Молекули хлорофілу, які перетворюють сонячну енергію, локалізуються приблизно в 1000 тилакоїдів кожного хлоропласту таким чином, що за відповідного освітлення одиницею поглинання є переважна їх більшість. Водночас вони є своєрідним «озером», у якому плаває множина так званих фотосинтетичних одиниць.
Уявлення про фотосинтетичну одиницю (ФСО) як комплекс пігментів, що бере участь у поглинанні, збиранні квантів і передачі їх спеціалізованим молекулам хлорофілу, майже одночасно було сформульовано Дж. Гафроном, Р. Емерсоном і У. Арнольді. На їхню думку, основна маса молекул пігментів, які складають ФСО, забезпечує збирання енергії та передачі її спеціальним компонентам, що фотохімічно перетворюють цю енергію. Таким чином вони виконують функцію пускового механізму фотосинтезу (рис. 15).
Такий функціонально активний комплекс було названо реакційним центром (РІД). Фотохімічні реакції здійснюють лише пігменти РІД, тоді як усі інші лише поглинають світло і переносять енергію збудження на хлорофіли реакційних центрів. Згодом було доведено, що один РІД припадає приблизно на 400...600 молекул пігменту. Частка пігментів РІД становить близько 1 % від суми всіх пігментів хлоропластів.
Загальноприйняті нині уявлення про фотохімічні реакції хлорофілів РІД сформувалися наприкінці 50-х років минулого століття під впливом класичних робіт Р. Емеосона зі співробітниками. Вони експериментально довели, що в процесі фотосинтезу рослин беруть участь дві різні фотохімічні реакції. Як виявилося, швидкість фотосинтезу за спільної дії квантів червоного світла (700 нм) та короткохвильового (680 нм) була більшою, ніж сума швидкостей фотосинтезу в разі окремого освітлення цими квантами (ефект підсилення Емерсона).
Зокрема, квантовий вихід фотосинтезу (максимальне число О2, які звільняються в перерахунку на кожний поглинутий квант) неадитивно зростав, інколи аж до 40 %. Такий ефект підсилення було пояснено функціонуванням двох фотохімічних систем (ФС І та ФС II), які послідовно виконують первинні фотохімічні реакції. Нині навіть вдалося розділити фотосистеми як за допомогою інгібіторів фотосинтезу (хімічне розділення), так і диференційним центрифугуванням (механічне розділення). Тилакоїдну мембрану вдалося розділити на дві фракції — легку та важку, які осаджувалися відповідно при 24 000 та 144 000 д. До легкої входять в основному фрагменти мембран тилакоїдів строми, які є складовими частками ФС І. Аналіз важкої фракції показав, що вона збагачена компонентами ФС II.
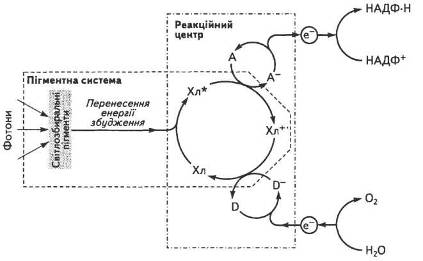
Рис. 15. Спрощена схема роботи пігментної системи хлоропласта
Згідно з сучасними уявленнями, хлорофіли реакційних центрів виконують основну функцію в перетворенні енергії сонячних квантів. Завдяки специфічним зв'язкам та організації їхніх молекул-донорів і молекул-акцепторів хлорофіли здатні використовувати одержану енергію збудження для утворення потоку електронів шляхом оборотних окисно-відновних реакцій. В результаті цього в тилакоїдній мембрані формується протонний і електричний градієнти. Максимуми поглинання хлорофілів реакційних центрів двох систем, за яких відбуваються окисно-відновні процеси, різні: максимум хлорофілу реакційного центру ФС І (Р700) — 700 нм, тоді як хлорофілу реакційного центру ФС II (Р680) — 680 нм. Кожен із хлорофілів має відповідні донори та акцептори електронів, які започатковують серію окисно-відновних перетворень відповідних переносників електронів у тилакоїдній мембрані. Ймовірна схема структури реакційних центрів ФС І і ФС II наведена на рис. 16.
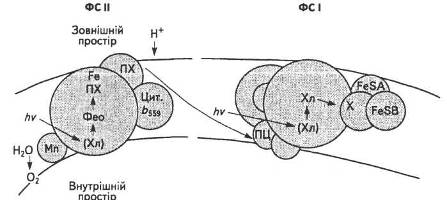
Рис. 16. Реакційні центри ФС І і ФС II
Донори електронів локалізовані всередині, тоді як їхні акцептори — зовні тилакоїдної мембрани, що і забезпечує миттєвий розподіл заряду на мембрані.
Фотосинтетична одиниця (ФСО) — це мінімальне функціональне угруповання тилакоїда, здатне до фотосинтетичного розподілу заряду і складається з пігментів антенного комплексу та реакційних центрів.
За останніми даними, на один хлоропласт припадає 2 • 106 ФСО. Ці найдрібніші одиниці здатні здійснювати окиснення й відновлення НАДФ+, мають антенні комплекси пігментів-світлозбирачів, двіфотосистеми, що формують разом із багатьма переносниками електронно-транспортний ланцюг (ЕТЛ) тилакоїдної мембрани.
Досі немає єдиної думки щодо структури ФСО. Насамперед, було запропоновано моноцентральну модель ФСО, згідно з якою в усьому комплексі є лише один РІД. Більшість учених віддають перевагу мультицентральній моделі, яка припускає наявність кількох реакційних центрів (рис. 17).

Рис. 17. Модель мультииентральної (а) та уніцентральної (б) фотосинтетичних одиниць:
о — пігмент антенного комплексу;
• — Р70О або Р680 в реакційному центрі;
* — молекула пігменту антени, яка перейшла у фотозбуджений стан після поглинання фотона або внаслідок перенесення енергії
В хлоропластах вищих рослин існує як мінімум два типи фото-;истем: ФС І і ФС II.
Фотосистема — це дискретна одиниця організації в тилакоїдах певних пігментів разом з іншими молекулами — переносниками електронів і протонів, які беруть участь у світловій стадії фотосинтезу.
Точна локалізація фотосистем у тилакоїдній мембрані хлоропластів поки що не відома. Вважають, що мембрани тилакоїдів строми містять переважно ФС І, а мембрани гран — ФС І і ФС II. Таким чином, у тилакоїдній мембрані молекули пігментів і переносники електронів об'єднані в надмолекулярні функціональні комплекси.
Z-cxeмa фотосинтезу
Р. Хілл і Ф. Бендалл запропонували схему електронної стадії фотосинтетичного процесу, яка після низки уточнень одержала назву Z-схеми фотосинтезу (рис. 18).
Переносники електронів ЕТЛ розміщуються відповідно до зменшення електровід'ємного редокс-потенціалу. В фотосинтезі вищих рослин перенесення електрона від двох молекул води (окисно-відновний потенціал 0,82 В) до НАДФ+ (окисно-відновний потенціал — 0,32 В) здійснюється за рахунок енергії чотирьох квантів світла з участю відповідно двох фотосистем, що діють послідовно. Механізм передачі електронів в обох фотосистемах — одноелектронний.
Під час поглинання кванта світла ФС II за період, менший за 105 с, електрон від води переноситься в ЕТЛ, проти термодинамічного потенціалу, а саме: від позитивного редокс-потенціалу фередоксину на НАДФ+. Саме в цьому полягає основна суть фотосинтезу — адже в процесі такого транспортування запасається вільна енергія поглинутих квантів. У мембрані тилакоїду розміщується близько 200 таких ланцюгів.
Шлях електрона від води до НАДФ+ з участю обох фотосистем називають нециклічним.
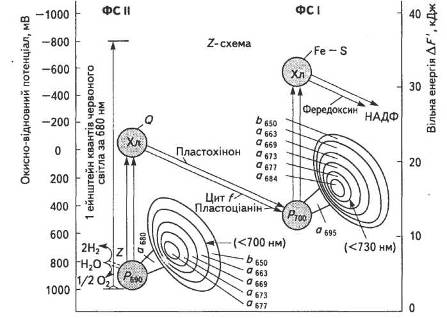
Рис. 18. Z-схема фотосинтезу
Існує й циклічне транспортування електронів з участю лише ФС І, яке надходить від неідентифікованого компонента X на фередоксин, далі — на цитохром Ь6 (Ео = — 0,18 В) і пластохінон (Пх). З відновленого Пх • Н2 електрони транспортуються на цитохром f, далі — на пластоціанін і, нарешті, заповнюють дірку того самого пігменту Р700.
Можливий також псевдоциклічний шлях транспортування електрона, коли в результаті передачі електрона на кисень він відновлюється до Н2О, чим практично маскується процес фотолізу води в ФС II (рис. 19).
Як видно зі Z-схеми, на шляху від однієї молекули Н2О до НАДФ+ крізь переносники передаються два електрони. Якщо один квант енергії, що надходить в кожну фотосистему, піднімає на вищий рівень лише один електрон, то для перенесення двох електронів треба мінімум чотири кванти. Світлозбиральний апарат, який постачає енергію, складається з лабільного світлозбпрального комплексу (СЗК), що обслуговує обидві фотосистеми, та пігмент-білкових антенних комплексів ФС І і ФС II. Уявлення про Z-схему фотосинтезу на цей час є загальноприйнятим, хоча постійно піддається певним змінам і доповненням.
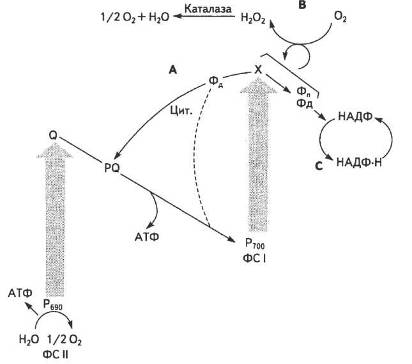
Рис. 19. Можливі шляхи перенесення електронів у хлоропластах:
А — циклічний; В — псевдоциклічний; С — нециклічний
Висновки
Особливості морфології, анатомії та фізіології листка повною мірою забезпечують здійснення його основної функції — фотосинтезу. Функціонування продихів регулює газообмін — надходження вуглекислого газу, виділення кисню та випаровування води.
Фотосинтетичні пігменти — досить складні органічні сполуки, тому їх біосинтез охоплює низку етапів. Хлорофіли, як і подібні до них за хімічними властивостями цитохроми (Fe-порфірини), характеризуються єдиним процесом синтезу від простих складових до тетрапірольних структур.
Пігменти пластид відносять до трьох класів, а саме: хлорофіли, каротиноїди та фікобіліни. Найважливіше значення в процесі фотосинтезу належить зеленим пігментам — хлорофілам. Нині відомо близько десяти хлорофілів, які відрізняються хімічним складом, забарвленням і поширенням серед живих організмів. Основними пігментами, без яких фотосинтез не відбувається, є хлорофіл а для зелених рослин і бактеріохлорофіл для фототрофних бактерій. Пігменти — це сполуки, які вибірково поглинають світло у видимій (400...700 нм) частині спектра.
Непоглинені ділянки сонячного спектра відбиваються, що й зумовлює забарвлення пігменту. Зелений пігмент хлорофіл поглинає червоні та сині промені, тоді як зелені в основному відбиваються.
Фотосинтез — це насамперед фізична проблема. Первинні процеси його розпочинаються з миттєвих реакцій поглинання електромагнітної енергії сонячних квантів, її запасання та стабілізації у вигляді електронного збудження, міграції енергії в пігмент-ліпопротеїдному комплексі хлоропластів.
Світлоіндуковане векторне транспортування електронів між двома фотосистемами, яке розпочинається від води й закінчується НАДФ+, забезпечується завдяки специфічному й несиметричному розташуванню низки окисно-відновних компонентів тилакоїдної мембрани — переносників електронів і протонів, які в сукупності складають електронно-транспортний ланцюг.
Список літератури
1. Бугай С.М. Растенееводство. – К.: Вища школа, 1975. – 375 с.
2. Избранные главы физиологии растений / В. Ф. Гавриленко, М. В. Гусев, К. А. Никитина. — М.: Изд-во Моск. ун-та, І986. — 436 с.
3. Кочубей С. М. Организация фотосинтетического аппарата высших растений. — Киев: Альтерпрес, 2001. — 204 с.
4. Мокроносов А. Т., Гавриленко В. Ф. Фотосинтез: физиологические и биохимические аспекты. — М.: Изд-во Моск. ун-та, 1992. — 319 с.
5. Максимов Н.А. Краткий курс физиологии растений, 1958.
6. Мусієнко М. М. Фотосинтез. — Киев: Вища школа, 1995. — 247 с.
7. Мусієнко М.М. Фізіологія рослин. – К.: Либідь, 2005. – 808 с.
8. Рубин Б.А. Курс физиологии растений. – М.: Высшая школа, 1961. –583 с.
9. Силаева А. Й. Структура хлоропластов и факторы среды. — Киев: Наук, думка, 1978. — 157 с.
10. Фотосинтез: В 2 т. / Под ред. Р. Говинджи. — М.: Мир, 1987. — Т. 1. 728 с; Т. 2. 460 с.
11. Эдварде Дж., Уокер Д. Фотосинтез С3- и С4-растений: механизмы и регуляция. — М.: Мир, 1986. — 590 с.
Дата: 2019-05-29, просмотров: 845.