Влияние и роль этой структуры весьма велико. Одни группы нейронов формации оказывают влияние на двигательный потенциал и вегетативные структуры спинного мозга, реализуя это влияние через нисходящие пути.
Другие ядра, особенно те, которые находятся в области мезенцефалона, или среднего мозга, имеют проекции в thalamus, а также «имеют свои представительства» на уровне субталамических ядер,systemnuclei subthalamici. Эти структуры таламуса имеют мощные коллатеральные волокна от разных волокон, которые являются восходящими. Например, ретикулярная формация таламуса тесно связана со спинномозговым путем тройничного нерва, спинно-бугорным пучком, с nucleus tracti solitarii, или ядром одиночного пути. В промежуточном мозге ретикулярная формация также формирует пути связи с зрительной, обонятельной системой, а также с вестибулярными и улитковыми ядрами.
Такая анатомия, в принципе, дает ответ на вопрос, почему наш сон прекращается при появлении яркого солнца, или звоне будильника. Активирующая роль сети резко возрастает при получении специфической информации.
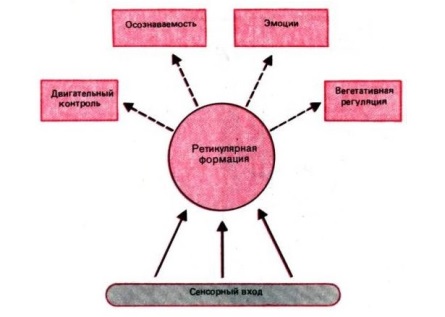
В настоящее время принято считать, что функции ретикулярной формации состоят в следующем:
- она поддерживает определенный уровень сознания и внимания;
- уровень внимания может регулироваться, вплоть до настороженности;
- регулирует уровни сна и бодрствования.
Несмотря на то, что строение ретикулярной формации «рассыпное», при поражении ее какой-либо области, например, при нарушении мозгового кровообращения в области ствола, часто наблюдаются различные нарушения сознания, вплоть до развития коматозного состояния.
Нисходящие пути ретикулярной сети – это латеральный и вентральный ретикулоспинальные пучки – оказывают разное влияние на мотонейроны спинного мозга. Информация, исходящая от этой структуры, может быть, как активирующей, так и ингибирующей.
Такая сложная организация возможна потому, что на саму ретикулярную формацию оказывает большое влияние кора. Роль коры очень велика, особенно в таких участках, как лобные зоны коры, мозжечок и базальные ганглии. Анатомия этих структур такова, что организация связей с формацией в этих участках самая плотная.
Латеральная (боковая часть) сетчатой структуры, которая находится в варолиевом мосту и средней части мозга, оказывает активизирующую деятельность, соответственно по ретикуло-спинномозговым и вестибуло-спинномозговым путям.
Кроме регуляции внимания и бодрствования, формация играет значительную роль в поддержании автоматического (бессознательного) мышечного тонуса, который необходим для стояния, ходьбы. Его нарушение приводит к невозможности перемещаться. Если разрушена анатомия поддержания равновесия спинномозговых рефлекторных дуг, тогда отдельные части рефлексов будут преобладать над другими, что вызовет общую дискоординацию движения.
Вегетативный отдел
Заслуживает особого внимания и та часть ретикулярной формации, которая поддерживает многие функции «растительной» жизни. Эти нейроны рассеяны по всему продолговатому мозгу и мосту, и функционально они тесно взаимосвязаны с вегетативными ядрами некоторых черепно-мозговых нервов.
Эти нейроны ретикулярной структуры получают импульсы из таламуса и вышележащих отделов. Многие из этих нейронов находятся в гипоталамусе. Такое обилие источников импульсов из таламуса объясняется тем, что именно гипоталамус является высшим центром вегетативной регуляции.
Так, саливация, или слюноотделение, в норме контролируется двумя ядрами: верхним и нижним слюноотделительными. При виде пищи, ее запахе и вкусе, а также при виде накрытого стола человек обостряет свое внимание к пище, и происходит рефлекторная наработка слюны. Но если голова человека «занята» сильными эмоциями, он находится в состоянии нервного стресса, то у него возникает сухость во рту. Точно так же, если человек спит, то даже если он спит на кухне, то слюна у него не вырабатывается.
Это показывает контролирующую работу ретикулярной формации в отношении вегетативных отделов.
- Другие нейроны контролируют работу сердечно-сосудистой системы. Так, есть опосредованная регуляция кровяного давления через ретикулярную формацию. Эту связь можно проследить в виде схемы:
сонный гломус, или каротидный синус, расположенный в области бифуркации сонных артерий, находит, что в крови снизилось содержание кислорода;
- восходящие импульсы об этом идут в составе блуждающего и языкоглоточного нерва в вегетативные ядра продолговатого мозга;
- окружающие нейроны ретикулярной формации посылают ингибирующее влияние в спинной мозг, которые предупреждают вазодилатацию, то есть расширение сосудов, и блокируют работу ядер вагуса.
- в результате работа сердца увеличивается, поскольку парасимпатические влияния вагуса (десятой пары черепно-мозговых нервов) тормозятся, то сердце попадает под влияние антагонистов – симпатических нервов. Это повышает давление, силу и частоту сердечных сокращений, и увеличивает фракцию выброса
Также ядра ретикулярной формации контролируют дыхание, точнее, центры вдоха и выдоха, которые находятся дорзальнее (сзади) нижних олив. Есть ядра, отвечающие за глотание. Ведь глотание – это очень сложный процесс, и все мышцы, от полости рта и до желудка должны участвовать в акте глотания синхронно, но каждая в своё время. Поэтому центр глотания, как и рвотный центр, также контролируется ретикулярной формацией.
Никогда не бывает, что человек испытывает рвоту во сне. Он обязательно проснется, а потом его вырвет. Этот механизм предусмотрен природой, как профилактика от захлёбывания рвотными массами. Рвота может возникнуть в бессознательном состоянии, у пациентов с тяжелым инсультом, или у алкоголиков, но в норме ретикулярная формация обязательно «разбудит» человека, получив импульс о приближении рвоты и тошноты.
В заключение нужно сказать, что ретикулярная формация связана с таким большим количеством структур, что главные факты о ее роли в работе центральной нервной системы, по всей видимости, еще впереди. Имеются данные о связи этой структуры с эмоциями, и даже с процессами абстрактного (математического) мышления.
Время покажет.
3.2 Пирамидная система, пирамидный путь
(лат. tractus pyramidalis) — система нервных структур. Поддерживает сложную и тонкую координацию движений.
Пирамидная система — одно из поздних приобретений эволюции. Низшие позвоночные пирамидальной системы не имеют, она появляется только у млекопитающих, и достигает наибольшего развития у обезьян и, особенно, у человека. Пирамидная система играет особую роль в прямохождении
Кора полушарий головного мозга в V слое содержит клетки Беца (или гигантские пирамидные клетки)
Пирамидный путь осуществляется нервными волокнами, которые исходят от клеток Беца и спускаются в спинной мозг, не прерываясь. Волокна перекрещиваются на границе головного и спинного мозга (большая часть — в продолговатом мозге, меньшая — в спинном). Далее они проходят через спинной мозг (передние и боковые столбы спинного мозга). Импульсы от коры головного мозга эти волокна передают либо непосредственно, либо через вставочные нейроны.
Непосредственное раздражение определенных участков коры головного мозга приводит к судорогам мышц, соответствующих участку коры — проекционной двигательной зоне *(при раздражении верхней трети передней центральной извилины возникает судорога мышц ноги, средней — руки, нижней — лица, причем, на стороне, противоположной очагу раздражения в полушарии). Эти судороги носят название парциальных (джексоновских). Их открыл английский невролог Д. Х. Джексон (1835—1911). В проекционной двигательной зоне каждого полушария головного мозга представлены все мышцы противоположной половины тела.
Пирамидная система человека содержит около 1 млн нервных волокон. Различают следующие типы волокон:
| Тип нервных волокон | Диаметр | Скорость проведения | Функция |
| Толстые, быстропроводящие | 16 мкм | до 80 м/с | обеспечивают быстрые фазные движения |
| Тонкие, медленнопроводящие | 4 мкм | от 25 до 7 м/с | отвечают за тоническое состояние мышц |
Наибольшее количество пирамидных клеток (клеток Беца) иннервирует мелкие мышцы, отвечающие за тонкие дифференцированные движения кисти, мимику и речевой акт. Значительно меньшее их количество иннервирует мышцы туловища и нижних конечностей
В зависимости от локализации патологического процесса различают следующие проявления:
| Локализация патологического процесса пирамидного пути | Симптомы |
| проекционные зоны коры головного мозга | Центральный (спастический) паралич (или парез) т.е. -полная (неполная) утрата произвольных движений (из-за повышеного тонуса мышц – дрожание). (нет нарушения питания конечностей – гипо- и атрофии) |
| в области внутренней капсулы | гемиплегия (верхних или нижних конечност, моноплегия - одной) паралич руки и ноги на стороне, противоположной локализации очага. |
| в области ствола мозга | Альтернирующие синдромы — сочетание гемиплегии на стороне, противоположной очагу, с признаками нарушения функций черепно-мозгового нерва на стороне поражения. |
| в спинном мозге | Гемиплегия или паралич ноги на стороне повреждения — перекрест волокон остался выше. |
(Лечат основное заболевание + физич нагрузки)
Экстрапирамидная система
(лат. extra — вне, снаружи, в стороне + pyramis греч. — пирамида) — совокупность структур (образований) головного мозга, участвующих в управлении движениями, поддержании мышечного тонуса и позы, минуя кортикоспинальную (пирамидную) систему. Структура расположена в больших полушариях и стволе головного мозга
Экстрапирамидные проводящие пути образованы нисходящими проекционными нервными волокнами, по происхождению не относящимися к гигантским пирамидным клеткам (клеткам Беца) коры больших полушарий мозга. Эти нервные волокна обеспечивают связи нейронов подкорковых структур (мозжечок, базальные ядра, ствол мозга) головного мозга со всеми отделами нервной системы, расположенными дистальнее.
Частью экстрапирамидной системы является стриопаллидарная система, которая объединяет ядра полосатого тела и их афферентные и эфферентные пути. В этой системе выделяют филогенетически новую часть — стриатум, к которой относятся хвостатое ядро и скорлупа чечевицеобразного ядра, и филогенетически старую часть — паллидум (бледный шар). Стриатум и паллидум различаются по своей нейроархитектонике(композиции), связям и функциям.
(Стриатум получает волокна из коры большого мозга, центрального ядра таламуса и чёрного вещества. Эфферентные волокна из стриатума направляются в паллидум, а также в чёрное вещество. Из паллидума волокна идут в таламус, гипоталамус, к субталамическому ядру и в ствол головного мозга. Последние образуют чечевицеобразную петлю и частично оканчиваются в ретикулярной формации, частично идут к красному ядру преддверным и оливным ядрам) Следующее звено экстрапирамидных путей составляют ретикулярно-спинномозговой, красноядерно-спинномозговой, преддверно-спинномозговой и оливоспинномозговой пути, оканчивающиеся в передних столбах и промежуточном сером веществе спинного мозга. Мозжечок включается в экстрапирамидную систему посредством путей, соединяющих его с таламусом, красным ядром и оливными ядрами.
Функционально экстрапирамидная система неотделима от пирамидной системы. Она обеспечивает упорядоченный ход произвольных движений, регулируемых пирамидной системой; регулирует врожденные и приобретённые автоматические двигательные акты, обеспечивает установку мышечного тонуса и поддержание равновесия тела; регулирует сопутствующие движения (например движения рук при ходьбе) и выразительные движения (мимика).
Экстрапирамидная система состоит из следующих структур головного мозга:
- базальные ганглии,
- красное ядро (подкорковый центр экстропир системы, участв в координ движ, наруш вызыв артельнирующие параличи (синдромы Клода, Бенидикта),
- интерстициальное ядро,
- тектум (верхнее двуххолмие – зрит центр - контроль движения глаз и др. функции) ,
- чёрная субстанция(подкорковый центр экстропир системы),
- ретикулярная формация моста и продолговатого мозга,
- Ядра вестибулярного аппарата (преддверия –части внутр уха, орган, воспринимающей изменения положения головы и тела в пространстве и направление движения тела)
- мозжечок (равновесие и мышечный тонус)
- премоторная область коры
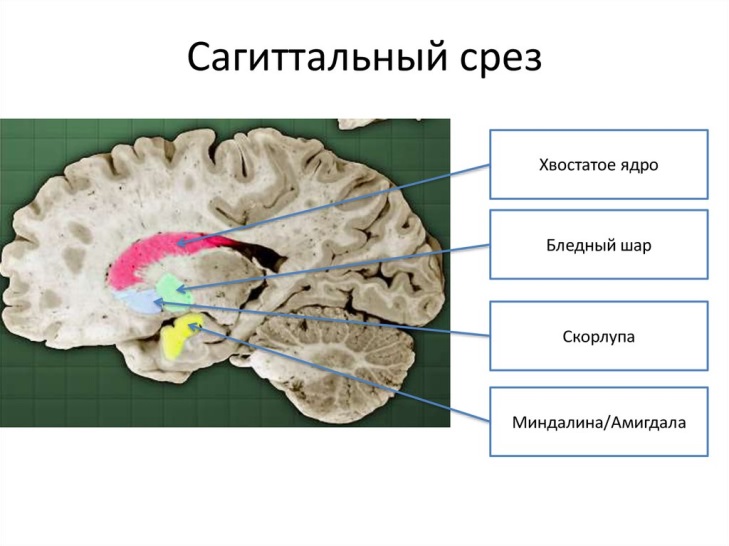
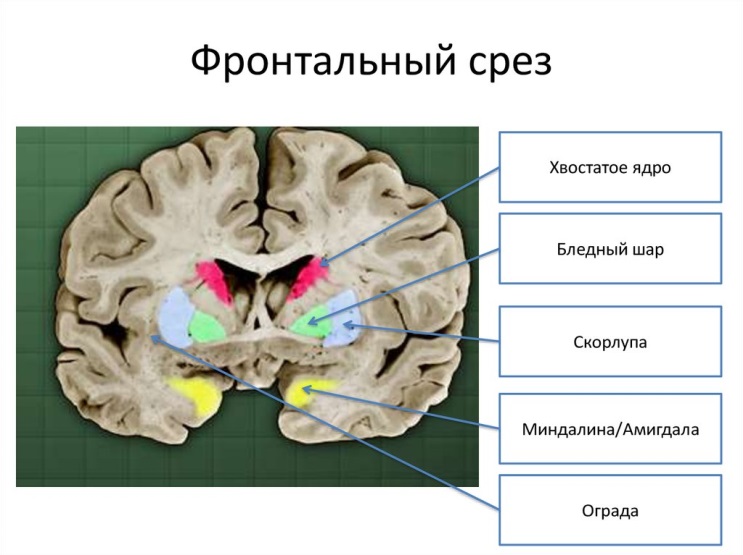
База́льные я́дра (также база́льные га́нглии, лат. nuclei basales) — несколько скоплений серого вещества, расположенных в белом веществе латеральнее таламуса на уровне основания полушарий конечного мозга. Базальные ядра входят в состав переднего мозга, расположенного на границе между лобными долями и над стволом мозга. Согласно топографии анатомического среза мозга, выделяют: 1.Полосатое тело (лат. corpus striatum) (чечевицеобразное ядро (лат. nucleus lentiformis)(скорлупа (лат. putamen) + бледный шар (лат. globus pallidus))+хвостатого ядро (лат. nucleus caudatus)-парная структура) 2. ограда (лат. claustrum) (функции ограды остаются недостаточно изученными, структуры относят к лимбической системе) 3.миндалевидное тело (лат. corpus amygdaloideum) (структуры миндалевидного тела относят к лимбической системе). 4.внутреннюю капсулу (лат. capsula interna - белое вещество между таламусом и чечевицеобразным ядром), 5.наружную капсулу (лат. capsula externa - между чечевицеобразным ядром и оградой) и 6.самую наружную капсулу (лат. capsula extrema - между оградой и островком)
Однако в последнее время, чаще используют функциональноую топографию, где под термином «базальные ядра» понимают: 1.полосатое тело 2. несколько ядер промежуточного и среднего мозга (2.1 субталамическое ядро (лат. nucleus subtalamicus), 2.2 чёрная субстанция (лат. substantia nigra) и 2.3 ножкомостовое ядро покрышки (лат. nucleus tegmentalis peduncolopontinus)), которые совместно обеспечивают функциональную регуляцию движений и мотивационных аспектов поведения.
Чёрная субстанция, также чёрное вещество (лат. Substantia nigra)(нейромеланин) находится в области четверохолмия среднего мозга. Играет важную роль в регуляции моторной функции, тонуса мышц, во многих вегетативных функциях: дыхании, сердечной деятельности, тонусе кровеносных сосудов Впервые обнаружена французским анатомом и врачом Феликсом Вик-д'Азиром в 1784 году. Представляет собой непрерывную полосу в срезах среднего мозга, анатомические исследования показали, что на самом деле, она состоит из двух частей с очень различными связями и функциями: pars compacta и pars reticulata. Данная классификация была впервые предложена Сано в 1910 году[3]. Pars compacta служит в основном в качестве приёмника сигналов — в цепи базальных ганглиев, поставляя дофамин полосатому телу. Pars reticulata служит в основном в качестве трансмиттера (передатчика), передавая сигналы от базальных ганглиев к другим многочисленным структурам головного мозга. Ядро pars compacta отвечает за обеспечение синтеза дофамина. Чёрная субстанция является важнейшей составной частью дофаминергической системы награды. Она также играет очень большую роль в мотивации и эмоциональной регуляции материнского поведения.
Играет значимую роль в развитии многих заболеваний, включая болезнь Паркинсона . (в чёрной субстанции расположены тела нейронов, аксоны которых, составляющие нигростриарный путь, который оканчиваются в неостриатуме в виде широкого сплетения терминальных микровезикул с высоким содержанием дофамина, именно этот путь является тем местом в мозге, поражение которого приводит к формированию синдрома паркинсонизма).
Приложение 1.*Зоны коры головного мозга
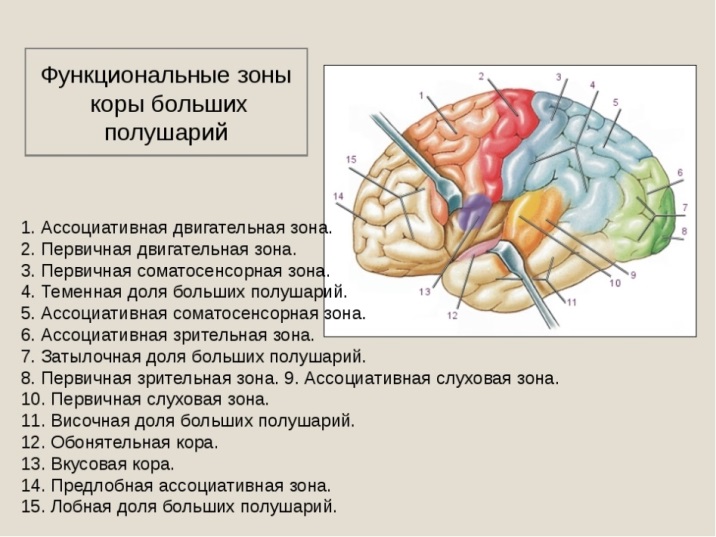
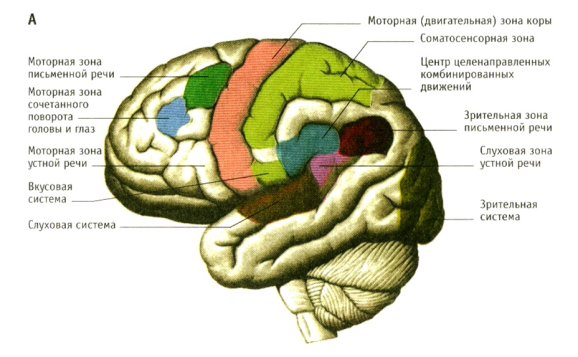
Различают ядерные зоны (корковые концы анализаторов) и ассоциативные зоны .
- первичные сенсорные зоны и первичные моторные зоны (проекционные зоны);
- вторичные сенсорные зоны и вторичные моторные зоны (ассоциативные одномодальные зоны);
- третичные зоны (ассоциативные разномодальные зоны);
Первичные сенсорные и моторные зоны занимают менее 10% поверхности коры головного мозга и обеспечивают наиболее простые сенсорные и двигательные функции
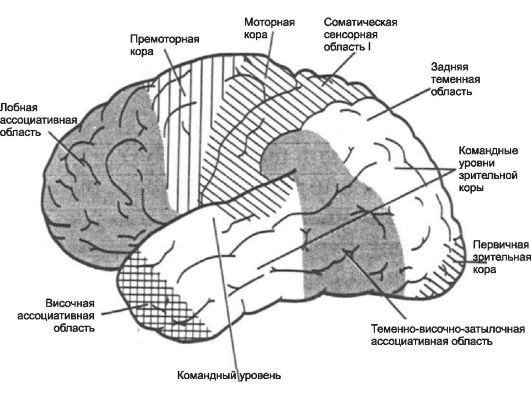
.
Ядерные зоны
Сенсорные зоны - это функциональные зоны коры головного мозга , которые через восходящие нервные пути получают сенсорную информацию от большинства рецепторов тела. Они занимают отдельные участки коры, связанные с определенными видами ощущений. Размеры этих зон коррелируют с числом рецепторов в соответствующей сенсорной системе.
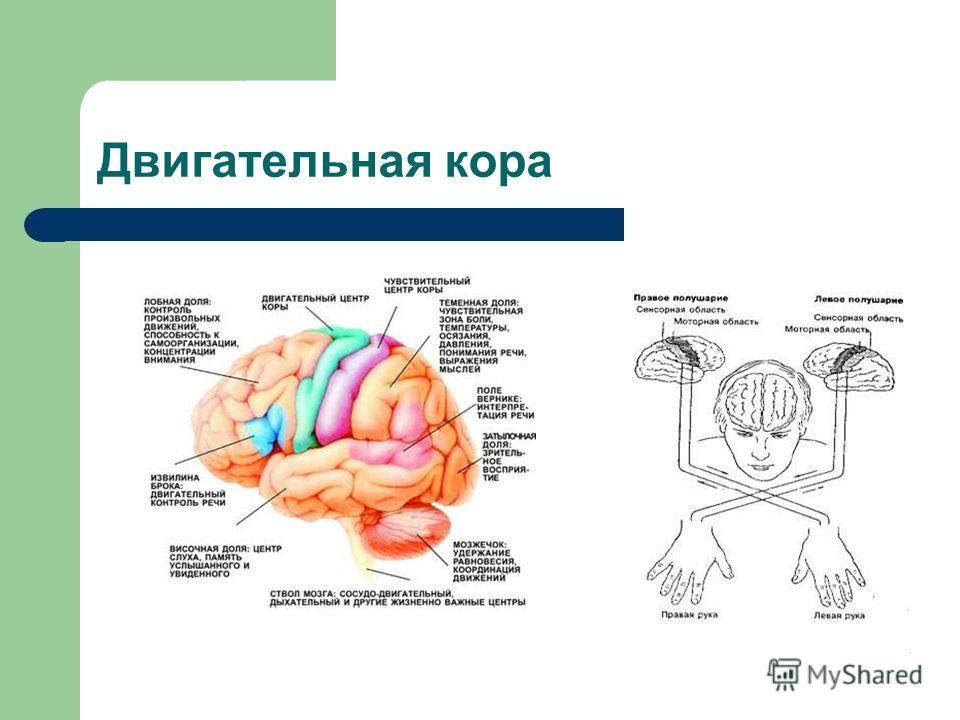
Двигательные зоны коры - это функциональные зоны коры головного мозга , посылающие двигательные импульсы к произвольным мышцам по нисходящим путям, которые начинаются в белом веществе больших полушарий . Многие двигательные импульсы идут прямо в спинной мозг через два больших пирамидных тракта ( кортикоспинальных тракта ), проходящих в стволе мозга . Остальные двигательные импульсы передаются по экстрапирамидным путям , здесь же идут двигательные импульсы от базальных ганглиев и мозжечка . В продолговатом мозге все пути перекрещиваются, так что импульсы, идущие от коры левого полушария , иннервируют правую половину тела и наоборот. Тела нейронов, участвующих в образовании пирамидных трактов, лежат в двигательных зонах коры, а их аксоны образуют синапсы непосредственно с мотонейронами спинного мозга в том его сегменте, где эти нейроны выходят на периферию.
Пути. В головном мозге нет никаких промежуточных синапсов, поэтому импульсы и последующие ответы на них по пути не задерживаются и не видоизменяются. Главным экстрапирамидным трактом является ретикулоспинальный тракт , переключающий импульсы от ретикулярной формации , которая лежит в стволе мозга между таламусом и продолговатым мозгом (из различных отделов головного мозга, контролирующих двигательную активность, импульсы поступают в определенные участки ретикулярной формации, где они модифицируются под воздействием импульсов, идущих от коры, и становятся либо возбуждающими, либо тормозными. (импульсы от мозжечка и премоторной зоны коры , управляющей координированными движениями, поступают в ту область ретикулярной формации, которая находится в продолговатом мозгу и посылает импульсы, стимулирующие тормозные мотонейроны . Последние подавляют активацию определенных мышц, что дает возможность осуществлять сложные координированные движения тела, другие комбинации двигательных импульсов,стимулируют возбуждающие нейроны, и общее воздействие ретикулярной формации на двигательную активность оказывается.возбуждающим). Большинство волокон сенсорных нейронов на своем пути через таламус к коре отдает коллатерали (боковые ветви) в ретикулярную формацию, участвуя в образовании ретикулярной активирующей системы , которая тонизирует кору и участвует в пробуждении организма от сна . Недостаточная активность этой системы или ее разрушение приводит соответственно к глубокому сну или коме . Как полагают, многие вещества, вызывающие общий наркоз , оказывают свое действие, временно блокируя синаптическую передачу в этой системе. Предполагается так же, что ретикулярная активирующая система ответственна за возникновение и поддержание побуждений к действию и концентрации внимания .
Ассоциативные зоны
На долю корковых центров приходится лишь небольшая площадь коры больших полушарий, преобладают участки, непосредственно не выполняющие чувствительные и двигательные функции. Эти области называются ассоциативными. Они обеспечивают связи между различными центрами, участвуют в восприятии и обработке сигналов, объединении получаемой информации с информацией, заложенной в памяти. Современные исследования позволяют считать, что в ассоциативной коре расположены чувствительные центры высшего порядка . Речь и мышление человека осуществляются при участии всей коры полушарий большого мозга. Ассоциативные зоны связывают вновь поступающую сенсорную информацию с полученной ранее и хранящейся в блоках памяти, а также сравнивают между собой информацию, получаемую от разных рецепторов. Сенсорные сигналы интерпретируются, осмысливаются и при необходимости используются для определения наиболее подходящих ответных реакций, которые выбираются в ассоциативной зоне и передаются в связанную с ней двигательную зону, участвуют в процессах запоминания , учения и мышления, и результаты их деятельности составляют то, что обычно называют интеллектом. Отдельные крупные ассоциативные области расположены в коре рядом с соответствующими сенсорными зонами (зрительная ассоциативная зона расположена в затылочной зоне непосредственно впереди сенсорной зрительной зоны и осуществляет описанные выше ассоциативные функции , связанные со зрительными ощущениями). Некоторые ассоциативные зоны выполняют лишь ограниченную специализированную функцию и связаны с другим ассоциативными центрами, способными подвергать информацию дальнейшей обработке. (звуковая ассоциативная зона анализирует звуки , разделяя их на категории, а затем передает сигналы в более специализированные зоны, такие как речевая ассоциативная зона , где воспринимается смысл услышанных слов)
Эти зоны относятся к ассоциативной коре и участвуют в организации когнитивных функций (познавательных ) и сложных форм поведения .
- вторичные сенсорные зоны и вторичные моторные зоны (ассоциативные одномодальные зоны);
- третичные зоны (ассоциативные разномодальные зоны);
- паралимбические зоны и
- лимбические зоны
Функциональные зоны коры головного мозга и соответствующие им цитоархитектонические поля по Бродману (52 поля, отличаются по клеточному строению)
Первичные сенсорные и моторные
Зрительная (поле 17, или стриарная кора)
Слуховая (поля 41, 42)
Соматосенсорная (поля 3, 1, 2, в основном поле Зb)
Моторная (поле 4)
Вторичные сенсорные и моторные
Зрительная [поля 18—19, 20—21, 37 (?)]
Слуховая (поле 22)
Соматосенсорная (поле 5, передний отдел поля 7
Премоторная [поле 6, задний отдел поля 8 (?), поле 44 (?)]
Третичные
Префронтальная (поля 9, 10, 45, 46, 47, передние отделы полей 11, 12,32)
Теменно-височная [поля 39, 40, задний отдел поля 7, задний край верхней височной борозды, поле 36 (?)]
Паралимбические
Островок (поля 14, 15), височный полюс (поле 38), задние отделы орбитофронтальной коры (поля 11, 12), поясная извилина [поля 23, 24, 31 (?), 33, 25, 26, 29, задний отдел поля 32], пара-гиппокампальная извилина (поля 28, 34, 35, 30)
Лимбические
Подкорковые структуры (миндалевидное тело, безымянная субстанция, ядра прозрачной перегородки)
Аллокортекс (гиппокамп, периформная кора)
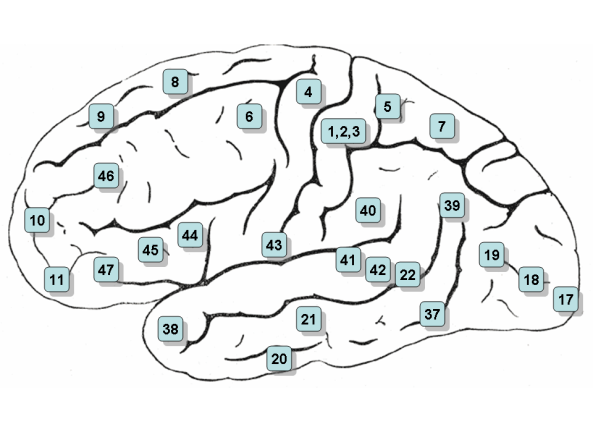
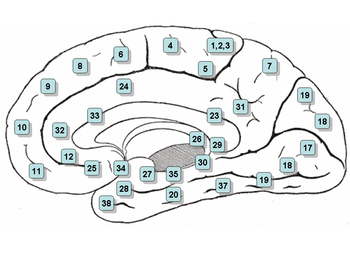
Первичные и вторичные поля (ядерная зона анализатора) получают импульсы непосредственно от таламуса, в то время как третичные — только от первичных и вторичных полей. Первичные поля производят специфический анализ импульсов определенной модальности. Вторичные поля осуществляют взаимодействие различных анализаторных зон. Третичные поля играют определяющую роль в сложных видах психической деятельности — символической, речевой, интеллектуальной
Поля 3, 1 и 2 — соматосенсорная область, первичная зона. Находятся в постцентральной извилине. В связи с общностью функций используется термин «поля 3, 1 и 2» (спереди назад)
Поле 4 — моторная область. Располагается в пределах прецентральной извилины
Поле 5 — вторичная соматосенсорная зона. Располагается в пределах верхней теменной дольки
Поле 6 — премоторная кора и дополнительная моторная кора (вторичная моторная зона). Располагается в передних отделах прецентральной и задних отделах верхней и средней лобной извилин.
Поле 7 — третичная зона. Расположена в верхних отделах теменной доли между постцентральной извилиной и затылочной долей
Поле 8 — располагается в задних отделах верхней и средней лобной извилин. Включает в себя центр произвольных движений глаз
Поле 9 — дорсолатеральная префронтальная кора
Поле 10 — передняя префронтальная кора
Поле 11 — обонятельная область
Поле 17 — ядерная зона зрительного анализатора — зрительная область, первичная зона
Поле 18 — ядерная зона зрительного анализатора — центр восприятия письменной речи, вторичная зона
Поле 19 — ядерная зона зрительного анализатора, вторичная зона (оценка значения увиденного)
Поле 20 — нижняя височная извилина (центр вестибулярного анализатора, распознавание сложных образов)
Поле 21 — средняя височная извилина (центр вестибулярного анализатора)
Поле 22 — ядерная зона звукового анализатора
- Поле 24 —детектор ошибок
Поле 28 — проекционные поля и ассоциативная зона обонятельной системы
- Поле 32 — дорсальная зона передней поясной коры. Рецепторная область эмоциональных переживаний.
- Поле 37 — Акустико-гностический сенсорный центр речи. Это поле контролирует трудовые процессы речью, ответственно за понимание речи. Центр распознавания лиц.
Поле 39 — ангулярная извилина, часть зоны Вернике (центр зрительного анализатора письменной речи)
Поле 40 — краевая извилина, часть зоны Вернике (двигательный анализатор сложных профессиональных, трудовых и бытовых навыков)
Поле 41 — ядерная зона звукового анализатора, первичная зона
Поле 42 — ядерная зона звукового анализатора, вторичная зона
Поле 43 — вкусовая область
Поле 44 — Центр Брока
Поле 45 — триангулярная часть поля Бродмана (музыкальный моторный центр)
Поле 46 — двигательный анализатор сочетанного поворота головы и глаз в разные стороны
Поле 47 — ядерная зона пения, речедвигательная его составляющая
- Поле 52 — ядерная зона слухового анализатора, которая отвечает за пространственное восприятие звуков и речи
Борозды и извилины латеральной поверхности полушария конечного мозга (лобная и теменная области)
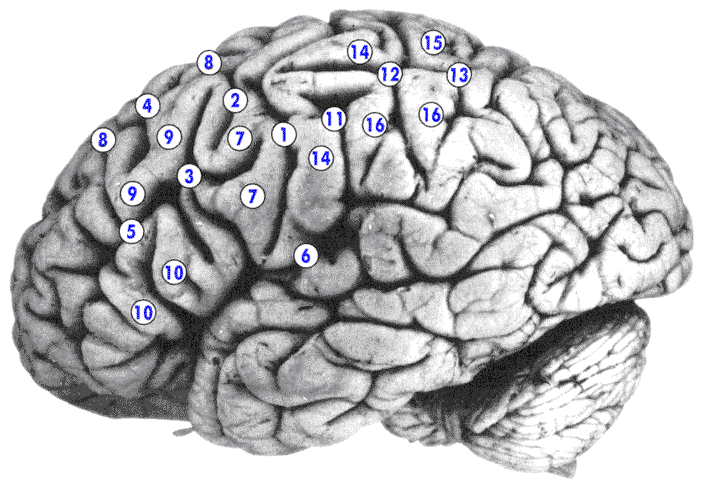
1 - центральная (роландова) борозда (sulcus centralis (Rolandi)) ;
2 - верхняя предцентральная борозда (sulcus precentralis superior) ;
3 - нижняя предцентральная борозда (sulcus precentralis inferior) ;
4 - верхняя лобная борозда (sulcus frontalis superior)
5 - нижняя лобная борозда (sulcus frontalis inferior) ;
6 - латеральная (сильвиева) борозда (sulcus lateralis (Sylvii)) ; (в глубине борозды и на внутренней поверхности верхней височной извилины находится поле слухового анализатора коры конечного мозга
7 - предцентральная извилина (gyrus precentralis) ; Ядро двигательного анализатора находится главным образом в предцентральной извилине (двигательная область коры) лобн доля конечного мозга. В предцентральной извилине находятся двигательные поля коры полушарий головного мозга (двиг. обл коры).
8 - верхняя лобная извилина (gyrus frontalis superior) (в ростральной (носовой) части верхней, средней и нижней лобной извилины располагается высшая лобная ассоциативная зона коры конечного мозга(29% коры) - участие в формировании сложной познавательной деятельности и интеллектуальной деятельности При ее поражении наблюдаются расстройства внимания, нарушаются способность к абстракциям , ориентация в пространстве и во времени , притупляются эмоции . Появляются агрессивность и немотивированность поведения , резкая смена настроения, ослабевает память);
9 - средняя лобная извилина (gyrus frontalis medius) ;
10 - нижняя лобная извилина (gyrus frontalis inferior) ;
11 - постцентральная борозда (sulcus postcentralis) ;
12 - звезда (завиток) (vortex) (место соединения постцентральной борозды и межтеменной борозды теменной доли конечного мозга);
13 - межтеменная борозда (sulcus interparietalis) ;
14 - постцентральная извилина (gyrus postcentralis) (находится между центральной бороздой и постцентральной бороздой) (в постцентральной извилине и в коре верхней теменной дольки находится поле кожного анализатора коры конечного мозга (осязательная чувствительность, болевая чувствительность и температурная чувствительность),
15 - верхняя теменная долька (lobulus parietalis superior) (лежит после «завитка») ;
16 - нижняя теменная долька (lobulus parietalis inferior)(лежит после завитка).
Дата: 2019-03-05, просмотров: 733.