Быстрый прогресс в области вирусологических знаний, основанный в значительной мере на достижениях смежных естественных наук, обусловил возможность углубленного познания природы вирусов. Как ни в одной другой науке, в вирусологии прослеживается быстрая и четкая смена уровней познания — от уровня организма до субмолекулярного.
Приведенные периоды развития вирусологии отражают те уровни, которые являлись доминирующими в течение одного — двух десятилетий.
Уровень организма (30-40 гг. XX века). Основной экспериментальной моделью являются лабораторные животные (белые мыши, крысы, кролики, хомяки, обезьяны и т. д.), основным первым модельным вирусом был вирус гриппа.
В 40-е годы в вирусологию в качестве экспериментальной модели прочно входят куриные эмбрионы. Они обладали высокой чувствительностью к вирусам гриппа, оспы и некоторым другим. Использование этой модели стало возможным благодаря исследованиям австралийского вирусолога и иммунолога Ф. Бернета, автора первого пособия по вирусологии «Вирус как организм». В 1960 г. Ф. Бернет и П. Медавар удостоены Нобелевской премии в области вирусологии.
Открытие в 1941 г. американским вирусологом Херстом феномена гемагглютинации немало способствовало изучению взаимодействия вируса с клеткой на модели вируса гриппа и эритроцитов.
Большим вкладом отечественных вирусологов в медицинскую вирусологию явилось изучение природно-очаговых заболеваний — эпидемических энцефалитов. В 1937 г. была организована первая экспедиция, возглавляемая Зильбером, в составе которой были Левкович, Шубладзе, Чумаков, Соловьев и др. Благодаря проведенным исследованиям был открыт вирус клещевого энцефалита, выявлены его переносчики — иксодовые клещи, разработаны методы лабораторной диагностики, профилактики и лечения. Советскими вирусологами были изучены вирусные геморрагические лихорадки, разработаны препараты для диагностических и лечебно-профилактических целей.
Уровень клетки (40-50 гг. XX века). В 1949 г. происходит значительное событие в истории вирусологии — открытие возможности культивировать клетки в искусственных условиях. В 1952 г. Дж. Эндерс, Т. Уэллер, Ф. Роббинс получили Нобелевскую премию за разработку метода культуры клеток. Использование культуры клеток в вирусологии явилось подлинно революционным событием, послужившим основой для выделения многочисленных новых вирусов, их идентификации, клонирования, изучения их взаимодействия с клеткой. Появилась возможность получения культуральных вакцин. Эта возможность была доказана на примере вакцины против полиомиелита. В содружестве с американскими вирусологами Дж. Солком и А. Сэбином, советскими вирусологами М.П. Чумаковым, А.А. Смородинцевым и др. была разработана технология производства, апробирована и внедрена в практику убитая и живая вакцины против полиомиелита. В 1959 г. была проведена массовая иммунизация детского населения в СССР (около 15 млн.) живой полиомиелитной вакциной, в результате резко снизилась заболеваемость полиомиелитом и практически исчезли паралитические формы заболевания. В 1963 г. за разработку и внедрение в практику живой полиомиелитной вакцины М.П. Чумакову и А.А. Смородинцеву была присуждена Ленинская премия. В 1988 г. ВОЗ приняла решение о глобальной ликвидации заболеваемости полиомиелитом. В России это заболевание не регистрируется с 2002 года.
Другим важным приложением техники выращивания вирусов явилось получение Дж. Эндерсом и Смородинцевым живой коревой вакцины, широкое применение которой обусловило значительное снижение заболеваемости корью и является основой для искоренения этой инфекции.
Широко внедрялись в практику и другие культуральные вакцины — энцефалитная, ящурная, антирабическая и т. д.
Молекулярный уровень (50-60 гг. XX века). В вирусологии широко стали использовать методы молекулярной биологии, а вирусы благодаря простой организации их генома стали распространенной моделью для молекулярной биологии. Ни одно открытие молекулярной биологии не обходится без вирусной модели, включая генетический код, всего механизма внутриклеточной экспрессии генома, репликации ДНК, процессинга (созревания) информационных РНК и т. д.
В свою очередь использование молекулярных методов в вирусологии позволило установить принципы строения (архитектуры) вирусных индивидуумов — вирионов, способы проникновения вирусов в клетку и их репродукции.
Субмолекулярный уровень (70-80 гг. XX века). Стремительное развитие молекулярной биологии открывает возможности изучения первичной структуры нуклеиновых кислот и белков. Появляются методы секвенирования ДНК, определения аминокислотных последовательностей белка. Получают первые генетические карты геномов ДНК-содержащих вирусов.
В 1970 г. Д. Балтимором и одновременно Г. Теминым и С. Мизутани была открыта обратная транскриптаза в составе РНК-содержащих онкогенных вирусов, фермент, переписывающий РНК на ДНК. Становится реальным синтез гена с помощью этого фермента на матрице, выделенной из полисом иРНК. Появляется возможность переписать РНК в ДНК и провести ее секвенирование.
В 1972 г. возникает новый раздел молекулярной биологии — генная инженерия. В этом году публикуется сообщение П. Берга в США о создании рекомбинантной молекулы ДНК, которое положило начало эре генной инженерии. Появляется возможность получения большого количества нуклеиновых кислот и белков путем введения рекомбинантных ДНК в состав генома прокариот и простых эукариот. Одним из основных практических приложений нового метода является получение дешевых препаратов белков, имеющих значение в медицине (инсулин, интерферон) и сельском хозяйстве (дешевые белковые корма для скота).
Этот период характеризуется важными открытиями в области медицинской вирусологии. В фокусе изучения — три наиболее массовых болезни, наносящих огромный ущерб здоровью людей и народному хозяйству — грипп, рак, гепатит.
Установлены причины регулярно повторяющихся пандемий гриппа. Детально изучены вирусы рака животных (птиц, грызунов), установлена структура их генома и идентифицирован ген, ответственный за злокачественную трансформацию клеток — онкоген. Установлено, что причиной гепатитов А и В являются разные вирусы: гепатит А вызывает РНК-содержащий вирус, отнесенный к семейству пикорнавирусов, а гепатит В — ДНК-содержащий вирус, отнесенный к семейству гепаднавирусов. В 1976 г. Бламберг, исследуя антигены крови у аборигенов Австралии, обнаружил так называемый австралийский антиген, который он принял за один из антигенов крови. Позже было выявлено, что этот антиген является антигеном гепатита В, носительство которого распространено во всех странах мира. За открытие австралийского антигена Бламбергу в 1976 г. была присуждена Нобелевская премия.
Другая Нобелевская премия в 1976 г. присуждена американскому ученому К. Гайдушеку, который установил вирусную этиологию одной из медленных инфекций человека — куру, наблюдающейся в одном из туземных племен на острове Новая Гвинея и связанной с ритуальным обрядом — поеданием зараженного мозга умерших родственников.
Начиная со второй половины 80-х годов вирусологи активно включились в разработку неожиданно возникшей в мире проблемы ВИЧ-инфекции. Этому способствовал значительный опыт работы отечественных ученых с ретровирусами.
Медицинская микробиология, вирусология и иммунология во многом обязаны исследованиям отечественным ученым таким как Н.Ф. Гамалея (1859-1949), П.Ф. Здродовский (1890-1976), Л.А. Зильбер (1894-1966), Д.И. Ивановский (1864-1920), Л.А. Тарасевич (1869-1927), В.Д. Тимаков (1904-1977), Е.И. Марциновский (1874-1934), В.М. Жданов (1914-1987), З.В. Ермольева (1898-1979), А.А. Смородинцев (1901-1989), М.П. Чумаков (1909-1990), П.Н. Кашкин (1902-1991), Б.П. Первушин (1895-1961) и многих других.
НАУЧНЫЕ ВИРУСОЛОГИЧЕСКИЕ УЧРЕЖДЕНИЯ
Первые вирусологические лаборатории в нашей стране были созданы в 30-е годы: в 1930 г. — лаборатория по изучению вирусов растений в Украинском институте защиты растений, в 1935 г. — отдел вирусов в Институте микробиологии АН СССР, а в 1938 г. он был реорганизован в отдел вирусов растений, которым в течение многих лет руководил В.Л. Рыжков. В 1935 г. была организована Центральная вирусологическая лаборатория Наркомздрава РСФСР в Москве, которой заведовал Л.А. Зильбер, а в 1938 г. эта лаборатория реорганизована в отдел вирусов Всесоюзного института экспериментальной медицины, его руководителем был назначен А.А. Смородинцев. В 1946 г. на базе отдела вирусов был создан Институт вирусологии АМН СССР, которому в 1950 г. присвоено имя Д.И. Ивановского.
В течение 50-х и 60-х годов созданы научные и производственные вирусологические учреждения в нашей стране: Институт полиомиелита и вирусных энцефалитов АМН СССР, Институт вирусных препаратов Министерства здравоохранения СССР, Киевский институт инфекционных болезней, Всесоюзный научно-исследовательский институт гриппа Министерства здравоохранения СССР в Ленинграде и ряд других.
Важную роль в подготовке кадров вирусологов сыграла организация в 1955 г. кафедры вирусологии в Центральном институте усовершенствования врачей МЗ СССР. Кафедры вирусологии были созданы на биологических факультетах Московского и Киевского университетов.
ГЛАВА 2. ХИМИЧЕСКИЙ СОСТАВ ВИРУСОВ
Просто организованные вирусы представляют собой нуклеопротеиды или нуклеокапсиды и состоят из нуклеиновой кислоты (РНК или ДНК) и нескольких кодируемых ею белков, формирующих вирусную оболочку вокруг нуклеиновой кислоты — капсид.
Рис. 2. Просто устроенные вирусы (без оболочки).
Сложно организованные вирусы содержат дополнительные оболочки, белковые или липопротеидные, и имеют более сложный химический состав. Помимо нуклеиновой кислоты и белков, они содержат липиды в наружных оболочках и углеводы в составе белков наружных оболочек (гликопротеидов). Обычно липиды и углеводы имеют клеточное происхождение. В составе некоторых вирусов обнаруживаются также клеточные нуклеиновые кислоты и белки.
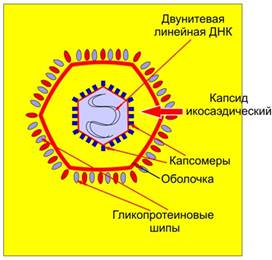
Рис. 3. Сложно устроенные вирусы (с оболочкой).
НУКЛЕИНОВЫЕ КИСЛОТЫ
Клетки всех живых организмов содержат два вида нуклеиновой кислоты — ДНК и РНК. ДНК представляет собой двунитчатую молекулу, а РНК — однонитчатую. Двунитчатая ДНК — это клеточный геном, выполняющий функции хранения и репликации наследственной информации. Однонитчатая РНК представлена 3 классами молекул: 1) информационные РНК (иРНК), образующиеся в результате транскрипции генома и передающие заложенную в геноме информацию на белок-синтезирующий аппарат клетки; 2) рибосомальные РНК, являющиеся структурным элементом рибосомы; 3) тРНК, доставляющие аминокислоты к белок-синтезирующему аппарату.
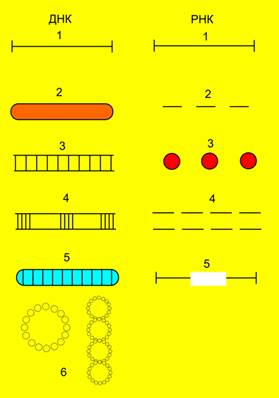
Рис. 4. Типы молекул вирусных ДНК и РНК:
ДНК: 1 - парвовирусы, 2 - фаг Х174, 3 - аденовирусы, 4 - фаг Т5, 5 - паповавирусы, 6 - вирус гепатита В;
РНК: 1- пикорнавирусы, тогавирусы, парамиксовирусы, рабдовирусы, 2 - ортомиксовирусы, 3 - буньявирусы, 4 - реовирусы, 5 - ретровирусы.
В отличие от клеток вирусы содержат лишь один вид нуклеиновой кислоты — либо РНК, либо ДНК. И та, и другая может быть хранителем наследственной информации, выполняя, таким образом, функции генома.
Вирусные нуклеиновые кислоты характеризуются поразительным разнообразием форм. Вирусный геном может быть представлен как однонитчатыми, так и двунитчатыми молекулами РНК и ДНК. ДНК может быть как линейной, так и кольцевой молекулой, РНК — как непрерывной, так и фрагментированной и кольцевой молекулой (рис. 4, табл. 1, 2).
Таблица 1.
Типы молекул вирусных ДНК
| Вирусы | Тип ДНК |
| Парвовирусы | Линейная однонитчатая |
| Фаг Х174 и другие фаги | Кольцевая однонитчатая |
| Аденовирусы, вирусы герпеса | Линейная двунитчатая |
| фаг Т5, Т7 и другие фаги | Линейная двунитчатая с разрывами в одной цепи |
| Вирусы оспы | Двунитчатая с замкнутыми кон цами |
| Паповавирусы, фаг RM2, вирус мозаики цветной капусты | Двунитчатая кольцевая со сверх витками или без них |
| Вирус гепатита В | Двунитчатая кольцевая с однонитчатым участком |
Таблица 2.
Типы молекул вирусных РНК
| Вирусы | Тип РНК |
| Пикорнавирусы, тогавирусы, парамиксовирусы, рабдовирусы | Линейная однонитчатая |
| Ортомиксовирусы, аренавирусы, вирус мозаики, костра | Фрагментированная однонитчатая |
| Буньявирусы | Фрагментированная однонитчатая кольцевая |
| Реовирусы, вирус раневых опухолей растений, вирус цитоплазматического полиэдроза насекомых | Фрагментированная двунитчатая |
| Ретровирусы | Линейная однонитчатая |
ВИРУСНЫЕ ДНК
Молекулярная масса вирусных ДНК варьирует в широких пределах от 1·106 до 250·106 (табл. 3). Самые большие вирусные геномы содержат несколько сотен генов, а самые маленькие содержат информацию, достаточную для синтеза лишь нескольких белков.
В геномах, представленных двунитчатыми ДНК, информация обычно закодирована на обеих нитях ДНК. Это свидетельствует о максимальной экономии генетического материала у вирусов, что является неотъемлемым свойством их как генетических паразитов. В связи с этим — оценка генетической информации не может быть проведена по молекулярной массе молекул.
Таблица 3.
Дата: 2019-04-23, просмотров: 490.