М¢яка (T.aestivum L.) і тверда (T.durum Dest) пшениці є поліплоїдними видами.
Сьогодні більшість дослідників вважають, що не менше п¢яти диплоїдних видів двох родів Triticum L. i Aegilops L. брали участь в становленні м¢якої пшениці: два геноми 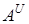 i
i 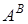 –належать дикорослим диплоїдним видам пшениці – T.urartu i T.boeoticum, інші три геноми B, D i G - роду Aegilops. Останній багатший, ніж рід Triticum L, з диплоїдами, носіями геномів. Існує ряд диплоїдних видів егілопса, геноми яких не залучені до синтезу пшеничних рослин. [9.74]
–належать дикорослим диплоїдним видам пшениці – T.urartu i T.boeoticum, інші три геноми B, D i G - роду Aegilops. Останній багатший, ніж рід Triticum L, з диплоїдами, носіями геномів. Існує ряд диплоїдних видів егілопса, геноми яких не залучені до синтезу пшеничних рослин. [9.74]
В різні історичні періоди в географічно розділених регіонах від різних материнських і батьківських форм виникли види секції Dicoccoides i Timopheevii, що мають відповідні геноми 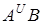 I
I 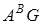 .
.
Якщо розглядати походження окремих геномів поліплоїдної пшениці на рівні сучасних знань, то донором першого геному твердої і м¢якої пшениці, а також інших поліплоїдних видів, які є їм родичами, вважають предка сучасного T.urartu, ядерний геном якого позначається символом 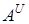 , а донором геному В-предка сучасного Ae.longissima або близьку до нього форму.
, а донором геному В-предка сучасного Ae.longissima або близьку до нього форму.
Таким чином, первинний тетраплоїд секції Dicoccoides виник при схрещуванні диплоїдних видів –дикої пшениці і егілопса – шляхом з¢єднання їх ядерних геномів. Донором цитоплазми твердої пшениці міг бути донор геному В - Ae.longissima або, за іншими джерелами, Ae.bicornis. Але повної генетичної відповідності цитоплазми цих егілопсів і цитоплазми твердої пшениці немає: ці відмінності могли виникнути в результаті філогенетичного розвитку цих видів пшениці і егілопса за остані десятки тисяч років в порівнянні з їх первинними формами.[6.63]
Запитання про донора компоненту гексаплоїдної пшениці дискутувалося з 20-х років ХХ –ст (Культурная флора СССР.Пшеница.-Л.,1979,-Т.1). В наш час вважають, що компонент отриманий гексаплоїдною пшеницею від первиного T.persicum Vav.
Донором геному D гексаплоїдної пшениці, що забезпечив її перехід з тетраплоїдного двогеномного рівня на трьох геномний, був диплоїдний вид Ae.tauchii Coss (=Ae.squarrosa L.) subsp strangulata (Eig.) Tzvel.
Первинний T.persicum спонтанно схрестився з Ae.tauchii subsp strangulata; в ареалі останього виникла перша гексаплоїдна пшениця. Геном D в поєднанні з двома іншими геномами забезпечив їй більшу внутрівидову різноманітність, розширення ареалу, завдяки підвищеній пристосованості до умов середовища, появи озимих форм пшениці під впливом діяльності людини, збільшення зимо- і морозостійкості, підвищення хлібопекарських якостей, але зниження вмісту білка в зерні, зниження стійкості до грибкових захворювань і отримання летальних генів.
Донором цитоплазми видів секції Triticum був вихідний для них тетраплоїдний вид, який в свою чергу отримав її від донора геному В.
У видів пшениць секції Timopheevii. донором першого геному ( 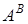 ) була дикоросла однозернянка T.boeoticum.Boiss, а донором геному G-Ae.speltoides.Таким чином, види секції Timopheevii виникли при гібридизаційному об¢єднанні геномів T.boeoticum iAe.speltoides.[1.36]
) була дикоросла однозернянка T.boeoticum.Boiss, а донором геному G-Ae.speltoides.Таким чином, види секції Timopheevii виникли при гібридизаційному об¢єднанні геномів T.boeoticum iAe.speltoides.[1.36]
Походження твердої і м¢якої пшениці (І) і видів секції Timopheevii (ІІ) представлені в схемі 1.
Генетичний потенціал господарсько цінних ознак при внутрівидових (міжсортових) схрещуваннях має певні межі. Потрібні пошуки нових методів збагачення генотипу м¢якої пшениці новими корисними ознаками. Один з таких методів – віддалена і міжвидова гібридизація.
Відомо, що багато видів пшениці не мають виробничого значення, але містять значний резерв корисних генів.
Ознайомлення з ботанічним складом пшениці (додаток. 1) показує, що більшість диких видів (T.boeoticum, T.urartu, T.araraticum, T.dicoccoides) і культурних плівчастих (T.monococcum, T.macha, T.zhukovskyi і ін.) практично не залучено до селекції. Слабко залучались до схрещення древні плівчасті види, що мали цінні для сучасної селекції ознаки (високий вміст білка в зерні і лізину в білку, скороспілість, комплексна стійкість до грибкових хвороб, посухостійкість, холодостійкість і ін.).
Схема 1
Походження твердої і м ¢ якої пшениці (І) і видів секції Timopheevii (ІІ)
І ♂ Прототрітікум (T.urartu)*♀В Протоегілопс (Ae.longissima або Ae.bicornic)
 |

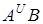 ♀ * ♂ D
♀ * ♂ D
тверда пшениця первиний Ae.tauchi subsp.
(T.durum) T.persicum strangulata

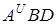
м¢яка пшениця
(T.aestivum)
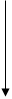 II ♂
II ♂ 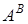 * ♀G
* ♀G
T.boeoticum Ae.speltoides
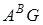
види пшениць секції
Timopheevii
Мало використовуються в схрещуваннях види T.timopheevii і його голозерний аналог – новий T.militinae, що мають високий імунітет; імунні до борошнистої роси холодостійкий вид T.persicum; стійкий до хвороб штучно створений октоплоїдний вид T. fungicidum і ін.[14.132]
При міжвидових схрещуваннях селекціонер зазвичай отримує генетично збагачену гібридну популяцію, де є лінії з новими трансгресивними ознаками, яких не має у вихідних форм.
У ВІРі проведена велика робота по міжвидовій гібридизації пшениці. Вона була розпочата ще М.І.Вавіловим. Накопичений великий матеріал по схрещуванню видів, і їх генетичної сумісності, наслідуванню корисних для селекції ознак різних видів пшениці і можливості їх селективного використання.
Найбільш широко поширеною на земній кулі і основною хлібною культурою є пшениця м¢яка. Ареал поширення охоплює п’ять континентів нашої планети: від північних широт до південних берегів Африки і Америки; вирощується на землях, які розміщені нижче рівня світового океану, і піднімається до висоти чотири тисячі метрів (в горах Перу). Все це свідчить про пластичність м¢якої пшениці. В нашій країні вирощується як основна зернова культура.
За способом життя ділиться на озимі і ярі форми, напівозимі і пізні ярі, а також двуручки. З морфолого-екологічними ознаками К.А.Фляксберг (1935) поділив м¢яку пшеницю на два підвиди: ірано-азіатські (колос товстого типу) та індоєвропейський колос ніжного типу. [20.145]
Пшениця м¢яка нараховує більше ста різновидностей (Дорофеєв і ін.). Ряд з них (Erytrospermum, lutescens, ferruginerum, graecum, albidium, milturum.). Дуже пластичні і поширені по всіх контенентах земної кулі. Найбільш багатий різновидностями м¢якої ярої пшениці азіатський континент і країни СНД, що підтверджує наявність тут центрів походження цього виду; найменше різновидностей в Австралії та Північній Америці. Найбільше сортів популяції нараховується в країнах СНД та на азіатському континенті.
М¢яка пшениця – ведуча продовольча культура багатьох країн світу. В країнах СНД вона є головною хлібною культурою. Сьогодні на цьому виді зосереджена основна увага селекціонерів пшениці. Її генoтип дозволяє створювати сорти, що відповідають потребам інтенсивного землеробства.
Сорти м¢якої пшениці по комплексу господарсько цінних ознак і біологічних властивостей дуже різноманітні. М¢яка пшениця генетично не сумісна з T.boeoticum I T.monococcum. Гібридизація м¢якої пшениці, яка відноситься до гексаплоїдних голозерних видів, з тетраплоїдними видами з геномами 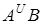 свідчить про їх часткову генетичну сумісність.
свідчить про їх часткову генетичну сумісність.
Гібридизація м¢якої пшениці з гексаплоїдними видами з геномами 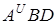 . Вдається легко, так як ці види генетично сумісні.
. Вдається легко, так як ці види генетично сумісні.
Генетична сумісність відсутня у випадку схрещування м¢якої пшениці з видами, що мають геном G.
Перед селекціонерами стоять задачі всесвітнього підвищення продуктивності і якості зерна м¢якої пшениці. Прагнучи покращити господарські ознаки селекціонери все частіше звертаються до схрещування м¢якої пшениці з іншими видами. Особливо яскравий приклад використання твердої пшениці для покращення м¢якої (сорти Саратовська 29 та ін.).
В наш час для селекційного покращення м’якої пшениці застосовується велика кількість сортів різного походження. Часто міжвидове або внутрівидове схрещування буває безрезультатним через наслідки, які викликаються гіброїдним некрозом, або хлорозом. Списки сортів – носіїв того чи іншого гена некрозу чи хлорозу – публікуються. Виявилось, що гени гібридного некрозу мають значне поширення А.C. Zeven(1971) повідомив, що з 2513 сортів м’якої пшениці гени 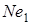 або
або 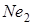 були в 49,7 % м’якої пшениці (А.А.Мкртичан і Т.А.Минасян, 1973) підтвердили ці дані Н.С.Саркисян і ін. (1971) визначили, що типово ярі пшениці мають ген
були в 49,7 % м’якої пшениці (А.А.Мкртичан і Т.А.Минасян, 1973) підтвердили ці дані Н.С.Саркисян і ін. (1971) визначили, що типово ярі пшениці мають ген 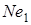 , а справжні озимі
, а справжні озимі 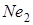 .[3.82]
.[3.82]
Дослідники при схрещуваннях часто зіштовхувалися з гібридною карликовістю. М.Танака (1961)вказав на існування двох домінантних генів, що викликають це явище- 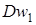 і
і 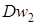 повідомив, що гібридна карликовість обумовлена взаємодією трьох домінантних генів
повідомив, що гібридна карликовість обумовлена взаємодією трьох домінантних генів 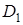
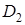 ,
, 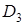 що діють комплементарно. Один домінантний ген не може дати ефекту гібридної карликовості. Наявність великої кількості таких генів пов¢язано з різким зниженням продуктивності гібридів і часто з їх летальністю. Ген
що діють комплементарно. Один домінантний ген не може дати ефекту гібридної карликовості. Наявність великої кількості таких генів пов¢язано з різким зниженням продуктивності гібридів і часто з їх летальністю. Ген 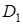 локалізований в хромосомі 2 Д, знайдений в сорті м’якої пшениці з Південної Європи, Африки, Азії ген , локалізований в хромосомі 2 В, зустрічається в сорті з різних континентів; ген локалізований в хромосомі 4 В, визначений лише в сорті Європи (Zeven A.C, 1970).
локалізований в хромосомі 2 Д, знайдений в сорті м’якої пшениці з Південної Європи, Африки, Азії ген , локалізований в хромосомі 2 В, зустрічається в сорті з різних континентів; ген локалізований в хромосомі 4 В, визначений лише в сорті Європи (Zeven A.C, 1970).
Розглянемо результати використання міжвидової гібридизації в селекції м’якої пшениці.
Міжвидова гібридизація як метод використовується багатьма селекціонерами. На ній основані роботи по ЦМС м’якої пшениці; по переводі на озимий спосіб життя і підвищення зимостійкості твердої пшениці; по збільшені кількості білку в зерні; по імунітету до грибкових хвороб, короткостебельності і т.д.
М.І.Вавілов (1935) для покращення харчових властивостей м’якої пшениці пропонував використовувати в схрещуваннях спельту і звичайну полбу. Д.Костов (1932) рекомендував використовувати метод потрійних видових гібридів з метою подолання бар¢єру несхрещуваності між м¢якою пшеницею і T.monococcum носієм генів стійкості до хвороб. Цей метод пропонує наступний хід схрещування (T.dicoccum *T.Monococcum) *T.aestivum.
Д.Циков (1964) виявив, що схрещування м’якої пшениці з кулезерною перспективне, так як дає гібриди з високою якістю зерна і скороспілістю. Крім цих якостей, така гібридизація дозволяє отримати рослини з міцною соломиною, посухостійкістю, морозостійкістю. Появу у таких видів гібридів ознаки морозостійкості помітив ще В.Р.Каришнєв (1950).[16.157]
Виконання програми селекції стійких до хвороб сортів пов¢язане з використанням в схрещуваннях T.timopheevii як генетичного джерела імунітету. Генетичний бар¢єр несхрещуваності між пшеницями Тимофєєва і м¢якою намагалися подолати шляхом створення амфідиплоїдів (Жебрак А.Р. 1957 ; Таврин Е.В. 1963, і ін.). Отриманні амфідиплоїди розщеплюються по стійкості до бурої іржі.
Але поєднувати імунітет пшениці Тимофєєва з генотипом м’якої пшениці важко. В.О.Гулканяк (1947), А Kiss. (1956), Д.Циков (1961), і інші не досягли їх об¢єднання. З іншого боку, не можна відкидати результати отриманні іншими дослідниками R.G.Shands (1941,1954),R.W.Allard (1954) отримали цитологічні стабільні лінії м’якої пшениці, що мають високу стійкість до стеблової і бурої іржі борошнистої роси, пильної головні як у проростків, так і в дорослих рослин. Ці лінії мали транслокацію сегментів хромосом від пшениці Тимофєєва. П.М.Жуковський (1957), Н.А.Скуригіна (1970) і інші вважали, що пшениця Тимофєєва може бути використана як донор генів стійкості до грибкових хвороб при селекційному покращенні м’якої пшениці. Н.А.Скуригіна (1970) помітила, що стійкі до хвороб родини м¢якої пшениці появилися приблизно в 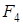 , імунність до грибкових захворювань помічена лише в гібриді, що мають фенотип пшениці Тимофєєва; гібридних рослин проміжного типу і типом м’якої пшениці, спельти, пшениці Вавілова помічена більша або менша схильність до хвороб. При цьому В.Л.Менабде (1946, 1948), Н.А.Скуригіна (1963), помітили в
, імунність до грибкових захворювань помічена лише в гібриді, що мають фенотип пшениці Тимофєєва; гібридних рослин проміжного типу і типом м’якої пшениці, спельти, пшениці Вавілова помічена більша або менша схильність до хвороб. При цьому В.Л.Менабде (1946, 1948), Н.А.Скуригіна (1963), помітили в  майже повне зникнення ознак пшениці Тимофєєва. В наступній роботі (1970) Н.А.Скуригіна отримала згадані стійкі до грибкових захворювань лінії м’якої пшениці, які не поєднували цю якість з опушеністю листків і вузлів, характерні для пшениці Тимофєєва. В досліді Е.Ф.Мігушової по схрещуванню з видами, що мають геном G., встановлений тісний зв¢язок між імунітетом рослин і щетинистою опушеністю вузлів і листків, що, можливо, виражається в формі зчеплення генів.[19.148]
майже повне зникнення ознак пшениці Тимофєєва. В наступній роботі (1970) Н.А.Скуригіна отримала згадані стійкі до грибкових захворювань лінії м’якої пшениці, які не поєднували цю якість з опушеністю листків і вузлів, характерні для пшениці Тимофєєва. В досліді Е.Ф.Мігушової по схрещуванню з видами, що мають геном G., встановлений тісний зв¢язок між імунітетом рослин і щетинистою опушеністю вузлів і листків, що, можливо, виражається в формі зчеплення генів.[19.148]
При міжвидовій гібридизації м’якої пшениці зазвичай використовують насичуюче схрещування, які вдаються досить легко.
Використання видів пшениці в практичній селекції пов¢язане з затриманням форм, які легко обмолочуються.
Культурні види пшениці мають дві різні генетичні системи, що забезпечують легку обмолочуваність зерна. Легкий вимолот на гексаплоїдному рівні пов¢язаний з наявністю фактору Q., який мають T. Aestivum, T.sphorococum I T.compactum (Мак Кейл; 1968), а також T.petropavlovskiy При схрещуванні цих видів з плівчастими і голозерними тетраплоїдними видами (F1- строге самозапилення) в F2 і наступних поколіннях, вищеплялась м¢яка пшениця як і при схрещуванні з T.persicum (дослід Е.Ф.Мігушової).[11.176]
Дата: 2019-05-29, просмотров: 371.