданий, сводящих к минимуму «влияние реакции», межполушарные различия практически исчезают. Газзанига и Ле До делают вывод, что хотя полушария руководят различными видами реакций, они воспринимают, научаются и обрабатывают информацию одинаково.
Исследования функциональных различий полушарий мозга продолжаются и сегодня (см. Hellige, 1993; Ornstein, 1997), однако они уже не связаны с предложенными ранее гипотезами о право- и левополушарных функциях. Интересно, что в недавних работах, проведенных на пациентах с расщепленным мозгом, была получена информация, которая не могла бы появиться на свет, если бы исследователи исходили из популярных дихотомий 1970-х и 1980-х гг. Так, например, Кронин-Колумб (Cronin-Columb, 1995) показывал правому и левому полушариям пациентов «картинки-цели» знакомых им объектов. После того как пациент видел «цель», ему показывали еще 20 картинок и просили отобрать связанные с картинкой-целью. Когда при выполнении задания пациенты пользовались правым полушарием, они, как правило, при отборе картинок использовали линейную классификацию. Так, первая отобранная картинка была теснее связана с мишенью, чем вторая, вторая — теснее, чем третья, и т. д. Если же использовалось левое полушарие, преобладала иная система отбора. Результаты недавних исследований также предполагают, что в правом полушарии лучше развита память, работающая с деталями изображений, воспринимаемых с помощью зрения (Metcalfe, Funnell & Gazzaniga, 1995). В левом полушарии сильнее представлены стратегии поиска с помощью визуального представления объекта (Kingstone, Enns, Mangun & Gazzaniga, 1995).
Без сомнения, исследования латеральности мозга привели к удивительным открытиям, и еще более удивительные открытия ждут нас впереди. Однако из-за того что эти открытия зачастую стимулируют воображение, важно сконцентрироваться на настоящих экспериментальных данных, позволяющих провести четкую грань между фактом и вымыслом.
Реальные клетки и констелляции клеток
Спустя годы с того момента, как Хебб впервые использовал понятия фракционирования, подкрепления, констелляций клеток и фазовых последовательностей, психологи были удивлены точностью его догадок о строении нервной системы.
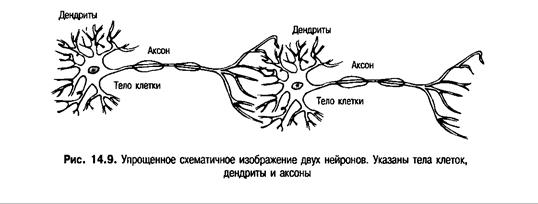
Предположения Хебба получили свое подтверждение после исследования процессов научения, происходящих между двумя нейронами. Каждый нейрон состоит из тела клетки, одного (или более) длинного отростка — аксона, передающего электрохимическую информацию от тела клетки, и множества ветвящихся отростков — дендритов, получающих электрохимическую информацию от аксонов других клеток. Упрощенное схематичное изображение пары клеток мозга представлено на рис. 14.9.
Клетки мозга млекопитающих находятся в водной среде, содержащей ионы калия, натрия и хлора, а также ионизированные белковые молекулы. Каждую клетку мозга можно рассматривать как хрупкий и чувствительный проводник подверженного изменениям электрохимического баланса. У млекопитающих нервные клетки участвуют в метаболических процессах, направленных прежде всего на поддержание баланса ионов калия и натрия внутри и вне клетки. Внутри нервной клетки преобладают ионы калия, вне нее — ионы натрия. Такое состояние «сбалансированного напряжения» принято называть потенциалом покоя клетки. Данный термин характеризует различие электрического заряда с внешней и внутренней сторон мембраны нервной клетки. У типичной, находящейся в покое нервной клетки млекопитающих внутренняя сторона клеточной мембраны по отношению к внешней заряжена отрицательно, и в среднем различие зарядов составляет около 70 мВ.
Если поляризация мембраны нарушается, разница электрических зарядов внутри и вне клетки начинает стремиться к нулю, и клетка может достичь порогового состояния, при котором она более не может поддерживать разницу ионов внутри и вне себя. В этот момент происходит небольшое изменение распределения ионов на мембране, связанное прежде всего с переносом и заменой ионов натрия на ионы калия. Это в свою очередь изменяет электрическое состояние клетки, у которой внутренняя сторона мембраны приобретает положительный заряд по отношению к внешней. Далее клетка расходует энергию на восстановление потенциала покоя. Весь процесс изменения ионного баланса и «перезарядки» мембраны принято называть потенциалом действия. Потенциал действия распространяется от тела клетки по всей длине аксона.
Окончание аксона реагирует на возникновение потенциала действия высвобождением химического вещества — нейротрансмиттера, например ацетилхолина или дофамина, в щель синапса (пространство между окончанием аксона одной и телом или дендритами другой клетки). Рецепторы дендритов или тел окружающих клеток отвечают на выброс нейротрансмиттера химическими реакциями, которые переводят эти клетки в пороговое состояние или, наоборот, препятствуют его возникновению.
Клетки мозга связаны с сотнями, а может быть, и тысячами других нервных клеток. Их собственная возбуждающая или ингибирующая активность является результатом постоянного суммирования химической информации, поступающей от окружающих клеток. Можно предположить, что на фундаментальном уровне научение связано с изменением взаимоотношений между двумя клетками. Именно на этом уровне было первоначально сосредоточено внимание Хебба. Научение заключается в изменении реакции нервной клетки, получившей сигнал, на ней-ротрансмиттер, высвобожденный пославшей сигнал нервной клеткой. В упрощенном варианте можно представить себе ранее не обучавшуюся нервную клетку, которая в ответ на нейротрансмиттер, выделяемый другой нервной клеткой, не генерирует собственный потенциал действия. Научением можно назвать состояние, при котором получающая сигнал нервная клетка начинает надежно и предсказуемо генерировать потенциал действия в ответ на активность посылающей сигнал клетки. Хотя Хебб предполагал, что активность одной клетки при соприкосновении с другой может изменять взаимоотношения между ними, он мог лишь предполагать, какие процессы происходят при этом. Тем не менее исследования последних лет выявили механизмы, весьма похожие на предположения, высказанные Хеббом.
Научение у моллюсков p. Aplysia
Главным препятствием на пути понимания возможных механизмов научения, подкрепления и фракционирования является большое количество нейронов, участвующих в реализации самых простых форм поведения у млекопитающих. Кэн-дел с соавторами (Castellucci & Kandel, 1974; Dale, Schacher & Kandel, 1988; Kandel & Schwartz, 1982; Kupfermann, Castellucci, Pinsker & Kandel, 1970) сумели разрешить эту проблему, проведя исследования на морских моллюсках p. Aplysia. Нервная система этих животных устроена относительно просто, и в то же время для них характерны формы поведения, при которых можно предположить существование клеточных констелляций. На спинной части тела этого морского моллюска располагаются три внешних органа: жабры, мантия и сифон. Все эти структуры рефлекторно сжимаются, если задеть мантию или сифон животного.
Если один из этих рефлекторно сокращающихся органов многократно слабо стимулировать, у животного развивается реакция габитуации — в ответ на внешнюю стимуляцию органы постепенно перестают сокращаться. Таким образом, нервный путь, изначально активированный поступавшим извне стимулом, «вычитается» из более сложного паттерна нервной деятельности. Этот процесс во многом соответствует представлениям Хебба о фракционировании, однако каков механизм габитуации?

Исследования Кэндела (Castellucci & Kandel, 1974) показали, что критическим событием для развития габитуации является уменьшение выделения нейротрансмиттера (нейротрансмиттеров) из сенсорного нейрона, который передает информацию о слабой стимуляции, к двигательному нейрону, который запускает рефлекторное сокращение внешних органов. Каким образом и почему сенсорный нейрон научается игнорировать слабую, постоянно повторяющуюся стимуляцию, неизвестно. Тем не менее реакция сокращения внешних органов легко восстанавливается, и этот факт свидетельствует о том, что габитуация представляет собой нечто иное, нежели простое утомление или истощение запасов нейротрансмиттера (нейротрансмиттеров) (Kupfermann, Castellucci, Pinsker & Kandel, 1970).
Процесс реактивации, который также принято назвать сенсибилизацией, наблюдается, например, в тех случаях, когда животное получает электрический разряд в области хвоста. После удара током повторная слабая стимуляция вновь вызывает рефлекторное сокращение. Сенсибилизация более сложна, чем габитуация. Она связана с работой еще одного нейрона, называемого промежуточным нейроном, который активизируется, например, при электрошоке. В этом случае дополнительный промежуточный нейрон стимулирует сенсорный нейрон, заставляя его высвобождать дополнительную порцию нейротрансмиттера на дендриты двигательного нейрона, контролирующего сокращение органов (Geary, Hammer & Byrne, 1989; Dale, Schacher & Kandel, 1988). Таким образом, сенсибилизация, по-видимому, связана с созданием простой констелляции клеток, состоящей из трех элементов, и может служить иллюстрацией предложенной Хеббом модели феномена подкрепления. Кроме того, как читатель, наверное, уже успел догадаться, в ходе дальнейших исследований Кэнделу удалось показать, что процессы с участием промежуточных нейронов (например, процесс сенсибилизации) лежат в основе процесса классического обусловливания (Kandel & Schwartz, 1982).
Долгосрочное потенцирование
Исследования Кэндела отчасти позволили ответить на вопрос, как изменяются коммуникативные паттерны между клетками. Дополнительные механизмы были раскрыты при изучении явления, получившего название долгосрочного потенцирования (ДП) (Bliss & Lomo, 1973; Lomo, 1966). Если одну из частей гиппокампа (структуры, принимающей участие в консолидации памяти) стимулировать одиночными слабыми электрическими импульсами, то по характеру распределения нервно-электрической активности, вызванной этим начальным импульсом, можно судить о силе связей этой области с другими частями гиппокампа. В частности, если стимулировать клетки гиппокампа, находящиеся в области перфорантного пути, можно зарегистрировать активность в области зубчатой извилины. Если после слабого импульса сразу же следует более сильная высокочастотная электрическая стимуляция, связь между клетками перфорантного пути и зубчатой извилины очень существенно изменяется. Вначале, при небольшой силе стимула, сигнал распространяется медленно, но после высокочастотной стимуляции слабое электрическое воздействие на область перфорантного пути вызывает значительно более сильную активность зубчатой извилины и прилегающих к ней структур. Таким образом, более сильная высокочастотная стимуляция усиливает действие первоначальной более слабой стимуляции, и эффект усиления может сохраняться до нескольких месяцев (рис. 14.10).
В некоторых частях гиппокампа долгосрочное потенцирование развивается по описанной выше схеме. В других областях той же части мозга ДП не возникает до тех пор, пока слабая и более сильная высокочастотная стимуляция не будут наблюдаться одновременно. Кэндел (Kandel, 1991) предположил, что первая форма ДП является отражением нервной основы неассоциативного научения (габитуации и сенсибилизации), а вторая — ассоциативного научения. Важно отметить, что ассоциативное научение влечет за собой события, описанные в нейрофизиологическом постулате Хебба. Клетка — источник стимула, оказывающая слабое влияние на клетку-получатель, сохраняет активность в тот момент, когда клетка-получатель стимулируется другой, более сильной клеткой — источником стимула. Одновременная активность слабой клетки-источника и клетки-получателя изменяет электрохимическую чувствительность и взаимосвязь между двумя клетками. Нейрофизиологи называют синапс между связанными таким образом нейронами синапсом Хебба.
В последнее время значительное количество работ было посвящено поиску механизмов ассоциативного ДП в синапсах Хебба. Так, например, было показано, что подобный эффект вызывает нейротрансмиттер глютамат, к которому, однако, существует по меньшей мере два типа глютаматных рецепторов на дендритах (Cotman, Monaghan & Ganong, 1988; Nakanishi, 1992). Один из них, NMDA глюта-матный рецептор (названный в соответствии с химической процедурой, использующейся для отделения его от других глютаматных рецепторов) невозможно активировать, пока рядом лежащие не-NMDA рецепторы той же клетки-получателя не будут активированы глютаматом. Если рецепторы обоих типов одновременно получают стимул, NMDA рецепторы активируются и позволяют ионам кальция и натрия попасть внутрь клетки. Исследователи полагают, что ионы кальция запускают серию событий, которые некоторым образом увеличивают чувствительность не-NDMA рецепторов (Bading, Ginty & Greenberg, 1993; Baudry & Lynch, 1993; Lynch & Baudry, 1984, 1991; Schuman & Madison, 1991).
Поначалу предполагалось, что ДП будет наблюдаться только в том случае, если потенцирующий стимул имеет высокую частоту, около ста импульсов в секунду. Позднее, исходя из того что в мозге очень низка вероятность возникновения высокочастотной стимуляции, ДП стали рассматривать лишь как лабораторное явление. Интересно, что при помещении крысы в новую сложную среду, во время реализации исследовательского поведения (предположительно связанного с научением в новой среде), можно зарегистрировать генерируемые изнутри низкочастотные серии импульсов (называемых тета-ритмами). Они возникают в области перфорантного пути и распространяются на клетки, лежащие в области зубчатой извилины, проходя, таким образом, тот же путь, что и в опытах с искусственно вызванным ДП (Vanderwolf, Kramis, Gillespie & Bland, 1975). Результаты исследований свидетельствуют о том, что искусственно вызванный тета-ритм вызывает ДП столь же эффективно, как и сильная высокочастотная стимуляция, использованная в классическом ДП эксперименте (Diamond, Dunwiddie & Rose, 1988; Staubli & Lynch, 1987). На основании этих открытий ДП, вызванное внутренним потенцирующим импульсом, сегодня рассматривается как возможный способ некоторых форм естественного научения (Escobar, Alcocer & Chao, 1998; Stanton, 1996), хотя и эта точка зрения подвергается сомнению (см., например, Hoelscher, 1997).
Долгосрочное угнетение
Научение связано с вовлечением новых констелляций клеток и фазовых последовательностей, которые необходимы для успешного когнитивного и моторного поведения. В то же время научение приводит к разрушению фазовых последовательностей, которые в настоящее время не нужны или активно препятствуют осуществлению эффективных форм поведения. ДП представляет собой механизм стимуляции и вовлечения в констелляцию ранее не связанных с ней нейронов. Явление долгосрочного угнетения (ДУ) можно рассматривать как возможный способ дезактивации нервных клеток и их выхода из констелляции. При ДУ, если две клетки-источника стимулируют одну клетку-получатель, она теряет способность отвечать на поступающие к ней стимулы (Kerr & Abraham, 1995). Явление ДУ было обнаружено в мозжечке, некоторых частях гиппокампа (Akhondzadeh & Stone, 1996; Doyere, Errington, Laroche & Bliss, 1996) и коры головного мозга (Kirk-wood, Rozas, Kirkwood, Perez & Bear, 1999). На сегодняшний день роль NMDA-pe-цепторов в ДУ еще не выяснена; весьма возможно, что это явление связано с работой иного, нежели глютамат, нейротрансмиттера (Kirkwood, Rozas, Kirkwood, Perez & Bear, 1999).
Таким образом, существует целый ряд способов, с помощью которых может осуществляться простое научение двух нейронов или небольшой группы нервных клеток. Мы также рассмотрели несколько вариантов детализации клеточных констелляций Хебба. Хотя некоторые ранние замечания Хебба о механизмах работы клеточных констелляций оказались неверны, центральное положение его теории, говорящее о том, что клетки развиваются и работают группами, по-видимому, верно. Тем не менее мозг человека настолько сложен, что реальная связь между деятельностью групп клеток мозга и сложными формами научения, например запоминанием имен, развитием речи, социальных или когнитивных правил, еще долгие годы будет оставаться предметом многих исследований.
Новый коннекционизм. Искусственные клетки и констелляции клеток
Хебб и представить себе не мог, что его идеи найдут применение в абстрактном мире компьютерного моделирования. Тем не менее новейший подход к изучению способов научения нервной системы вообще не связан с исследованием настоящих нервных клеток. Компьютерные модели деятельности клеток мозга используются для исследования научения, памяти, забывания и других процессов, происходящих в головном мозге. Наиболее влиятельные исследователи, работающие в этой области, — Дэвид Румельхарт и Джеймс Мак-Клелланд, — исходя из предположения о том, что мозг может обрабатывать информацию одновременно или параллельно, назвали этот подход параллельно распределенной обработкой информации (ПРОИ) (McClelland & Rumelhart, 1988; Rumelhart, McClelland & PDP Research Group, 1986). У данной научной области пока еще нет общепринятого названия, но чаще всего ее называют новым коннекционизмом, а использующиеся в таких исследованиях модели — нейронными сетями (Bechtel & Ab-rahamsen, 1991).
В компьютерных моделях прежде всего необходимо определить группу компьютерных нейронов, а также их потенциальные взаимосвязи и отношения. Далее к этим искусственно созданным нейронам применяется ряд упрощенных допущений, выведенных из того, что на сегодняшний день известно исследователям о реальных нервных клетках. Кроме того, изменения отдельных компьютерных нейронов и их связей регулируются простыми логическими правилами научения. И наконец, искусственную нервную систему «обучают», а затем изучают, определяя, каким образом она изменяется в результате полученного опыта.
Для того чтобы продемонстрировать идею нового коннекционизма, воспользуемся примером простой нейронной сети, названной ассоциативным паттерном (Bechtel & Abrahamsen, 1991, Hinton & Anderson, 1981; Rumelhart, McClelland & PDP Research Group, 1986). Тем не менее следует помнить, что на сегодняшний день с помощью нервных сетей удалось смоделировать гораздо более сложные явления.
Во-первых, рассмотрим простой набор элементов, представленных на рис. 14.11. Данная нейронная сеть состоит всего из четырех элементов: двух входных и двух выходных нейронов. Их можно рассматривать как сенсорные и моторные нейроны соответственно. Пунктирными линиями обозначены возможные нервные связи между элементами.
Ввод из внешней среды (или от программиста) активирует входные нейроны. Выходные нейроны активируются в зависимости от того 1) какова сила соединения входящих элементов и 2) какое количество входящих элементов связано с ними. Данное правило активации выходящих элементов отражает свойство сум-мации, характерное для действующих нейронов. Свойство суммации заключается в том, что реально существующие нейроны складывают сигналы, поступающие к ним от окружающих клеток. Сумма сигналов определяет уровень активности нейрона. Это правило можно записать следующим образом:
A 0= S ( woi ) Ai .
Данное уравнение означает, что выходящая активация элемента (А()) представляет собой сумму его входящих активаций (Aj), умноженную на силы их связности (woi). Здесь мы можем предположить, что данная гипотетическая система ранее не участвовала в процессах научения и все woi равны нулю, то есть сенсорный ввод не влияет на моторный вывод.
Предположим, что мы хотим научить нашу нейронную сеть различать сосны и ели. В результате наша система должна говорить «сосна», если входящий сигнал — изображение сосны, и «ель», если входящий сигнал — изображение ели. Важно помнить, что в реальной нервной системе надписанные этикетки или ярлычки не связаны с сенсорным вводом или моторным выводом. В упрощенном варианте сенсорный ввод в виде изображения сосны и моторный вывод в форме слова «сосна» представляют собой не более чем примеры активизации и торможения в нервной системе. Именно в этом смысле мы предлагаем «сосну» и «ель» нашей гипотетической нейронной сети. Произвольным образом присвоим «сосне» сенсорный код (+1,-1), обозначающий, что для первого сенсорного элемента характерна возбуждающая активность +1, а для второго — тормозящая активность -1. «Ели», напротив, присвоим код (-1,+ 1), означающий тормозящую активность -1 в случае первого сенсорного элемента и возбуждающую активность + 1 в случае второго.
Мы знаем, что (+1,-1) — это «сосна» и (-1,+1) — «ель», и можем надежно относить к определенной категории каждый из этих двух видов деревьев. Проблема заключается в том, чтобы научить компьютерную нейронную сеть правильно относить каждое изображение к определенной категории и, как следствие, демонстрировать различие этих двух типов входящей информации. Это значит, что при вводе (+1,-1) в качестве сенсорной информации мы хотим, чтобы первый моторный нейрон выводил нам +1, а второй -1. Соответственно при вводе «ель» моторные нейроны должны вести себя противоположным образом. Для того чтобы закрепить этот эффект, мы должны обучить систему. Нам нужно развить связи между сенсорными и моторными элементами таким образом, чтобы могли сформироваться желаемые взаимоотношения ввода-вывода. Отметим, что клетки не научаются возбуждаться (+1) или затормаживать (-1). Предполагается, что, как и в случае реально существующих нейронов, способность к возбуждению и торможению является простыми наследственными свойствами клеток. Научение происходит за счет связей между клетками и зависит от типа и силы связей, сформированных в нейронных сетях.
В этом месте наших рассуждений мы должны вывести правило научения — логическое, но произвольное правило, которому наша компьютерная система будет следовать, изменяя связи между клетками. Простейшее из подобных правил названо правилом Хебба. Оно представлено математическим выражением, призванным отразить предположение Хебба о том, что связь между двумя одновременно активными клетками будет усиливаться или становиться более эффективной. Запишем это выражение следующим образом:
Awoi = Irate ( Aj )( A 0),
где A woi — изменение силы или значение связи между вводом и выводом; Irate — константа, которая отражает темп научения; Aj — значение уровня активации входящего элемента; А0 — значение уровня активации выходящего элемента. В нашем простом примере значение уровней активации может быть равно -1 или +1.
Правило гласит, что если два элемента активируются в одном направлении (оба имеют значение +1 или -1), то результат такой взаимной активности будет положительным, и, следовательно, более положительным будет значение связи. Если же элементы одновременно активизируются в различных направлениях (один имеет значение +1, а другой —1), результат будет отрицательным, и более отрицательным будет значение связи. Таким образом, легко проследить, как изменятся связи между элементами в наших «сосновых» и «еловых» примерах.
Начнем с того, что все веса или связи приравняем к нулю (0), а темп научения (Irate) определим как 1/п, где п - количество входных элементов. Другими слова ми, темп научения в нашем случае будет равен Уг. (Это произвольное определение идеально для нашего простого примера и гарантирует, что достаточно будет одной попытки научения.)
Начать можно с обучения сети внешнему сигналу «сосна». Компьютерная программа устанавливает уровни активации входных элементов равными +1 в первом и -1 во втором случае, а уровням активации выходных клеток будут приданы желаемые значения +1 в случае первого и -1 в случае второго нейронов. Приведенная ниже матрица демонстрирует состояние входных элементов, выходных элементов и веса (силы связей) в начале обучения.
Для изменения силы связей от начальных нулевых значений воспользуемся правилом Хебба. Так, например, мы можем изменить связь между входящим элементом 1 и выходящим элементом 1 с помощью простой подстановки в формуле научения:
Aw — величина изменений (от нуля), которые возникли вследствие активации первого входящего и первого выходящего нейронов; Aj = активация входящего элемента 1 = +1; А0 = активация выходящего элемента 1 = +1; Irate = Уі.
Следовательно, A w = V4 (+1) (-1) = Уг или 0,50.
Другие значения будут изменяться соответственно, и после предварительного обучения нейронной сети они будут иметь следующие значения.
Можно протестировать эту нейронную сеть и убедиться в том, было ли научение эффективным, введя входящие данные и позволив сети самой генерировать выходящие данные в соответствии с правилом выходящей активации (сумма-ции), приведенным выше. Если ввести «сосна» (+1,-1), выходящая клетка 1 прибавляет (0,50)(+1) + (-0,50)(-1) и получает +1; выходящая клетка 2 прибавляет (-0,50)(+1) + (0,50)(-1) и получает -1. В результате сеть выдает правильный ответ (+1,-1). Этот результат надежно повторяется, поскольку веса или силы связей были получены исходя из входящих данных и выходящих значений «сосна». Значительно более удивляет результат, получающийся, когда той же системе дают входящие данные «ель» (-1,+ 1). Попробуйте и убедитесь в этом сами.
Системы обратной связи
На сегодняшний день исследования нейронных сетей достигли уровня сложности, выходящего далеко за рамки этой книги. Простой пример противопоставления «сосны» и «ели» — всего лишь введение в область исследований, уже сегодня подающую большие надежды. Представьте себе нейронную сеть, скажем, из десяти входящих и десяти выходящих элементов, а также некоторого количества промежуточных нейронов. Наше простое объяснение и пример лишь бегло познакомит вас с явлениями, которые может имитировать подобная нейронная сеть. И в то же время общий принцип суммации во всех сетях во многом остается одним и тем же. Тем не менее простое правило научения Хебба в более сложных сетях обычно заменяют одной из форм правша дельта (McCelland & Rumelhart, 1988; Rumelhart, McCelland & PDP Research Group, 1986), называемого еще правилом обратной связи. Фундаментальное правило дельта выглядит следующим образом:
A woi = Irate (d0 - А0) (Aj).
Оно очень похоже на правило Хебба, за исключением усложнения члена, связанного с определением активации на выходе. В данном правиле (d0) представляет желаемое значение выходного элемента, а (AJ — его реальное значение. Система программируется таким образом, чтобы сила связей между элементами изменялась в направлении уменьшения различий между желаемым и реальным значением выходного элемента. Таким образом, это правило является самокорректирующимся правилом научения, которое изменяет силы связей до тех пор, пока значение выходного элемента не достигнет желаемой величины. Если (dQ - А0) приобретает нулевое значение, значения более не меняются и научение считается законченным.
Особое внимание исследователей нейронных сетей привлекла система обратной связи NETtalk (Sejnowski & Rosenberg, 1987). Эта система состоит из сканирующего компьютерного устройства с семью окнами, каждое из которых может сканировать одну печатную букву английского алфавита. Каждое сканирующее окно соединено с 29 входными элементами сети. Выходных элементов 26, и каждый из них соответствует отдельной фонеме английского языка. Каждый из выходных элементов в свою очередь запрограммирован на воспроизведение определенной фонемы через голосовой синтезатор. Между входящими и выходящими элементами находится восемьдесят встроенных или скрытых элементов. Каждый входящий элемент соединен с каждым скрытым элементом, а каждый скрытый элемент соединен с каждым выходящим элементом. Таким образом, в NETtalk существует 7 х 29 х 80 х 26 или 18320 взвешенных связей. Вначале значения определяются случайным образом. Когда система сканирует напечатанное слово и в нее поступает входящая информация, она поначалу выдает случайный набор звуков. Для подгонки значений используется правило обратной связи, и постепенно выходящая информация системы все более приближается к желаемому результату. В конце концов NETtalk научается читать слова вслух. Интересно, что после начального обучения, включающего одну тысячу слов, NETtalk уже оказывается способна правильно читать слова, не входившие в первоначальный набор, хотя и делает ошибки, обычные для людей, недавно научившихся читать. Так, например, если в наборе слов для первоначального обучения было слово rough, система прочтет новое слово tough правильно, но ошибется при чтении слова dough. Кларк (Clark, 1990) описывает, как NETtalk училась читать вслух.
Поначалу значения скрытых элементов и связи (между выбранными параметрами) были распределены в нейронной сети случайным образом, т. е. система не имела «представления» о каких бы то ни было правилах перехода от текста к фонемам. Ей нужно было научиться путем многократного повторения примеров освоиться в этой когнитивной области знаний, особенно каверзной из-за исключений, подсистем и контекстной зависимости перевода символов текста в фонемы. Научение происходило стандартным образом, то есть в соответствии с правилом обратной связи. Системе давали входные данные, автоматически, с помощью компьютеризированного «наставника» проверяли результат и сообщали, какой результат (т. е. какой фонемный код) она должна была произвести на выходе. Правило научения заставляло систему ежеминутно подгонять значения скрытых элементов таким образом, чтобы они как можно сильнее совпадали с правильным результатом. Процедура повторялась много тысяч раз. Удивительным образом система медленно училась воспроизводить вслух английский текст: начав с лепета, она стала произносить отчасти узнаваемые слова и наконец достигла стабильного впечатляющего результата.
Было бы неправильно думать, что исследования нейронных сетей — это поле деятельности одного лишь «компьютерного братства» — высококвалифицированных поограммистов, пишущих сложные программы, которые воспроизводят уже известные и понятные явления. Новый коннекционизм начался с относительно простых допущений: с правила научения, предложенного Хеббом, и предположения Лэшли о том, что память распределена между многими нервными клетками, а не сосредоточена в одном или двух нейронах. Исходя именно из этих посылок конструировались нейронные сети, воспроизводящие и простые задачи распознания изображений, и сложные процессы освоения языка и восстановления функций после повреждений мозга. Современные исследователи используют нейронные сети как средство познания того, как человеческий мозг определяет высоту и частоту различных звуков (May, Tiitinen, Ilmoniemi, Nyman, Taylor & Naeaetaenen, 1999), как мы научаемся мысленно представлять концепции числа и счета (Anderson, 1998) и каким образом болезнь Паркинсона (Mahurin, 1998) или болезнь Альцгеймера (Tippett & Farah, 1998) воздействует на мозг. Можно ожидать в ближайшие годы появления новых работ, посвященных нейронным сетям.
Краткое содержание главы
После нелегких лет учебы в университете Далхаузи Хебб поступил в магистрату РУ университета Мак-Гилл, где обучение было основано на традициях школы Павлова. Познакомившись с гештальт-психологией и работами Лэшли, посвященными физиологии мозга, Хебб разочаровался в учении Павлова. Работая в Чикагском университете под руководством Лэшли, Хебб пришел к убеждению, что головной мозг функционирует не как сложный синтезатор, а скорее как целостная структура, состоящая из взаимосвязанных частей. Такая гештальт-кон-цепция мозга подтвердилась в дальнейшем, когда Хебб, работая с Пенфилдом, наблюдал практически полное сохранение интеллекта у пациентов, перенесших удаление значительных областей мозга.
Основной заслугой Хебба можно считать появление в психологической науке понятий констелляции клеток и фазовой последовательности. Констелляция клеток — это группа клеток мозга, связанная с определенным объектом окружающей среды. Если эта группа клеток активизируется в отсутствие связанного с ней объекта, у человека возникает идея этого объекта. Фазовая последовательность — это серия связанных между собой констелляций клеток. Если в окружающей среде несколько событий, как правило, происходят вместе, то на нервном уровне они оказываются представленными в виде фазовой последовательности. Стимуляция фазовой последовательности приводит к возникновению потока взаимосвязанных идей. Хебб предполагал существование двух типов научения. Во-первых, это научение в начале жизни, когда наблюдается медленный рост констелляций клеток и фазовых последовательностей. Во-вторых, это научение с помощью инсайта, которое можно наблюдать у взрослых людей. Научение взрослых особей в большей степени связано с перестройкой констелляций клеток и фазовых последовательностей и в меньшей — с их развитием.
Теория активации подразумевает, что стимулы окружающей среды имеют две функции: 1) ключевую функцию, связанную с передачей информации об окружающей среде, и 2) активирующую функцию, связанную с возбуждением ретикулярной активирующей системы. Для того чтобы интеллектуальная деятельность была оптимальной, активация не должна быть слишком сильной или слишком слабой. Если для оптимального выполнения определенного задания уровень активации слишком низок, любой стимул, способный его увеличить, будет воспринят организмом как подкрепление. Если же уровень активации слишком высок, подкреплением будет любой стимул, способный его понизить.
Сенсорная депривация негативно сказывается на когнитивной деятельности организма, поскольку влечет за собой изменение связей между нервными контурами и событиями окружающей среды. Результаты исследований, посвященных сенсорной депривации, свидетельствуют о том, что организмы нуждаются в нормальном уровне стимуляции столь же сильно, как в пище, воде или кислороде. Согласно данным экспериментов, животные, выросшие в богатой сенсорными стимулами среде, впоследствии научаются лучше, чем животные, выросшие в аскетичной среде. Хебб предположил, что у животных, воспитывавшихся в обогащенной среде, формируются более сложные нервные контуры, которые в дальнейшем могут быть использованы при научении.
Исследуя феномен страха, Хебб обнаружил, что шимпанзе не испытывали страха при виде совершенно знакомых и совершенно незнакомых объектов. Страх и ужас у них вызвали знакомые объекты, представленные им в незнакомом виде. Хебб предположил, что знакомые объекты активировали связанные с ними нервные контуры, но следовавшие за предъявлением объектов события не поддерживали активность нервных клеток. Возникал конфликт, который порождал страх. Эта теория позволила также объяснить, почему сенсорная депривация сказывалась столь негативно на психике и самочувствии людей, участвовавших в эксперименте.
Хебб полагал, что существует два типа памяти — кратковременная и долговременная память. В кратковременной памяти информация сохраняется менее минуты. Этот вид памяти связан с реверберирующей нервной деятельностью, вызванной событием окружающей среды. Если определенное явление повторяется достаточно часто, оно переводится в долговременную память. Процесс перевода информации из кратковременной в долговременную память носит название консолидации. Если в момент консолидации происходит травма, кратковременная память не переводится в долговременную память. Результаты исследований свидетельствуют о том, что полная консолидация информации занимает около часа. Новейшие исследования показали, что для различных типов долгосрочной памяти характерны различные механизмы консолидации.
Теоретические работы Хебба дали начало исследованиям различных нейрофизиологических явлений. Олдс и Милнер, проводившие в лаборатории Хебба исследования систем активации, случайно открыли центры подкрепления в мозге. Сперри обнаружил, что при рассечении зрительного перекреста и мозолистого тела в одном черепе появляются два независимых мозга. Этим независимым мозговым структурам можно привить противоположные привычки; один мозг может отдыхать, когда второй остается активным. Дальнейшие исследования мозга показали, что хотя правое и левое полушария мозга анатомически сходны, каждое из них доминирует в отношении определенных функций. Кэндел и другие исследователи, изучая отдельные нейроны и небольшие группы нервных клеток, открыли механизмы, с помощью которых могут возникать настоящие констелляции клеток и фазовые последовательности. Были исследованы различия механизмов ассоциативного и неассоциативного научения. Специалисты в области компьютерных технологий использовали идеи Хебба для построения рабочих компьютерных моделей, воспроизводящих самые разные явления — научение языку, восстановление мозга после повреждений, заболевания, связанные с изменением деятельности мозга.
Следует признать, что за исключением работ Хебба, исследования, проводящиеся в рамках нейрофизиологической парадигмы, часто распылены и слабо связаны между собой. Тем не менее они существенно обогащают наши знания о процессах научения. В главе 3 мы уже говорили о том, что наше понимание процессов научения будет более полным, если мы рассмотрим это явление с различных точек зрения. Нейрофизиологическая парадигма как раз дает нам возможность взглянуть на процесс научения еще под одним углом зрения.
Оценка теории Хебба. Вклад в развитие науки
Основная заслуга Хебба в том, что он сумел показать, что сложные когнитивные процессы можно изучать, используя в качестве фундаментальных инструментов исследования отдельные нейроны и синапсы. В этом отношении Хебб четко отделил свои построения от других теорий, основывавшихся на абстрактных С-Р связях. В наше время студенты, изучающие психологию и нейрофизиологию, воспринимают фундаментальную связь между синаптической активностью и явлениями высшей нервной деятельности как данность. Тем не менее именно Хебб первым связал между собой эти явления и построил простые модели возникновения высших нервных процессов на основе простых синаптических событий. С момента возникновения теории Хебба прошло более 50 лет, а она все еще оказывает эвристическое влияние на нейрофизиологию и компьютерные исследования нейронных сетей.
Согласно Хеббу, фундаментальный принцип научения связан лишь с повторением и ассоциацией идей и основывается на понимании того, какую роль в данном процессе могут играть нейроны. Хотя сам Хебб стремился видоизменить основной принцип научения и включить в него научение через подкрепление, сегодня становится ясно, что его теория не нуждается в подобном дополнении. Она может объяснить и перцептивное научение, и формирование условных рефлексов на основе подкрепления, и высшие эмоциональные и когнитивные процессы. В этом смысле теория Хебба столь же элегантна, как и построения Газри. Ее научная привлекательность заключена в ее простоте, а не в ее прямой связи с биологическими и нейрофизиологическими механизмами.
Как и Толмен, Хебб видел различия между мотивацией и научением и учитывал трудности, связанные с их разделением при наблюдении за поведением. Созданные Хеббом теория активации и концепция оптимальной активации не решили эту проблему, но указали новые пути ее концептуализации. Теоретические построения Хебба позволили ответить на вопросы, возникшие вместе с появлением гипотезы Халла, связанной с ослаблением потребности, и объяснили, почему в некоторых случаях мы стремимся к ослаблению потребности, а в других — наоборот, к ее усилению. Таким образом, работы Хебба, посвященные природе активации, эффектам сенсорной депривации, подкреплению и природе страха, оказали заметное влияние как на изучение мотивации, так и на исследования научения.
Критика
Несомненно, Хебб не был первым среди тех ученых, которые связывали научение с мозгом. В некотором смысле его идеи формирования связей между смежными активными областями мозга не слишком отличались от представлений Павлова. Точно так же до него исследователи уже использовали свои представления о деятельности мозга для объяснения высших когнитивных процессов. Можно сказать, что Хебб поменял уровень анализа и перешел от рассмотрения крупных областей мозга к исследованию небольших групп нейронов, придерживаясь в то же время основных принципов школы Павлова.
Еще один повод для критики дало явное нежелание Хебба пересмотреть некоторые положения его теории в свете более поздних открытий нейрофизиологии. Тем не менее следует признать, что Хебб рассматривал свою систему скорее как спекулятивную модель теории, нежели как законченные, формальные теоретические построения. Весьма может быть, что Хебб воспринимал открытие многочисленных нейротрансмиттеров, физиологических основ подкрепления и нервных структур, участвующих в процессе консолидации, как интересные, но не связанные с его основной моделью находки или же как промежуточные этапы развития науки о мозге. С другой стороны, следует упомянуть об одном достаточно простом вопросе, который, однако, мог иметь большое значение для теоретических построений Хебба.
Как указывает Квинлэн (Quinlan, 1991), Хебб основывал своей нейрофизиологический постулат исключительно на явлении возбуждения. Тем не менее по мере развития наших представлений о нервной системе стало понятно, что большинство нервных связей (и большинство нейротрансмиттеров) оказывают тормозящее действие. Это означает, что чаще всего в нервной системе наблюдается ингибирование: одна нервная клетка ограничивает возбуждение другой клетки. Без сомнения, открытие этого фундаментального явления скажется на науке о мозге, несмотря на то что Хебб проигнорировал его в своем нейрофизиологическом постулате. При построении нейронных сетей правило Хебба необходимо для изменения силы связей между клетками, однако компьютерные модели не будут работать, если исходить только из этого принципа научения. Интересно отметить, что отвергнутая Хеббом теория Павлова при анализе деятельности мозга учитывает как процессы возбуждения, так и явления торможения.
Вопросы для обсуждения
1. Обсудите наблюдения, которые Хебб сделал в начале своей научной карьеры и которые он позднее пытался объяснить с точки зрения своей нейрофизиологической теории.
2. Обсудите предложенные Лэшли концепции общей активности и эквипотенциальное™.
3. Опишите синтетическую концепцию мозга. Какие аргументы Хебб выдвигал против этой концепции и что он предложил в качестве альтернативы?
4. Почему, по мнению Хебба, нейрофизиологические объяснения научения были непопулярны в то время, когда он стал профессором в университете Мак-Гилл?
5. Обсудите концепции констелляции клеток и фазовой последовательности.
6. В чем, по мнению Хебба, заключается различие между научением в детстве и во взрослом возрасте?
7. Опишите эффекты сенсорной депривации и объяснение, предложенное для этого явления Хеббом.
8. Как Хебб мог бы объяснить сновидения? Почему, например, взрыв в карбюраторе автомобиля может спровоцировать у человека сон, связанный с преступлением?
9. Как Хебб мог бы объяснить гештальт-принцип завершения?
10. Кратко изложите суть исследований страха, проводившихся Хеббом. Что он обнаружил и как объяснил наблюдавшиеся явления?
11. Что Хебб подразумевал под оптимальным уровнем стимуляции?
12. Опишите различия между ключевой и активирующей функциями стимула.
13. Почему, по мнению Хебба, люди иногда отходят от обычного для них образа жизни в поисках стимуляции?
14. Опишите связь между теориями активации и подкрепления.
15. В чем заключаются различия между кратковременной и долговременной памятью? В свой ответ включите объяснение консолидации и связанных с ней явлений.
16. Опишите уникальные характеристики подкрепления с помощью прямой стимуляции мозга.
17. Кокаин стимулирует высвобождение дофамина в клетках nucleus accumbens. В чем подкрепляющие свойства кокаина сходны с подкрепляющим действием прямой стимуляции мозга?
18. Кратко расскажите об исследованиях Сперри на расщепленном мозге.
19. Перечислите функции, по-видимому, связанные с деятельностью правого и левого полушарий мозга.
20. Опишите, каким образом Кимура использовала дихотомическое прослушивание для исследования межполушарной асимметрии.
21. Почему может быть неверно описание деятельности полушарий мозга с помощью дихотомии?
Часть VI
Эволюционная теория
Г лава 15
Дата: 2019-04-23, просмотров: 530.