Метилирование ДНК
| 
| 
|
| Цитозин | 5-метилцитозин | Тимин |
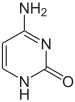
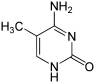
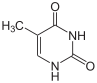
Структура цитозина, 5-метилцитозина и тимина. Тимин может возникать путём деаминирования 5-метилцитозина
Азотистые основания в составе ДНК могут быть ковалентно модифицированы, что используется при регуляции экспрессии генов. Например, в клетках позвоночных метилирование цитозина с образованием 5-метилцитозина используется соматическими клетками для передачи профиля генной экспрессии дочерним клеткам. Метилирование цитозина не влияет на спаривание оснований в двойной спирали ДНК. У позвоночных метилирование ДНК в соматических клетках ограничивается метилированием цитозина в последовательности ЦГ. Средний уровень метилирования отличается у разных организмов от 0 до 1 %. Другие модификации оснований включают метилирование аденина у бактерий и гликозилирование урацила с образованием «J-основания» в кинетопластах.
Метилирование цитозина с образованием 5-метилцитозина в промоторной части гена коррелирует с его неактивным состоянием. Метилирование цитозина важно также для инактивации Х-хромосомы у млекопитающих. Метилирование ДНК используется в геномном импринтинге. Значительные нарушения профиля метилирования ДНК происходят при канцерогенезе.
Несмотря на биологическую роль, 5-метилцитозин может спонтанно утрачивать аминную группу (деаминироваться), превращаясь в тимин, поэтому метилированные цитозины являются источником повышенного числа мутаций.
Повреждения ДНК.
ДНК может повреждаться разнообразными мутагенами, к которым относятся окисляющие и алкилирующие вещества, а также высокоэнергетическая электромагнитная радиация - ультрафиолетовое и рентгеновское излучение. Тип повреждения ДНК зависит от типа мутагена. Например, ультрафиолет повреждает ДНК путём образования в ней димеров тимина, которые возникают при образовании ковалентных связей между соседними основаниями.
Оксиданты, такие как свободные радикалы или пероксид водорода, приводят к нескольким типам повреждения ДНК, включая модификации оснований, в особенности гуанозина, а также двухцепочечные разрывы в ДНК. По некоторым оценкам, в каждой клетке человека окисляющими соединениями ежедневно повреждается порядка 500 оснований. Среди разных типов повреждений наиболее опасные - это двухцепочечные разрывы, потому что они трудно репарируются и могут привести к потерям участков хромосом (делециям) и транслокациям.
Многие молекулы мутагенов вставляются (интеркалируют) между двумя соседними парами оснований. Большинство этих соединений, например, бромистый этидий, даунорубицин, доксорубицин и талидомид, имеет ароматическую структуру. Для того чтобы интеркалирующее соединение могло поместиться между основаниями, они должны разойтись, расплетая и нарушая структуру двойной спирали. Эти изменения в структуре ДНК мешают транскрипции и репликации, вызывая мутации. Поэтому интеркалирующие соединения часто являются канцерогенами, наиболее известные из которых - бензопирен, акридины, афлатоксин и бромистый этидий. Несмотря на эти негативные свойства, в силу их способности подавлять транскрипцию и репликацию ДНК, интеркалирующие соединения используются в химиотерапии для подавления быстро растущих клеток рака.
Некоторые вещества (цисплатин, митомицин C, псорален) образуют поперечные сшивки между нитями ДНК и подавляют синтез ДНК, благодаря чему используются в химиотерапии некоторых видов рака (см. Химиотерапия злокачественных новообразований).
Суперскрученная ДНК.
 Если взяться за концы верёвки и начать скручивать их в разные стороны, она становится короче и на верёвке образуются «супервитки». Так же может быть суперскручена и ДНК. В обычном состоянии цепочка ДНК делает один оборот на каждые 10,459 основания, но в суперскрученном состоянии спираль может быть свёрнута туже или расплетена. Выделяют два типа суперскручивания: положительное - в направлении нормальных витков, при котором основания расположены ближе друг к другу; и отрицательное - в противоположном направлении. В природе молекулы ДНК обычно находятся в отрицательном суперскручивании, которое вносится ферментами - топоизомеразами. Эти ферменты удаляют дополнительное скручивание, возникающее в ДНК в результате транскрипции репликации.
Если взяться за концы верёвки и начать скручивать их в разные стороны, она становится короче и на верёвке образуются «супервитки». Так же может быть суперскручена и ДНК. В обычном состоянии цепочка ДНК делает один оборот на каждые 10,459 основания, но в суперскрученном состоянии спираль может быть свёрнута туже или расплетена. Выделяют два типа суперскручивания: положительное - в направлении нормальных витков, при котором основания расположены ближе друг к другу; и отрицательное - в противоположном направлении. В природе молекулы ДНК обычно находятся в отрицательном суперскручивании, которое вносится ферментами - топоизомеразами. Эти ферменты удаляют дополнительное скручивание, возникающее в ДНК в результате транскрипции репликации.
 Структуры на концах хромосом
Структуры на концах хромосом
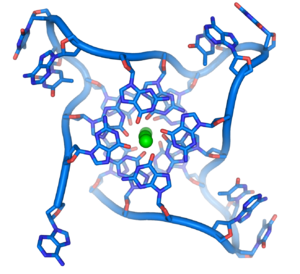
Структура теломер. Зелёным цветом показан ион металла, хелатированный в центре структуры.
На концах линейных хромосом находятся специализированные структуры ДНК, называемые теломерами. Основная функция этих участков - поддержание целостности концов хромосом. Теломеры также защищают концы ДНК от деградации экзонуклеазами и предотвращают активацию системы репарации. Поскольку обычные ДНК-полимеразы не могут реплицировать 3' концы хромосом, это делает специальный фермент - теломераза.
В клетках человека теломеры часто представлены одноцепочечной ДНК и состоят из нескольких тысяч повторяющихся единиц последовательности ТТАГГГ. Эти последовательности с высоким содержанием гуанина стабилизируют концы хромосом, формируя очень необычные структуры, называемые G-квадруплексами и состоящие из четырёх, а не двух взаимодействующих оснований. Четыре гуаниновых основания, все атомы которых находятся в одной плоскости, образуют пластинку, стабилизированную водородными связями между основаниями и хелатированием в центре неё иона металла (чаще всего калия). Эти пластинки располагаются стопкой друг над другом.
На концах хромосом могут образовываться и другие структуры: основания могут быть расположены в одной цепочке или в разных параллельных цепочках. Кроме этих «стопочных» структур теломеры формируют большие петлеобразные структуры, называемые Т-петли или теломерные петли. В них одноцепочечная ДНК располагается в виде широкого кольца, стабилизированного теломерными белками. В конце Т-петли одноцепочечная теломерная ДНК присоединяется к двухцепочечной ДНК, нарушая спаривание цепочек в этой молекуле и образуя связи с одной из цепей. Это трёхцепочечное образование называется Д-петля (от англ. displacement loop).
Биологические функции
ДНК является носителем генетической информации, записанной в виде последовательности нуклеотидов с помощью генетического кода. С молекулами ДНК связаны два основополагающих свойства живых организмов - наследственность и изменчивость. В ходе процесса, называемого репликацией ДНК, образуются две копии исходной цепочки, наследуемые дочерними клетками при делении, отсюда следует, что образовавшиеся клетки оказываются генетически идентичны исходной.
Генетическая информация реализуется при экспрессии генов в процессах транскрипции (синтеза молекул РНК на матрице ДНК) и трансляции (синтеза белков на матрице РНК).
Последовательность нуклеотидов кодирует информацию о различных типах РНК: информационных, или матричных (мРНК), рибосомальных (рРНК) и транспортных (тРНК). Все эти типы РНК синтезируются на основе ДНК в процессе транскрипции. Роль их в биосинтезе белков (процессе трансляции) различна. Информационная РНК содержит информацию о последовательности аминокислот в белке, рибосомальные РНК служат основой для рибосом (сложных нуклеопротеиновых комплексов, основная функция которых - сборка белка из отдельных аминокислот на основе иРНК), транспортные РНК доставляют аминокислоты к месту сборки белков - в активный центр рибосомы, «ползущей» по иРНК.
Структура генома.
Дата: 2019-03-05, просмотров: 232.