Помимо циклин-зависимых киназ в регуляцию митотических событий вовлечены как минимум ещё два типа киназ: polo-подобные киназы и киназы семейства aurora. Polo-подобные киназы (англ. polo-like kinase, Plk) представляют собой серин-треониновые протеинкиназы, активирующиеся на начальных и инактивирующиеся на поздних стадиях митоза или в начале фазы G1. Данные киназы вовлечены в различные митотические процессы: сборку веретена деления, функционирование кинетохора, цитокинез. Киназы семейства aurora также относятся к группе серин-треониновых протеинкиназ. У многоклеточных выделяются два основных представителя данного семейства: aurora A и aurora B. Киназа aurora A участвует в регуляции функционирования центросом и митотического веретена. Киназа aurora B участвует в регуляции процессов конденсации и разделения сестринских хроматид, а также обеспечивает присоединение кинетохоров к микротрубочкам веретена деления.
Комплекс, стимулирующий анафазу (англ. anaphase-promoting complex, APC), также называемый циклосома, представляет собой крупное белковое соединение, которому отводится решающая роль в активации анафазы. Функционально комплекс стимуляции анафазы представляет собой убиквитинлигазу и катализирует реакции присоединения молекул убиквитина к различным целевым белкам, которые в итоге подвергаются протеолизу.
В структуре комплекса стимуляции анафазы выделяется порядка 11-13 субъединиц. Ядро комплекса составляют: субъединица куллина (Apc2) и RING-домен (Apc11), к которому присоединяется убиквитин-конъюгирующий фермент(E2). Функционирование комплекса регулируется за счёт присоединения активирующей субъединицы в нужный момент клеточного цикла.
Белок Cdc20 (англ. cell division cycle protein 20 - «белок клеточного цикла 20») активирует комплекс APC при переходе делящейся клетки из метафазы в анафазу. Происходит это следующим образом. На стадии метафазы циклин-киназный комплекс M-Cdk путём фосфорилирования трансформирует ядро комплекса APC. В результате указанного конформационного изменения повышается вероятность присоединения активатора Cdc20. В итоге, активированный комплекс APCCdc20 обретает убиквитин-лигазную активность и убиквитинирует свои главные цели - секьюрин и митотические циклины.
Секьюрин (одна из главных мишеней APCCdc20) представляет собой ингибирующий белок, сдерживающий в неактивном состоянии фермент сепаразу. Вследствие реакции убиквитинирования секьюрин разрушается, а высвободившаяся при этом сепараза разрушает когезин. После деградации когезина, обеспечивающего сцепление сестринских хроматид, происходит разделение и расхождение хромосом к полюсам деления клетки.
Убиквитинирование и, как следствие, разрушение митотических циклинов (ещё одной важной мишени APCCdc20) запускает цепочку отрицательной обратной связи. Выглядит это следующим образом. Циклин-киназный комплекс M-Cdk активирует убиквитин-лигазный комплекс APCCdc20, который целенаправленно разрушает митотические циклины, что ведёт к деградации циклин-киназного комплекса M-Cdk, то есть цепочка реакций приводит к разрушению изначального активатора этой цепочки. Но поскольку активность APCCdc20 зависит от комплекса M-Cdk, инактивация циклин-киназы M-Cdk приводит к инактивации APCCdc20. В итоге APCCdc20деактивируется к концу митоза.
Патология митоза.
Патология митоза развивается при нарушении нормального течения митотического деления и зачастую приводит к возникновению клеток с несбалансированными кариотипами, следовательно, ведёт к развитию мутаций и анеуплоидии. Также в результате развития отдельных форм патологии наблюдаются хромосомные аберрации. Незавершённые митозы, прекращающиеся по причине дезорганизации или разрушения митотического аппарата, приводят к образованию полиплоидныхклеток. Полиплоидия и формирование дву- и многоядерных клеток возникают в случае нарушений механизмов цитокинеза. При значительных последствиях патологии митоза возможна гибель клетки.
В нормальных тканях патология встречается в незначительных количествах. Например, в эпидермисе мышей встречается около 0,3% патологических митозов; в эпителии гортани и матки человека - около 2%. Патологические митозы часто наблюдаются при канцерогенезе, при различных экстремальных воздействиях, при лучевой болезни или вирусной инфекции, при раке и предраковых гиперплазиях. Частота патологических митозов также увеличивается с возрастом.
Условно различают патологию митоза функционального и органического типа. К функциональным нарушениям относят, например, гипореактивность вступающих в митоз клеток - снижение реакции на физиологические регуляторы, определяющие интенсивность пролиферации нормальных клеток. Органические нарушения возникают при повреждении структур, участвующих в митотическом делении (хромосомы, митотический аппарат, клеточная поверхность), а также при нарушении процессов, связанных с данными структурами (репликация ДНК, образование веретена деления, движение хромосом, цитокинез).
Предполагается, что сложный митотический процесс высших организмов развивался постепенно из механизмов деления прокариот. Это предположение подтверждается тем, что прокариоты появились приблизительно на миллиард лет раньше первых эукариот. Кроме того, в митозе эукариот и бинарном делении прокариот принимают участие схожие белки.
Возможные промежуточные стадии между бинарным делением и митозом можно проследить у одноклеточных эукариот, у которых в ходе деления не разрушается ядерная мембрана. У большинства же других эукариот, в том числе растений и животных, веретено деления формируется вне ядра, а ядерная оболочка разрушается в течение митоза. Хотя митоз у одноклеточных эукариот ещё недостаточно изучен, можно предположить, что он произошёл от бинарного деления и в конечном счёте достиг того уровня сложности, который имеется у многоклеточных организмов.
У многих простейших эукариот митоз также остался процессом, связанным с мембраной, однако теперь уже не плазматической, а ядерной. Возможно, в связи с увеличением размера и числа хромосом, структура типа мезосомы разделилась на два элемента: ЦОМТ на ядерной оболочке и кинетохор на хромосоме. Для соединения данных структур между собой в процессе эволюции развилась промежуточная система микротрубочек. В рамках данного представления, наиболее древним и примитивным считается закрытый внутриядерный плевромитоз. Сегрегация хромосом при этом происходит путём расхождения ЦОМТ, к которым хромосомы крепятся посредством микротрубочек. В свою очередь, ЦОМТ закреплены на ядерной оболочке и расходятся за счёт роста ядерной мембраны между ними.
От разных вариантов закрытого внутриядерного плевромитоза, вероятно, берут своё начало несколько параллельных эволюционных линий. В качестве эволюционно прогрессивных признаков при этом рассматриваются: распад ядерной оболочки во время митоза; переход ЦОМТ из ядра в цитоплазму; образование биполярного веретена; усиление спирализации хромосом; формирование экваториальной пластинки в метафазе. Таким образом, эволюция митотического деления идёт в направлении от закрытого внутриядерного плевромитоза к открытому ортомитозу.
Эндомитоз представляет собой разновидность митоза без деления ядра или клетки, в результате чего в клетке накапливается множество копий одних и тех же хромосом, собранных в одном ядре. Этот процесс может также включать эндоредупликацию, а клетки в этом случае называются эндоплоидными. Примером клеток, подвергающихся эндомитозу, могут служить мегакариоциты, дающие начало тромбоцитам.
Крайним случаем эндомитоза является образование гигантских политенных хромосом, появляющихся как результат многократного воспроизведения хромосом без последующего расхождения. Такие хромосомы встречаются в слюнных железах некоторых насекомых, у личинок двукрылых в ядрах клеток кишечника и у некоторых растений в ядрах синергид (например, гороха) .
Митоз происходит в следующих случаях:
· Рост и развитие. Количество клеток в организме в процессе роста увеличивается благодаря митозу. Это лежит в развитии многоклеточного организма из единственной клетки - зиготы, а также роста многоклеточного организма.
· Перемещение клеток. В некоторых органах организма, например, коже и пищеварительном тракте, клетки постоянно отшелушиваются и заменяются новыми. Новые клетки образуются путём митоза, а потому являются точными копиями своих предшественников. Схожим путём происходит замена красных кровяных клеток - эритроцитов, имеющих короткую продолжительность жизни - 4 месяца, а новые эритроциты формируются путём митоза.
· Регенерация. Некоторые организмы способны восстанавливать утраченные части тела. В этих случаях образование новых клеток часто идёт путём митоза. Например, благодаря митозу морские звёзды восстанавливают утраченные лучи.
· Бесполое размножение. Некоторые организмы образуют генетически идентичное потомство путём бесполого размножения. Например, гидры размножаются бесполым способом при помощи почкования. Поверхностные клетки гидры подвергаются митозу и образуют скопления клеток, называемые почками. Митоз продолжается и в клетках почки, и она вырастает во взрослую особь. Сходное клеточное деление происходит при вегетативном размножении растений.
Лекция 16. Мейоз.
Мейо́з (от др.-греч. - уменьшение), или редукционное деление, клетки - деление ядра эукариотической клетки с уменьшением числа хромосом в два раза. Происходит в два этапа (редукционный и эквационный этапы мейоза). Мейоз происходит в половых клетках и связан с образованием гамет.
С уменьшением числа хромосом в результате мейоза в жизненном цикле происходит переход от диплоидной фазы к гаплоидной. Восстановление плоидности (переход от гаплоидной фазы к диплоидной) происходит в результате полового процесса.
В связи с тем, что в профазе первого, редукционного, этапа происходит попарное слияние (конъюгация) гомологичных хромосом, правильное протекание мейоза возможно только в диплоидных клетках или в чётных полиплоидах (тетра-, гексаплоидных и т. п. клетках). Мейоз может происходить и в нечётных полиплоидах (три-, пентаплоидных и т. п. клетках), но в них, из-за невозможности обеспечить попарное слияние хромосом в профазе I, расхождение хромосом происходит с нарушениями, которые ставят под угрозу жизнеспособность клетки или развивающегося из неё многоклеточного гаплоидного организма.
Этот же механизм лежит в основе стерильности межвидовых гибридов. Поскольку у межвидовых гибридов в ядре клеток сочетаются хромосомы родителей, относящихся к различным видам, хромосомы обычно не могут вступить в конъюгацию. Это приводит к нарушениям в расхождении хромосом при мейозе и, в конечном счете, к нежизнеспособности половых клеток, или гамет (основным средством борьбы с этой проблемой является применение полиплоидных хромосомных наборов, поскольку в данном случае каждая хромосома конъюгирует с соответствующей хромосомой своего набора). Определённые ограничения на конъюгацию хромосом накладывают и хромосомные перестройки (масштабные делеции, дупликации, инверсии или транслокации).
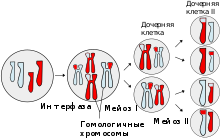
Фазы мейоза.
Мейоз состоит из 2 последовательных делений с короткой интерфазой между ними.
· Профаза I - профаза первого деления очень сложная и состоит из 5 стадий:
· Лептотена, или лептонема - упаковка хромосом, конденсация ДНК с образованием хромосом в виде тонких нитей (хромосомы укорачиваются).
· Зиготена, или зигонема - происходит конъюгация - соединение гомологичных хромосом с образованием структур, состоящих из двух соединённых хромосом, называемых тетрадами или бивалентами и их дальнейшая компактизация.
· Пахитена, или пахинема - (самая длительная стадия) - в некоторых местах гомологичные хромосомы плотно соединяются, образуя хиазмы. В них происходит кроссинговер - обмен участками между гомологичными хромосомами.
· Диплотена, или диплонема - происходит частичная деконденсация хромосом, при этом часть генома может работать, происходят процессы транскрипции (образование РНК), трансляции (синтез белка); гомологичные хромосомы остаются соединёнными между собой. У некоторых животных в ооцитах хромосомы на этой стадии профазы мейоза приобретают характерную форму хромосом типа ламповых щёток.
· Диакинез - ДНК снова максимально конденсируется, синтетические процессы прекращаются, растворяется ядерная оболочка; центриоли расходятся к полюсам; гомологичные хромосомы остаются соединёнными между собой.
К концу профазы I центриоли мигрируют к полюсам клетки, формируются нити веретена деления, разрушаются ядерная мембрана и ядрышки.
· Метафаза I - бивалентные хромосомы выстраиваются вдоль экватора клетки.
· Анафаза I - микротрубочки сокращаются, биваленты делятся, и хромосомы расходятся к полюсам. Важно отметить, что, из-за конъюгации хромосом в зиготене, к полюсам расходятся целые хромосомы, состоящие из двух хроматид каждая, а не отдельные хроматиды, как в митозе.
· Телофаза I - хромосомы деспирализуются и появляется ядерная оболочка.
Второе деление мейоза следует непосредственно за первым, без выраженной интерфазы: S-период отсутствует, поскольку перед вторым делением не происходит репликации ДНК.
· Профаза II - происходит конденсация хромосом, клеточный центр делится и продукты его деления расходятся к полюсам ядра, разрушается ядерная оболочка, образуется веретено деления, перпендикулярное первому веретену.
· Метафаза II - унивалентные хромосомы (состоящие из двух хроматид каждая) располагаются на «экваторе» (на равном расстоянии от «полюсов» ядра) в одной плоскости, образуя так называемую метафазную пластинку.
· Анафаза II - униваленты делятся и хроматиды расходятся к полюсам.
· Телофаза II - хромосомы деспирализуются и появляется ядерная оболочка.
В результате из одной диплоидной клетки образуется четыре гаплоидных клетки. В тех случаях, когда мейоз сопряжён с гаметогенезом (например, у многоклеточных животных), при развитии яйцеклеток первое и второе деления мейоза резко неравномерны. В результате формируется одна гаплоидная яйцеклетка и три так называемых редукционных тельца (абортивные дериваты первого и второго делений).
У некоторых простейших мейоз протекает иначе, чем описанный выше типичный мейоз многоклеточных. Например, может протекать только одно, а не два последовательных, мейотических деления, а кроссинговер - проходить во время другой фазы мейоза. Предполагается, что такой одноступенчатый мейоз примитивен и предшествовал возникновению двухступенчатого мейоза, обеспечивающего эффективную рекомбинацию генома.
Значение мейоза.
· У организмов, размножающихся половым путём, предотвращается удвоение числа хромосом в каждом поколении, так как при образовании половых клеток мейозом происходит редукция числа хромосом.
· Мейоз создает возможность для возникновения новых комбинаций генов (комбинативная изменчивость), так как происходит образование генетически различных гамет.
· Редукция числа хромосом приводит к образованию «чистых гамет», несущих только один аллель соответствующего локуса.
· Расположение бивалентов экваториальной пластинки веретена деления в метафазе 1 и хромосом в метафазе 2 определяется случайным образом. Последующее расхождение хромосом в анафазе приводит к образованию новых комбинаций аллелей в гаметах. Независимое расхождение хромосом лежит в основе третьего закона Менделя.
Дата: 2019-03-05, просмотров: 127.