ЗМІСТ
Вступ. 2
Розділ 1. Значення фотосинтезу. 3
1.1 Суть та значення фотосинтезу. 3
1.2 Загальне рівняння фотосинтезу та походження кисню.. 7
Розділ 2. Листок як орган фотосинтезу. 13
Розділ 3. Фотосинтетичні пігменти листка. 19
3.1 Хлорофіли. 19
3.2 Каротиноїди. 22
3.3 Фікобіліни. 24
3.4 Оптичні властивості фотосинтетичних пігментів. 27
3.5 Біосинтез пігментів фотосинтетичного апарату рослин. 28
Розділ 4. Енергетика фотосинтезу. 33
4.1 Первинні процеси фотосинтезу. 33
4.2 Фотосинтетична одиниця, реакційний центр, фотосистема. 36
4.3 Z-cxeмa фотосинтезу. 40
Висновки. 43
Список літератури. 45
Вступ
Нині людство все більше розуміє ту очевидну істину, яку вперше обґрунтували видатні вчені К. А. Тімірязєв і В. І. Вернадський, що екологічне благополуччя біосфери, а отже й існування самої людини та цивілізації в цілому, визначається станом рослинного покриву планети. Значення фотосинтезу в біосферних процесах Землі настільки велике й різноманітне, а його природа настільки унікальна, що проблема фотосинтезу правомірно вважається однією з найважливіших проблем не лише науки, а й практики. Більш як 3,5 млрд. років тому відбувалася подія, якій судилося стати могутньою рушійною силою еволюції органічної матерії — виник фотосинтез. Еволюційний процес випробував різні варіанти енергозабезпечення життя. Перші прокаріоти типу бактерій були ферментуючими гетеротрофами, які добували їжу шляхом розщеплення органічних речовин абіотичного походження. Життя набуло якісно нове, практично невичерпне джерело енергії — Сонце.
Прокаріоти — це ціанобактерії, що дали початок розвитку рослинного царства, знайшли ключ до фотоавтотрофії. Центральне місце в цьому процесі зайняв зелений пігмент — хлорофіл. Один з основоположників фізіології рослин К. А. Тімірязєв, підкреслюючи космічне значення зеленої рослини, образно писав, що зелений листок, а вірніше хлорофіл, є фокусом у світовому просторі, в який, з одного боку, йде енергія Сонця, а з другого — беруть початок усі прояви життя на Землі.
Мета роботи охарактеризувати протікання процесів живлення рослин вуглецем.
Завдання роботи:
1) розглянути значення та суть процесу фотосинтезу;
2) дати характеристику листку як органу фотосинтезу;
3) дати характеристику основних фото синтезуючих пігментів листка;
4) розглянути енергетичні процеси фотосинтезу.
Розділ 1. Значення фотосинтезу
Розділ 3. Фотосинтетичні пігменти листка
Хлорофіли
Хлорофіли — це складні ефіри дикарбонової кислоти хлорофіліну, в якої одна карбоксильна група етирифікована залишком метилового спирту, а і інша — залишком спирту фітолу.
Методом хроматографічного аналізу хлорофіли було розділено на хлорофіл а — C55H72O5N4Mg Ta хлорофіл b — C55H70O6N4Mg.
Основу молекули становить порфіринове ядро, що складається з чотирьох пірольних кілець, атоми азоту яких сполучені в центрі магнієм. По периферії пірольні кільця зв'язані між собою метино-вими містками (=СН—). Крім того, є додаткове п'яте циклопента-нове кільце, яке містить кетогрупу (=С=О).
Гідрофільні властивості хлорофілу пов'язані з його азотовмісним порфіриновим ядром — «головкою» молекули. Довгий вуглецевий ланцюг (фітольний «хвіст»), який приєднується до порфіринової частини молекули, надає хлорофілу гідрофобних властивостей. Здатність фітолу розчинятися в жирах має важливе значення для збереження структури хлоропластів. Саме завдяки наявності залишку фітолу хлорофіли добре розчинні в спирті, бензині, ацетоні, але нерозчинні у воді. Протилежні гідрофільні та гідрофобні властивості цих частин молекули мають важливе значення під час локалізації хлорофілу в ламелах хлоропласта між білками та ліпідами. Структурну формулу хлорофілів наведено на рис. 6.
Хлорофіл а наявний у хлоропластах або в аналогічних структурах усіх організмів, здатних до фотосинтезу, за винятком бактерій які містять бактеріохлорофіл а.
Усі вищі рослини та зелені водорості містять також хлорофіл b Різниця між ними полягає в тому, що до хлорофілу ь біля третього атома вуглецю другого пірольного кільця замість метильної групи (-СН3), яка характерна для хлорофілу а, приєднана альдегідна (-СОН)-група. Ці два хлорофіли відрізняються також за забарвленням: хлорофіл а має синьо-зелений відтінок, тоді як хлорофіл b -жовто-зелений. Вміст хлорофілу а в листку приблизно втроє більший, ніж хлорофілу b.
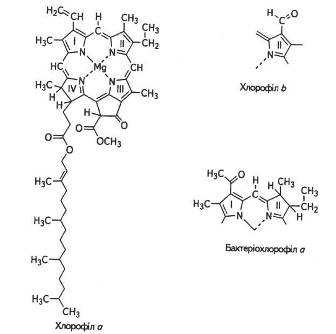
Рис. 6. Структурна формула хлорофілів
Молекула хлорофілу електрично нейтральна, бо подвійний позитивний заряд магнію компенсується двома надлишковими електронами, які розподілені між чотирма атомами азоту пірольних кілець.
Якщо з молекули хлорофілу видалити фітол, одержимо хлорофілід.
У разі заміщення атома Мд2+ воднем утворюється феофітин. У різних представників рослинного світу трапляються й інші види хлорофілів та інших пігментів.
Різні види пігментів розпізнають за допомогою їхніх спектральних характеристик (рис. 7).
Спектри поглинання у різних груп хлорофілів залежать від характеру заміщувальної групи біля пірольних кілець і типу органічного розчинника. Так, бактеріохлорофіл а має спектр поглинання з максимумами 400, 600 і 700 нм. Завжди чітко вирізняються два максимуми поглинання: в області коротких хвиль та відповідно в довгохвильовій. Наприклад, спектри поглинання хлорофілу а — 420 і 662 нм, а хлорофілу b — 455 і 644 нм. В листку спектри поглинання хлорофілу змінюються залежно від його стану, ступеня агрегації йадсорбції на певних білках. Виявлено форми хлорофілів, які поглинають світло з довжиною хвилі 700, 710 і навіть 720 нм.
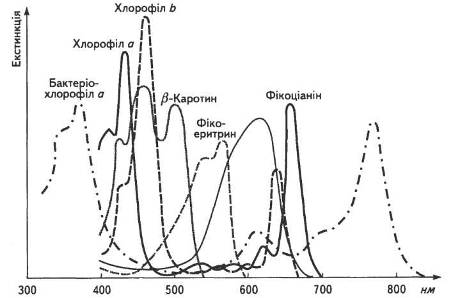
Рис. 7. Спектральні характеристики пігментів
Хлорофіл у живій інтактній клітині здатний до фотоокиснення та фотовідновлення. Здатність до окисно-відновних реакцій пояснюється наявністю в молекулі хлорофілу подвійних зв'язків з рухливими π-електронами та атомів азоту з неподіленими електронами. Азот пірольних кілець може окиснюватися (віддавати електрони) або відновлюватися (приймати електрони).
Важливою властивістю хлорофілів є їхня здатність до взаємодії один з одним, тому в хлоропластах можуть траплятися хлорофіли як у мономерніп, так і в агрегованій формі.
Встановлено, що хлорофіл у мембранах пластид перебуває у вигляді пігмент-ліпопротеїдних комплексів з різним ступенем агрегації. Особливості спектра дії фотосинтезу свідчать про те, що в процесах поглинання світла беруть участь і додаткові пігменти.
Каротиноїди
Поряд із зеленими пігментами в хлоропластах є й такі, що належать до групи каротиноїдів. Каротиноїди — це найпоширеніші в рослинному світі жиророзчинні жовті, оранжеві та червоні пігменти аліфатичної будови. Вони є обов'язковим компонентом фотосинтетичного апарату. За хімічною природою всі вони — полімери вуглеводню і складають ланцюг із 40 вуглецевих атомів (рис. 4.9), побудований з 8 залишків ізопрену.
Довжина ланцюгів досягає 3 нм і вони часто закінчуються шестичленними циклами. Каротиноїди можуть бути ациклічними (лікопін), моноциклічними або біциклічними. Окиснені форми каротиноїдів називають ксантофілами. Ксантофіли становлять близько 50 % всіх каротиноїдів листка.
У хлоропластах вищих рослин поряд із хлорофілами найчастіше трапляються β-каротин (С40Н56) і ксантофіли — лютеїн (С40Н56О2) і віолаксантин (С40Н56О4). В значних кількостях є також а-каротин і неоксантин.
Каротини, як і ксантофіли, характеризуються гідрофобними властивостями, тому добре розчинні в жирах, що дає їм змогу формувати комплекси в ліпідному прошарку мембрани. Ксантофіли, порівняно з каротинами, мають різноманітнішу структуру, бо до їхнього складу можуть входити різні групи, що містять О2, а саме: гідроксильні, метоксильні, кетогрупи та інші. Між собою ксантофіли відрізняються лише ступенем окиснення.
Як допоміжні пігменти фотосинтетичного апарату каротиноїди забезпечують поглинання квантів двома піками в синьо-фіолетовий та синій областях спектра (420...490 нм) і деякою мірою в зеленій (490...550 нм). Максимуми поглинання залежать від замісників біля вуглецевого скелета, типу розчинника та кількості подвійних зв'язків. В амплітуді від 400 до 550 нм вони, як правило, мають дві-три смуги поглинання.
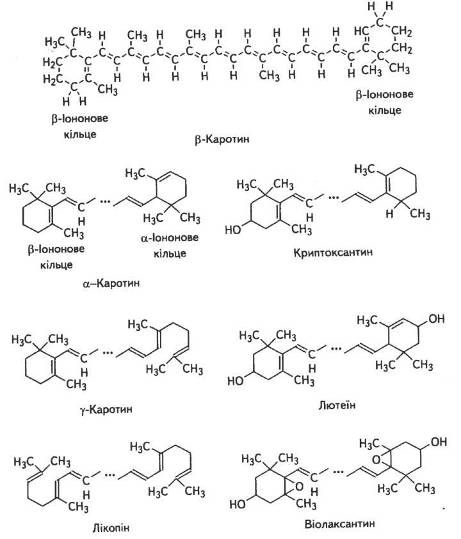
Рис. 8. Структурні формули каротиноїдів
Отже, каротиноїди розширюють спектр дії фотосинтезу, забезпечуючи, поглинання від 10 до 20 % енергії сонячних квантів, причому близько 50 % енергії поглинається в короткохвильовій області — зоні високих енергій. Ці пігменти виконують функцію світлопоглинання, передаючи енергію свого електронно-збудженого стану до хлорофілу а. Зворотний процес передачі неможливий. Слід підкреслити, що каротиноїди, на відміну від хлорофілів, не здатні до флуоресценції.
Такі каротиноїди, як віолаксантин, неоксантин, зеаксантин та інші, поглинаючи світло в короткохвильовій високоенергетичній частині спектра, виконують захисну функцію, як хімічні буфери в реакціях фотосинтезу. Можливий механізм захисту полягає в тому, що каротиноїди здатні реагувати зі збудженою молекулою хлорофілу, забираючи від нього енергію, чим попереджають його фотоокиснення. Енергія фотозбудженої молекули хлорофілу переходить до каротиноїду, хлорофіл набуває нормального енергетичного стану, а енергія виділяється у вигляді тепла. Завдяки цьому каротиноїди оберігають хлорофіл та інші біологічно активні сполуки від фотоокиснення.
Слід згадати, що каротиноїдам належить ще одна специфічна функція в регулюванні фотосинтетичного апарату рослинного організму. Річ у тім, що хлоропласти переміщуються в клітині під впливом синіх променів, які знову ж таки поглинаються каротиноїдами.
Фізіологічна функція каротиноїдів не обмежується їхньою участю в передачі енергії на молекули хлорофілів. Каротиноїди — переносники активного кисню, вони беруть участь в окисно-відновних реакціях завдяки наявності значної кількості подвійних зв'язків. їм належить певна функція у статевому процесі рослин, а саме: вони зумовлюють забарвлення пелюсток квітів, плодів, коренеплодів. Залишається мало з'ясованою їхня функція в кисневому обміні, участь у формуванні фотоперіодичної реакції, в ростових процесах, зокрема під час проростання насіння, в проявах фототаксису та фототропізму.
Фікобіліни
Такі фотосинтезуючі організми, як синьо-зелені водорості, червоні та деякі інші представники водоростей, окрім хлорофілів і каротиноїдів, мають ще одну групу допоміжних пігментів — фікобіліни. Цю назву вони одержали завдяки своїй подібності до тваринних пігментів — білірубінів.
Білінами називають досить поширені в природі тетрапіроли з відкритим ланцюгом, без металу. В тваринному царстві вони часто трапляються як нефункціональні катаболіти гема. Рослинні біліни (фікобіліни) функціонують як сенсибілізатори і фоторецептори, що забезпечується їхнім ковалентним зв'язком з апобілками. Отже, подібно до хлорофілів, фікобіліни — тетрапіроли, але чотири залишки піролу в них створюють незамкнутий витягнутий або згорнутий ланцюг. Піроли з'єднуються між собою метиленовими і метиновими містками, тоді як хромофорні групи, як правило, ковалентно зв'язані білком.
Хромофорною групою фікоціанінів є фікоціанобілін, фікоерит-ринів — фікоеритробілін (рис. 9).
Відрізняються вони між собою лише взаєморозміщенням пірольних і піроленінових кілець. У фікобілінпротеїнах одна молекула білка може зв'язувати кілька хромофорних груп. Фікоеритробілін у сполученні з білком називають фікоеритрином. Він добре розчинний у воді, поглинає кванти зеленої частини спектра. Фікоеритробіліни (C34H47N4O8) трапляються переважно у червоних водоростей, де й визначають їхнє забарвлення, адже фікоеритрини — білки червоного кольору, які мають максимуми поглинання 500...568 нм.
Фікоціанобіліни (C34H42N4O9) характерні для синьозелених водоростей, причому максимуми поглинання фікоціанінів дещо зсунуті в довгохвильову область — від 585 до 630 нм.
Фікоеритрин і фікоціанін складаються з двох різних білкових субодиниць, які позначають відповідно а (молекулярна маса 19 000) і р (молекулярна маса 21 000) у співвідношенні 1 : 1. Кожна з білкових субодиниць несе ковалентно зв'язаний з нею фікобілін. Як правило, фікобіліни, зв'язані з однією субодиницею, належать до одного типу, тобто до фікоеритробілінів або фіко-ціанобілінів.
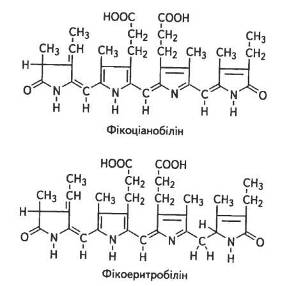
Рис. 9. Хромофорні групи фікоціанінів і фікоеритринів
До фікобілінів належать також алофікоціаніни — пігменти з максимумами поглинання від 585 до 650 нм, хромофорну групу їх поки що не визначено. Цю назву алофікоціаніни одержали завдяки тому, що спочатку їх приймали за одну з форм фікоціаніну. На відміну від інших фікобілінпротеїнів їхня молекула складається лише з одного типу білкових одиниць (молекулярна маса 15 кДа), причому кожна субодиниця несе одну молекулу ало-фікоціаніну.
Алофікоціанін характерний для синьозелених, хоча трапляється також у червоних і криптомонадових водоростей. А загалом поширення окремих типів пігментів фікобілінів серед синьозелених, червоних і бурих водоростей досить специфічне.
Слід зазначити, що на відміну від хлорофілів і каротиноїдів, локалізованих у ламелах, фікобіліни концентруються або в стромі, або формують особливі впорядковані ансамблі на поверхні мембран — фікобілісоми (рис. 10).
Як правило, в таких водоростях фікобіліни присутні в значно більшій концентрації порівняно з хлорофілами, тому саме вони і зумовлюють їхнє забарвлення. Фікоціаніни, фікоеритрини і алофікоціаніни трапляються в різних співвідношеннях, причому залежно від умов освітлення формується переважно такий набір пігментів, який найкраще використовує відповідний спектп.
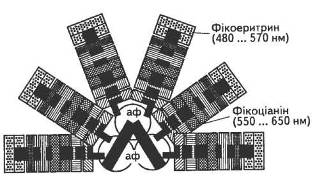
Рис. 10. Модель фікобілісоми: аф — алофікоціаніни
Пігменти даної групи виконують функцію світлозбиральної антени та забезпечують ефективну передачу поглинутої ними енергії сонячного світла до хлорофілів а.
Фікобіліни зумовлюють явище філогенетичної хроматичної адаптації водоростей в їхній вертикальній зональності.
Як відомо, червоні промені, що відповідають спектру поглинання хлорофілу, поглинаються, проходячи крізь товщу води. Вже на глибині 34 м зникають червоні промені, 177 м — жовті, 322 м — зелені, 500 м — сині та фіолетові. Відповідно до цього певні екологічні ніші займають спочатку зелені, потім синьо-зелені і, нарешті, червоні водорості. Наявність фікобілінів дає змогу водоростям у процесі фотосинтезу використовувати промені, які проникають на певну глибину, та займати відповідні екологічні ніші.
Z-cxeмa фотосинтезу
Р. Хілл і Ф. Бендалл запропонували схему електронної стадії фотосинтетичного процесу, яка після низки уточнень одержала назву Z-схеми фотосинтезу (рис. 18).
Переносники електронів ЕТЛ розміщуються відповідно до зменшення електровід'ємного редокс-потенціалу. В фотосинтезі вищих рослин перенесення електрона від двох молекул води (окисно-відновний потенціал 0,82 В) до НАДФ+ (окисно-відновний потенціал — 0,32 В) здійснюється за рахунок енергії чотирьох квантів світла з участю відповідно двох фотосистем, що діють послідовно. Механізм передачі електронів в обох фотосистемах — одноелектронний.
Під час поглинання кванта світла ФС II за період, менший за 105 с, електрон від води переноситься в ЕТЛ, проти термодинамічного потенціалу, а саме: від позитивного редокс-потенціалу фередоксину на НАДФ+. Саме в цьому полягає основна суть фотосинтезу — адже в процесі такого транспортування запасається вільна енергія поглинутих квантів. У мембрані тилакоїду розміщується близько 200 таких ланцюгів.
Шлях електрона від води до НАДФ+ з участю обох фотосистем називають нециклічним.
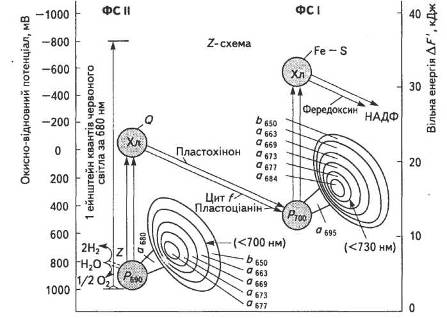
Рис. 18. Z-схема фотосинтезу
Існує й циклічне транспортування електронів з участю лише ФС І, яке надходить від неідентифікованого компонента X на фередоксин, далі — на цитохром Ь6 (Ео = — 0,18 В) і пластохінон (Пх). З відновленого Пх • Н2 електрони транспортуються на цитохром f, далі — на пластоціанін і, нарешті, заповнюють дірку того самого пігменту Р700.
Можливий також псевдоциклічний шлях транспортування електрона, коли в результаті передачі електрона на кисень він відновлюється до Н2О, чим практично маскується процес фотолізу води в ФС II (рис. 19).
Як видно зі Z-схеми, на шляху від однієї молекули Н2О до НАДФ+ крізь переносники передаються два електрони. Якщо один квант енергії, що надходить в кожну фотосистему, піднімає на вищий рівень лише один електрон, то для перенесення двох електронів треба мінімум чотири кванти. Світлозбиральний апарат, який постачає енергію, складається з лабільного світлозбпрального комплексу (СЗК), що обслуговує обидві фотосистеми, та пігмент-білкових антенних комплексів ФС І і ФС II. Уявлення про Z-схему фотосинтезу на цей час є загальноприйнятим, хоча постійно піддається певним змінам і доповненням.
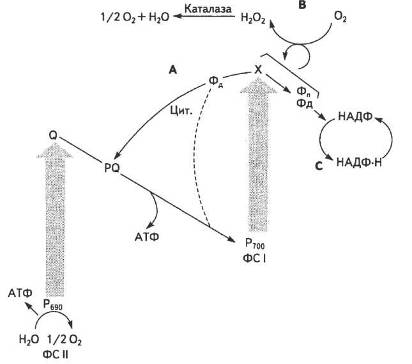
Рис. 19. Можливі шляхи перенесення електронів у хлоропластах:
А — циклічний; В — псевдоциклічний; С — нециклічний
Висновки
Особливості морфології, анатомії та фізіології листка повною мірою забезпечують здійснення його основної функції — фотосинтезу. Функціонування продихів регулює газообмін — надходження вуглекислого газу, виділення кисню та випаровування води.
Фотосинтетичні пігменти — досить складні органічні сполуки, тому їх біосинтез охоплює низку етапів. Хлорофіли, як і подібні до них за хімічними властивостями цитохроми (Fe-порфірини), характеризуються єдиним процесом синтезу від простих складових до тетрапірольних структур.
Пігменти пластид відносять до трьох класів, а саме: хлорофіли, каротиноїди та фікобіліни. Найважливіше значення в процесі фотосинтезу належить зеленим пігментам — хлорофілам. Нині відомо близько десяти хлорофілів, які відрізняються хімічним складом, забарвленням і поширенням серед живих організмів. Основними пігментами, без яких фотосинтез не відбувається, є хлорофіл а для зелених рослин і бактеріохлорофіл для фототрофних бактерій. Пігменти — це сполуки, які вибірково поглинають світло у видимій (400...700 нм) частині спектра.
Непоглинені ділянки сонячного спектра відбиваються, що й зумовлює забарвлення пігменту. Зелений пігмент хлорофіл поглинає червоні та сині промені, тоді як зелені в основному відбиваються.
Фотосинтез — це насамперед фізична проблема. Первинні процеси його розпочинаються з миттєвих реакцій поглинання електромагнітної енергії сонячних квантів, її запасання та стабілізації у вигляді електронного збудження, міграції енергії в пігмент-ліпопротеїдному комплексі хлоропластів.
Світлоіндуковане векторне транспортування електронів між двома фотосистемами, яке розпочинається від води й закінчується НАДФ+, забезпечується завдяки специфічному й несиметричному розташуванню низки окисно-відновних компонентів тилакоїдної мембрани — переносників електронів і протонів, які в сукупності складають електронно-транспортний ланцюг.
Список літератури
1. Бугай С.М. Растенееводство. – К.: Вища школа, 1975. – 375 с.
2. Избранные главы физиологии растений / В. Ф. Гавриленко, М. В. Гусев, К. А. Никитина. — М.: Изд-во Моск. ун-та, І986. — 436 с.
3. Кочубей С. М. Организация фотосинтетического аппарата высших растений. — Киев: Альтерпрес, 2001. — 204 с.
4. Мокроносов А. Т., Гавриленко В. Ф. Фотосинтез: физиологические и биохимические аспекты. — М.: Изд-во Моск. ун-та, 1992. — 319 с.
5. Максимов Н.А. Краткий курс физиологии растений, 1958.
6. Мусієнко М. М. Фотосинтез. — Киев: Вища школа, 1995. — 247 с.
7. Мусієнко М.М. Фізіологія рослин. – К.: Либідь, 2005. – 808 с.
8. Рубин Б.А. Курс физиологии растений. – М.: Высшая школа, 1961. –583 с.
9. Силаева А. Й. Структура хлоропластов и факторы среды. — Киев: Наук, думка, 1978. — 157 с.
10. Фотосинтез: В 2 т. / Под ред. Р. Говинджи. — М.: Мир, 1987. — Т. 1. 728 с; Т. 2. 460 с.
11. Эдварде Дж., Уокер Д. Фотосинтез С3- и С4-растений: механизмы и регуляция. — М.: Мир, 1986. — 590 с.
ЗМІСТ
Вступ. 2
Розділ 1. Значення фотосинтезу. 3
1.1 Суть та значення фотосинтезу. 3
1.2 Загальне рівняння фотосинтезу та походження кисню.. 7
Розділ 2. Листок як орган фотосинтезу. 13
Розділ 3. Фотосинтетичні пігменти листка. 19
3.1 Хлорофіли. 19
3.2 Каротиноїди. 22
3.3 Фікобіліни. 24
3.4 Оптичні властивості фотосинтетичних пігментів. 27
3.5 Біосинтез пігментів фотосинтетичного апарату рослин. 28
Розділ 4. Енергетика фотосинтезу. 33
4.1 Первинні процеси фотосинтезу. 33
4.2 Фотосинтетична одиниця, реакційний центр, фотосистема. 36
4.3 Z-cxeмa фотосинтезу. 40
Висновки. 43
Список літератури. 45
Вступ
Нині людство все більше розуміє ту очевидну істину, яку вперше обґрунтували видатні вчені К. А. Тімірязєв і В. І. Вернадський, що екологічне благополуччя біосфери, а отже й існування самої людини та цивілізації в цілому, визначається станом рослинного покриву планети. Значення фотосинтезу в біосферних процесах Землі настільки велике й різноманітне, а його природа настільки унікальна, що проблема фотосинтезу правомірно вважається однією з найважливіших проблем не лише науки, а й практики. Більш як 3,5 млрд. років тому відбувалася подія, якій судилося стати могутньою рушійною силою еволюції органічної матерії — виник фотосинтез. Еволюційний процес випробував різні варіанти енергозабезпечення життя. Перші прокаріоти типу бактерій були ферментуючими гетеротрофами, які добували їжу шляхом розщеплення органічних речовин абіотичного походження. Життя набуло якісно нове, практично невичерпне джерело енергії — Сонце.
Прокаріоти — це ціанобактерії, що дали початок розвитку рослинного царства, знайшли ключ до фотоавтотрофії. Центральне місце в цьому процесі зайняв зелений пігмент — хлорофіл. Один з основоположників фізіології рослин К. А. Тімірязєв, підкреслюючи космічне значення зеленої рослини, образно писав, що зелений листок, а вірніше хлорофіл, є фокусом у світовому просторі, в який, з одного боку, йде енергія Сонця, а з другого — беруть початок усі прояви життя на Землі.
Мета роботи охарактеризувати протікання процесів живлення рослин вуглецем.
Завдання роботи:
1) розглянути значення та суть процесу фотосинтезу;
2) дати характеристику листку як органу фотосинтезу;
3) дати характеристику основних фото синтезуючих пігментів листка;
4) розглянути енергетичні процеси фотосинтезу.
Розділ 1. Значення фотосинтезу
Дата: 2019-05-29, просмотров: 538.