Пшениця – основна хлібна культура на землі. В результаті розселення її в різні еколого-географічні райони внаслідок активної селективної діяльності людини світовий генофонд пшениці став широким і різноманітним. Селекціонер для вдалого підбору вихідних форм при схрещуванні повинен вільно орієнтуватись в систематиці роду (додаток 1).
В сучасній тритікології отримала визнання гіпотеза про дифілетичне походження тетраплоїдної пшениці. В різний історичний час в різних географічних районах Близького Сходу від різних материнських і батьківських видів в природі на територіальному рівні виникли два первинних амфідиплоїди, що відрізняються по генному складу в цитоплазмі. Ними були дикорослі полби T.dicoccoides (геноми 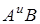 ) і T. Araraticum (геноми
) і T. Araraticum (геноми 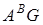 )В нашому розумінні вони були першими поліплоїдними видами двох підродів основними генами яких є
)В нашому розумінні вони були першими поліплоїдними видами двох підродів основними генами яких є 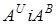 Поєднання цих геномів з трьома геномами (B,G,D) роду Aegilops дало поліплоїдні пшениці.
Поєднання цих геномів з трьома геномами (B,G,D) роду Aegilops дало поліплоїдні пшениці.
В залежності від генного складу види в підродах групуються в 6 секцій. Філогенетичний розвиток роду йшов як по шляху появи видів гомологів в двох гомологічних підродах, так і по шляху становлення видів аналогів всередині кожного з цих підродів. Види гомологи мають різний геномний склад, одинаковий рівень плоїдності і схожі господарські особливості, які контролюються головними генами. Наприклад, полбі T.dicoccum гомологічна полба Т.timopheevii: вони мають різний геном ний склад,2n=28, важкий вимолот зерна і ламкий стержень колоса. Види-аналоги мають одинаків геномний склад, один і той же рівень плоїдності і різні господарські властивості, що контролюються головними генами. Наприклад, T.persicum - легкообмолочуваний аналог полби T dicoccum: вони мають одинаків геномний склад, 2n=28, але в першому випадку вимолот зерна легкий, стержень колоса міцний, в іншому – вимолот зерна важкий, а стержень ламкий.[15.56]
Види підродів Triticum мають 3 рівні плоїдності (2n=14,2n=28,2n=42) і є одно -, дво - і трьохгеномними.
За господарським значенням види з геномами 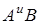 I
I 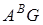 діляться на полби і голозерні тетраплоїди, види з геномами - на спельти і голозерні гексаплоїди. В обох підродах дикорослі види є як серед однозернянок, так і серед полб. Серед гексаплоїдів дикорослих видів немає.
діляться на полби і голозерні тетраплоїди, види з геномами - на спельти і голозерні гексаплоїди. В обох підродах дикорослі види є як серед однозернянок, так і серед полб. Серед гексаплоїдів дикорослих видів немає.
Дикорослі види секцій Urartu ( 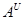 ) I Monococcon (
) I Monococcon ( 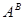 ) мають плоский колос (бічна сторона його ширша за лицеву), ламкий стержень, важкий вимолот зерна і одну зернівку (рідко 2) в колоску. Одомашнення торкнулось лише однозернянок з геномом . При цьому дикорослий вид T.boeoticum втратив спонтанну ламкість колосу і дуже важкий вимолот зерна. Культурна однозернянка T.monococcum, що виникла з цього, більш урожайна (в основному за рахунок деякого укрупнення зернівки і збільшення продуктивного кущіння). Кульмінація філогенетичного розвитку секції Monococcon - появлення природного мутанта T.Sinskajae - легкообмолочуваного аналога T.monococcum.
) мають плоский колос (бічна сторона його ширша за лицеву), ламкий стержень, важкий вимолот зерна і одну зернівку (рідко 2) в колоску. Одомашнення торкнулось лише однозернянок з геномом . При цьому дикорослий вид T.boeoticum втратив спонтанну ламкість колосу і дуже важкий вимолот зерна. Культурна однозернянка T.monococcum, що виникла з цього, більш урожайна (в основному за рахунок деякого укрупнення зернівки і збільшення продуктивного кущіння). Кульмінація філогенетичного розвитку секції Monococcon - появлення природного мутанта T.Sinskajae - легкообмолочуваного аналога T.monococcum.
Види секції Dicoccoides ( B) пройшли шлях розвитку від дикої полби до культурної і потім до голозерних тетраплоїдів. Всі полби зазвичай мають плоский колос з ламким стержнем, важким вимолотом зерна і двома зернівками в колоску. Одомашнення дикорослої полби Т. dicoccoides йшло по лінії втрати колосом спонтанної ламкості і дуже сильного одеревіння колоскової луски. Всі культурні полби мають менш важкий, ніж у дикорослої полби, вимолот зерна, а колосся їх ламаються лише при натискуванні. Подальший процес окультурення тетраплоїдної пшениці йшов по шляху отримання легкого вимолоту зерна, міцного колосового стержня і збільшення кількості зернівок в колоску. Це привело до появи колосів, в яких лицева сторона дорівнює або ширша від бічної. Втрата сильного одеревіння колосковими лусками, що відповідає за важкий обмолот, пов’язана, по J. Мас Кеy (1968), з мутаційним проявом фактора Q (T.persicum) і з становленням полігенної системи легкого обмолоту (T.durum і близькі до нього види).
В секції Timopheevii ( 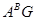 ) процес одомашнення йшов шляхом, схожим з гомологічною секцією Diccocoides від дикорослої полби T.araraticum до культурної полби T.timopheevii. Філогенетичний хід розвитку цієї секції закінчився появою в наші дні голозерного аналога T.timopheevii якому було дано назву T.militinae. В останнього виду легкий обмолот зерна забезпечений фактором
) процес одомашнення йшов шляхом, схожим з гомологічною секцією Diccocoides від дикорослої полби T.araraticum до культурної полби T.timopheevii. Філогенетичний хід розвитку цієї секції закінчився появою в наші дні голозерного аналога T.timopheevii якому було дано назву T.militinae. В останнього виду легкий обмолот зерна забезпечений фактором 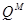 який виник внаслідок мутації. Його дія подібна до дії фактора Q в T.persicum. Полігенна система легкого обмолоту в тетраплоїдів- носіїв геномів
який виник внаслідок мутації. Його дія подібна до дії фактора Q в T.persicum. Полігенна система легкого обмолоту в тетраплоїдів- носіїв геномів 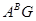 не сформувалась. Це, напевно, повязано з їх порівняно недавнім походженням із відсутністю масового виробництва, що обмежило їх селективне опрацювання.[8.34]
не сформувалась. Це, напевно, повязано з їх порівняно недавнім походженням із відсутністю масового виробництва, що обмежило їх селективне опрацювання.[8.34]
Види секції Triticum ( 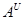 B
B 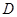 ) (виникли при спонтанному схрещуванні культурного тетраплоїда з Ae.tauschii. Якщо вихідним для них видом пшениці був T.persicum (Kihara H., Lilienfeld F., 1949), то первина гексаплоїдна форма мала легкий обмолот і достатньо міцний cтержень колоса. Гексаплоїдні види з важким обмолотом, можливо, з¢явились в результаті спонтанної гібридизації м¢якої пшениці з тетраплоїдними видами (Вавілов М.І., 1926, Сінська О.Н. 1955, Жуковський П.М, 1957), тобто вони є вториними за походженням. Слід відмітити, що плівчасті види секції Diccocoides, Monococcon, Timopheevii є первиними. Плівчасті гексаплоїдні види (спельти) мають специфічний тип ламкості : крім типу ламкості, який характерний для полб, стержень їх колосу може ламатися як в Ae.tauschii.
) (виникли при спонтанному схрещуванні культурного тетраплоїда з Ae.tauschii. Якщо вихідним для них видом пшениці був T.persicum (Kihara H., Lilienfeld F., 1949), то первина гексаплоїдна форма мала легкий обмолот і достатньо міцний cтержень колоса. Гексаплоїдні види з важким обмолотом, можливо, з¢явились в результаті спонтанної гібридизації м¢якої пшениці з тетраплоїдними видами (Вавілов М.І., 1926, Сінська О.Н. 1955, Жуковський П.М, 1957), тобто вони є вториними за походженням. Слід відмітити, що плівчасті види секції Diccocoides, Monococcon, Timopheevii є первиними. Плівчасті гексаплоїдні види (спельти) мають специфічний тип ламкості : крім типу ламкості, який характерний для полб, стержень їх колосу може ламатися як в Ae.tauschii.
Остання ознака особливо характерна для європейських форм Т.spelta,
В яких членик стержня (після розлому колоса) прилягає до черевної сторони колоса. Такий тип розлому пов¢язаний з наявністю геному D.
В групі спельт ламкість стрижня колоса відсутня в носія гена “вавілоїдності”-у T.vavilovii. В гексаплоїдних видів легкий обмоліт забезпечує фактор Q. В залежності від селективного опрацювання форм, - носіїв цього фактору- лицева сторона колосу може бути менша від бічної, дорівнювати їй або бути більшою за неї. Число зернівок в колоску в останньому випадку досягає 5-6.Така кількість зернівок могла розміститися в одному колоску після того, як була забезпечена можливість розсунення колоскових лусок. Останнє пов¢язане з втратою їх сильного одеревіння через дію фактору Q.
Секція Kiharae ( 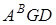 )гомологічна до секції Triticum і включає штучно синтезовані види T.kiharae I T.miguscovae.
)гомологічна до секції Triticum і включає штучно синтезовані види T.kiharae I T.miguscovae.
Kiharae ( 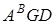 )був виділений у ВІРі В. Ф.Дорофеевим і Е.Ф. Мігушовою (1977) з амфідиплоїду Т.timopeevii, Ae.tauschii синтезованого в Японії і названий на честь японського видатного генетика Х.Кіхара. Вказаний вид є гомологом Т.spelta і має в своєму генетипі всі гени імунітету, характерні для Т. timopeevii, Т..miguscovae ( ) -перший в світі гомолог м¢якої пшениці був синтезований в Краснодарським НДІСГ Є.Г.Жировим (1980) при схрещуванні Т.militinae і А е.tauschii і названий на честь тритіколога Е.Ф.Мігушової. Створення імуного гомолога м¢якої пшениці відкриває нові шляхи в селекції роду Triticum[5.89]
)був виділений у ВІРі В. Ф.Дорофеевим і Е.Ф. Мігушовою (1977) з амфідиплоїду Т.timopeevii, Ae.tauschii синтезованого в Японії і названий на честь японського видатного генетика Х.Кіхара. Вказаний вид є гомологом Т.spelta і має в своєму генетипі всі гени імунітету, характерні для Т. timopeevii, Т..miguscovae ( ) -перший в світі гомолог м¢якої пшениці був синтезований в Краснодарським НДІСГ Є.Г.Жировим (1980) при схрещуванні Т.militinae і А е.tauschii і названий на честь тритіколога Е.Ф.Мігушової. Створення імуного гомолога м¢якої пшениці відкриває нові шляхи в селекції роду Triticum[5.89]
В 1940р. болгарський генетик Д.Костов отримав гексаплоїдний амфідиплоїд від схрещування Т.monococcum I Т.timopheevii, назвав його T.timococcum Kost (геноми ( 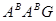 ).
).
А.Р.Жебрак 1944. схрестив Т.aestivum з Т. monococcum і отримав гексаплоїдний амфідиплоїд T.edvardii Zhebrak геномним складом 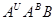 . Відомо що рід Triticum на октоплоїдному рівні не еволюціонував: в природі немає октоплоїдних видів пшениці. Вони створені експериментально людиною. Вперше октоплоїдні види з одинаковими геномами -
. Відомо що рід Triticum на октоплоїдному рівні не еволюціонував: в природі немає октоплоїдних видів пшениці. Вони створені експериментально людиною. Вперше октоплоїдні види з одинаковими геномами - 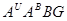 отримали П.М.Жуковський (1944) - Т.fungicidum Zhuk. (пшениця грибкобійна) від схрещування Т.persicum і Т.timopheevii з наступною поліплоїдизацією колхіцином-А.Р.Жебрак(1944)-Т.soveticum Zhebrak від схрещування Т.aestivumз Т.timopheevii.
отримали П.М.Жуковський (1944) - Т.fungicidum Zhuk. (пшениця грибкобійна) від схрещування Т.persicum і Т.timopheevii з наступною поліплоїдизацією колхіцином-А.Р.Жебрак(1944)-Т.soveticum Zhebrak від схрещування Т.aestivumз Т.timopheevii.
Згодом в1959р. Французький ботанік H.Heslot oтримав і описав разом з R.Ferrari октоплоїдний вид T.timonovum Heslot et Ferrari як автополплоїд Т.timopheevii. (геноми 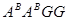 )[17.64]
)[17.64]
В 1981р. Співробітник ВІРа Н.Наврузбеков експериментально отримав октоплоїдну пшеницю від схрещування двох голозерних тетраплоїдних видів-Т.militinae і Т. Persicum, назвав свій амфідиплоїд в честь видатного тритіколога К.А. Фляксберга - Т. flaksbergeri Navr (геноми 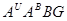 ). Ці види найбільш полігеномні- мають чотири різні геноми
). Ці види найбільш полігеномні- мають чотири різні геноми 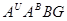 .
.
Експериментально отримані види пшениці не мають практичного значення вони не пройшли адаптацію в природі і на полях, але вони можуть використовуватись як вихідний матеріал.
Дата: 2019-05-29, просмотров: 342.