Фотосинтетичні пігменти — досить складні органічні сполуки, тому їх біосинтез охоплює низку етапів. Хлорофіли, як і подібні до них за хімічними властивостями цитохроми (Fe-порфірини), характеризуються єдиним процесом синтезу від простих складових до тетрапірольних структур (виразний приклад єдності шляхів метаболізму у тварин і рослин). Шляхом тетрапірольного синтезу утворюються й інші речовини, які використовують світло як сигнал (наприклад, фітохромна система), так що тетрапірольні структури формують майже весь залежний енергетичний та інформаційний апарат рослин.
Доведено, що в мембранах етіопластів і хлоропластів локалізовані специфічні центри біосинтезу пігментів, де й зосереджуються їхні попередники — протохлорофіліди та хлорофіліди. Реакції, що забезпечують синтез протохлорофіліду, відбуваються за відсутності світла, тоді як утворення з протохлорофіліду хлорофіл іду вимагає освітлення.
Біосинтез порфіринів у вищих рослин відбувається в три етапи:
• синтез амінолевулінової кислоти (АЛК) з п'ятивуглецевих дикарбонових (глутамінової, можливо, о-кетоглутарової) кислот;
• циклізація двох молекул АЛК з утворенням піролу порфобіліно-гена;
• утворення та циклізація тетрапіролу, формування магнійпротопорфірину, ферментативна етерифікація хлорофіліду фітолом.
У прокаріот і евкаріот АЛК синтезується із різних джерел: прокаріоти використовують для її синтезу гліцин і сукциніл-КоА, при цьому гліцин, перш за все, є джерелом азоту для побудови піроль-их кілець. Евкаріоти використовують різноманітні п'ятивуглецеві сполуки, зокрема глутамат, який спочатку фосфорилюється, а потім відновлюється.
На першому етапі синтез АЛК локалізований в хлоропластах, хоча самі ферменти, необхідні для цього, синтезуються в цитоплазмі клітини під контролем ядерних генів, а потім транспортуються в хлоропласти. Глутамінова кислота через 2-гідрооксиглута-рову перетворюється на 4,5-діоксивалеріанову (пентанову), яка потім амінується за рахунок аланіну чи інших амінокислот. Можливі й інші шляхи синтезу АЛК, які, ймовірно, локалізовані в різних компартментах клітини і по-різному регулюються. Здебільшого саме синтез АЛК обмежує швидкість біосинтезу хлорофілів, тому регуляція утворення даного проміжного продукту — надзвичайно важливий етап формування фотосинтетичного апарату рослин (рис. 11).
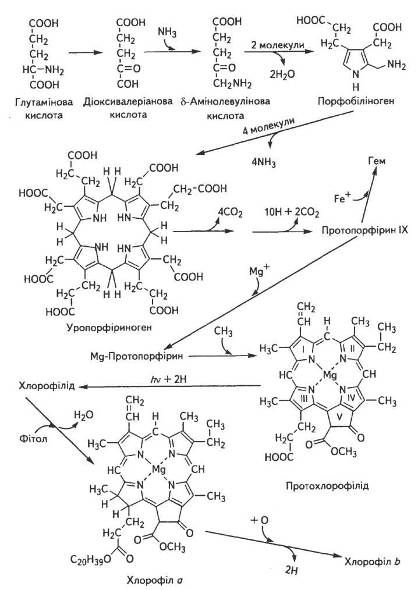
Рис. 11. Основні етапи біосинтезу хлорофілів
На другому етапі відбувається альдольна конденсація двох молекул АЛК з наступною ізомеризацією спочатку до піролу, а потім до порфобіліногену. Потім із чотирьох піролів синтезується уропорфіриноген, який поетапно перетворюється на протопорфірин IX .
На третьому етапі в результаті полімеризації чотирьох молекул порфобіліногена виникає тетрапірол, а пізніше, при замиканні кільця, утворюється уропорфіриноген І і ІІ. Перетворення порфобіліногена на уропорфіриноген ІІІ каталізується уропорфіриноген-1-синтетазою за наявності іншого білка КоА-синтетази.
Характерно, що в рослинній клітині лінійні тетрапіроли можуть бути хромофорними групами фітохрому, а циклічні — використовуватися для біосинтезу порфіринів, цитохромів, каталази чи пероксидази.
Уропорфіриноген III через серію реакцій окиснення та декарбоксилування перетворюється на протопорфірин IX. Завершальні реакції синтезу хлорофілів із протопорфірину IX повністю ще не з'ясовано, хоча відомо, що першою стадією, ймовірно, є ферментативне введення в молекулу атома магнію, потім метилування пропіонатного бічного ланцюга, насичення вінільної групи в кільці піролу IV, а відтак — замикання кільця V з утворенням протохлорофілу.
На цьому ж етапі формування молекули хлорофілу а відбувається етерифікація фітолом, який синтезується з ацетил-КоА через мевалонову кислоту.
Незважаючи на багато зусиль різних вчених, поки що нікому не вдаюся з'ясувати, здавалося б, просте питання: яким чином здійснюється біосинтез хлорофілу b. Цей хлорофіл, який завжди є в зелених листках, імовірно, утворюється з хлорофілу а заміщенням метильної групи СН3 біля другого пірольного кільця на альдегідну —СОН. Однак очевидний шлях — через окислення молекули хлорофілу а — поки що не доведено, а ніякого іншого шляху досі не знайдено.
Доведено, що швидкість синтезу та розпадання молекули хлорофілу приблизно однакова, тому його пул постійно оновлюється в хлоропласті. Нагадаємо, що в 1960 p. P. Вудворд і М. Штрель штучно синтезували молекули хлорофілу.
Важливе значення для синтезу хлорофілів має мінеральне живлення рослин. Для цього передусім необхідна достатня кількість заліза — каталізатора синтезу хлорофілів. Залізо необхідне для синтезу амінолевулінової кислоти та протопорфіринів. Для синтезу порфіринів має бути нормальне забезпечення азотом і магнієм, тому що обидва ці елементи входять до складу хлорофілу. Також потрібні йони міді, що забезпечують утворення стійких комплексів між хлорофілом і білком.
Біосинтез каротиноїдів розпочинається з ацетил-КоА через ме-валонову кислоту, гераніл-гераніолпірофосфат до лікопіну, який є попередником усіх інших каротиноїдів (рис. 12).
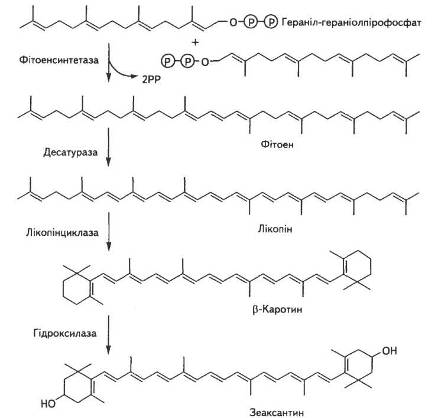
Рис. 12. Біосинтез каротиноїдів
Синтез їх відбувається в темряві, хоча різко прискорюється на світлі. Механізм біосинтезу фікобілінпротеїнів майже не вивчено.
Дата: 2019-05-29, просмотров: 382.