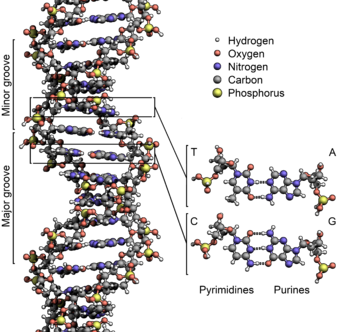
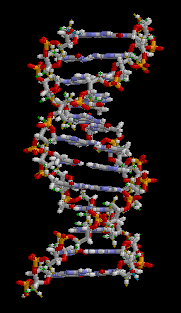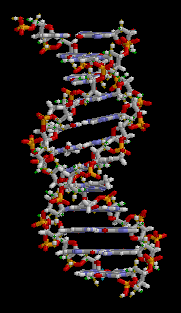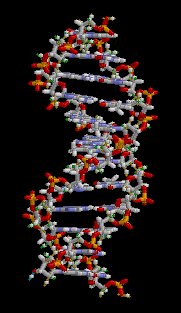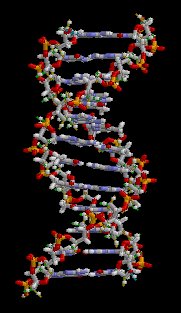
Структура ДНК (двойная спираль). Различные атомы в структуре показаны в разных цветах; детальная структура двух пар оснований показана снизу справа
Дезоксирибонуклеиновая кислота́ (ДНК) – биологический гетерополимер из нуклеотидов. Одна из 3 основных макромолекул (две другие белки и РНК) обеспечивающая хранение, передачу из поколения в поколение и реализацию генетической программы развития и функционирования живых организмов. Молекула ДНК хранит биологическую информацию в виде генетического кода, состоящего из последовательности нуклеотидов. ДНК содержит информацию о структуре различных видов РНК и белков.
В клетках эукариот (животных, растений и грибов) ДНК находится в ядре клетки в составе хромосом, а также в некоторых клеточных органеллах (митохондриях и пластидах). В клетках прокариотических организмов (бактерий и архей) кольцевая или линейная молекула ДНК, так называемый нуклеоид, прикреплена изнутри к клеточной мембране. У них и у низших эукариот (например, дрожжей) встречаются также небольшие автономные, преимущественно кольцевые молекулы ДНК, называемые плазмидами. Кроме того, одно- или двухцепочечные молекулы ДНК могут образовывать геном ДНК-содержащих вирусов.
С химической точки зрения ДНК - это длинная полимерная молекула, состоящая из повторяющихся блоков - нуклеотидов. Каждый нуклеотид состоит из трех компонентов. Азотистого основания, сахара (дезоксирибозы) и фосфатной группы. Связи между нуклеотидами в цепи образуются за счёт дезоксирибозы и фосфатной группы (фосфодиэфирные связи). В подавляющем большинстве случаев (кроме некоторых вирусов, содержащих одноцепочечную ДНК) макромолекула ДНК состоит из двух цепей, ориентированных азотистыми основаниями друг к другу. Эта двухцепочечная молекула закручена по - винтовой линии.

В целом структура молекулы ДНК получила традиционное, но ошибочное название»двойной спирали».
Спираль обычно выглядит так:
 .
.
На самом же деле она является»двойным винтом».
Винтовая линия может быть правой (A- и B-формы ДНК) или левой (Z-форма ДНК).
В ДНК встречается четыре вида азотистых оснований (аденин (A), гуанин (Г), тимин (T) и цитозин (Ц)). В латинской транскрипции (A, G, T, C). Азотистые основания одной из цепей соединены с азотистыми основаниями другой цепи водородными связями согласно принципу комплементарности: аденин (A) соединяется только с тимином (T), гуанин (G) - только с цитозином (C). Последовательность нуклеотидов позволяет»кодировать» информацию о различных типах РНК, наиболее важными из которых являются информационные, или матричные (мРНК), рибосомальные (рРНК) и транспортные (тРНК). Все эти типы РНК синтезируются на матрице ДНК за счёт копирования последовательности ДНК в последовательность РНК, синтезируемой в процессе транскрипции, и принимают участие в биосинтезе белков (процессе трансляции). Помимо кодирующих последовательностей, ДНК клеток содержит последовательности, выполняющие регуляторные и структурные функции. Кроме того, в геноме эукариот часто встречаются участки, принадлежащие «генетическим паразитам», например, транспозонам.
Расшифровка структуры ДНК (1953 год) стала одним из поворотных моментов в истории биологии. За выдающийся вклад в это открытие Фрэнсису Крику, Джеймсу Уотсону и Морису Уилкинсу была присуждена Нобелевская премия по физиологии или медицине 1962 года. Розалинд Франклин, которая получила рентгенограммы, без которых Уотсон и Крик не имели бы возможность сделать выводы о структуре ДНК, умерла в 1958 году от рака (Нобелевскую премию не дают посмертно).
 Френсис Крик,
Френсис Крик,  Джеймс Уотсон,
Джеймс Уотсон,
 Морис Уилкинс,
Морис Уилкинс,  Розалинд Франклин.
Розалинд Франклин.
ДНК как химическое вещество была выделена Иоганном Фридрихом Мишером в 1869 году из остатков клеток, содержащихся в гное. Он выделил вещество, в состав которого входят азот и фосфор. Вначале новое вещество получило название нуклеин, а позже, когда Мишер определил, что это вещество обладает кислотными свойствами, вещество получило название нуклеиновая кислота. Биологическая функция новооткрытого вещества была неясна, и долгое время ДНК считалась запасником фосфора в организме. Более того, даже в начале XX века многие биологи считали, что ДНК не имеет никакого отношения к передаче информации, поскольку строение молекулы, по их мнению, было слишком однообразным и не могло содержать закодированную информацию.
Постепенно было доказано, что именно ДНК, а не белки, как считалось раньше, является носителем генетической информации. Одно из первых решающих доказательств принесли эксперименты Освальда Эвери, Колина Маклауда и Маклина Маккарти (1944 г.) по трансформации бактерий. Им удалось показать, что за так называемую трансформацию (приобретение болезнетворных свойств безвредной культурой в результате добавления в неё мёртвых болезнетворных бактерий) отвечает выделенная из пневмококков ДНК. Эксперимент американских учёных Алфреда Херши и Марты Чейз (эксперимент Херши - Чейз, 1952 г.) с помеченными радиоактивными изотопами белками и ДНК бактериофагов показали, что в заражённую клетку передаётся только нуклеиновая кислота фага, а новое поколение фага содержит такие же белки и нуклеиновую кислоту, как исходный фаг.
Вплоть до 50-х годов XX века точное строение ДНК, как и способ передачи наследственной информации, оставалось неизвестным. Хотя и было доподлинно известно, что ДНК состоит из нескольких цепочек, состоящих из нуклеотидов, никто не знал точно, сколько этих цепочек и как они соединены.
В результате работы группы биохимика Эрвина Чаргаффа в 1949-1951 гг. были сформулированы так называемые правила Чаргаффа. Чаргаффу и сотрудникам удалось разделить нуклеотиды ДНК при помощи бумажной хроматографии и определить точные количественные соотношения нуклеотидов разных типов. Соотношение, выявленное для аденина (А), тимина (Т), гуанина (Г) и цитозина (Ц), оказалось следующим: количество аденина равно количеству тимина, а гуанина - цитозину: А=Т, Г=Ц. Эти правила, наряду с данными рентгеноструктурного анализа, сыграли решающую роль в расшифровке структуры ДНК.
Структура двойной спирали ДНК была предложена Френсисом Криком и Джеймсом Уотсоном в 1953 году на основании рентгеноструктурных данных, полученных Морисом Уилкинсом и Розалинд Франклин, и правил Чаргаффа. Позже предложенная Уотсоном и Криком модель строения ДНК была доказана, а их работа отмечена Нобелевской премией по физиологии или медицине 1962 г. Среди лауреатов не было скончавшейся к тому времени от рака Розалинд Франклин, так как премия не присуждается посмертно. 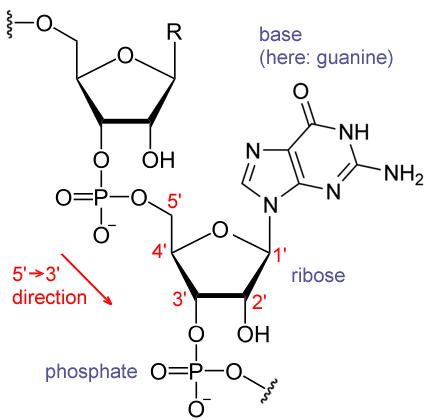 Дезоксирибонуклеиновая кислота (ДНК) представляет собой биополимер мономером которого является нуклеотид. Каждый нуклеотид состоит из остатка фосфорной кислоты, присоединённого по 5'-положению к дезоксирибозе, к которому также через гликозидную связь (C-N) по 1'-положению присоединено одно из четырёх азотистых оснований. Именно наличие характерного моносахарида и составляет одно из главных различий между ДНК и РНК, зафиксированное в названиях этих нуклеиновых кислот (в состав РНК входит моносахарид рибоза).
Дезоксирибонуклеиновая кислота (ДНК) представляет собой биополимер мономером которого является нуклеотид. Каждый нуклеотид состоит из остатка фосфорной кислоты, присоединённого по 5'-положению к дезоксирибозе, к которому также через гликозидную связь (C-N) по 1'-положению присоединено одно из четырёх азотистых оснований. Именно наличие характерного моносахарида и составляет одно из главных различий между ДНК и РНК, зафиксированное в названиях этих нуклеиновых кислот (в состав РНК входит моносахарид рибоза).
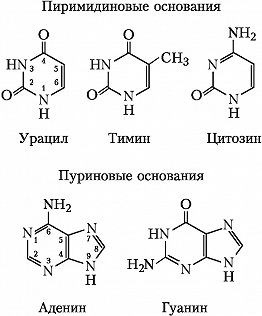 Исходя из структуры молекул, основания, входящие в состав нуклеотидов, разделяют на две группы: пурины (аденин [A] и гуанин [G]) образованы соединёнными пяти- и шестичленным гетероциклами; пиримидины (цитозин [C] и тимин [T]) - шестичленным гетероциклом.
Исходя из структуры молекул, основания, входящие в состав нуклеотидов, разделяют на две группы: пурины (аденин [A] и гуанин [G]) образованы соединёнными пяти- и шестичленным гетероциклами; пиримидины (цитозин [C] и тимин [T]) - шестичленным гетероциклом.
Следует отметить, что тимин (T) и урацил (U) не так строго приурочены к ДНК и РНК соответственно, как это считалось ранее. Так, после синтеза некоторых молекул РНК значительное число урацилов в этих молекулах метилируется с помощью специальных ферментов, превращаясь в тимин. Это происходит в транспортных и рибосомальных РНК.

В зависимости от концентрации ионов и нуклеотидного состава молекулы, двойная спираль ДНК в живых организмах существует в разных формах. На рисунке представлены формы A, B и Z
Модель ДНК Уотсона и Крика описывает структуру одной из нескольких форм двойной спирали. Эта основная форма ДНК, в которой большая часть ее молекул существует в клетке, была названа В-формой.
При понижении влажности В-форма ДНК переходит в А-форму. Изменение конформации углеводного остатка приводит к уменьшению расстояния между нуклеотидными парами вдоль оси спирали (примерно до 25 нм при 11 нуклеотидных остатках на виток спирали). Диаметр спирали увеличивается, изменяется ширина бороздок, пары оснований смещаются к периферии спирали. Вследствие этого внутри спирали возникает полость диаметром около 0,40 нм. А- и В-формы ДНК – это правозакрученные спирали.
При высокой концентрации солей или добавлении спирта спираль В-типа переходит в левозакрученную Z-форму. Левая спираль была обнаружена у полинуклеотида с чередующейся последовательностью нуклеотидов гуанин-цитозин. Повторяющаяся единица в Z-форме состоит из двух пар нуклеотидов (Г-Ц), в результате линия, соединяющая фосфатные группы, через каждые две пары имеет излом и принимает зигзагообразный вид (отсюда название Z-формы).
Полимер ДНК обладает довольно сложной структурой. Нуклеотиды соединены между собой ковалентно в длинные полинуклеотидные цепи. Эти цепи в подавляющем большинстве случаев (кроме некоторых вирусов, обладающих одноцепочечными ДНК-геномами) попарно объединяются при помощи водородных связей во вторичную структуру, получившую название двойной спирали. Остов каждой из цепей состоит из чередующихся фосфатов и сахаров. Внутри одной цепи ДНК соседние нуклеотиды соединены фосфодиэфирными связями, которые формируются в результате взаимодействия между 3'-гидроксильной (3'-ОН) группой молекулы дезоксирибозы одного нуклеотида и 5'-фосфатной группой (5'-РО3) другого. Асимметричные концы цепи ДНК называются 3' (три прайм) и 5' (пять прайм). Полярность цепи играет важную роль при синтезе ДНК (удлинение цепи возможно только путём присоединения новых нуклеотидов к свободному 3'-концу).
У подавляющего большинства живых организмов ДНК состоит не из одной, а из двух полинуклеотидных цепей. Эти две длинные цепи закручены одна вокруг другой в виде двойной спирали, стабилизированной водородными связями, образующимися между обращёнными друг к другу азотистыми основаниями входящих в неё цепей. В природе эта спираль, чаще всего, правозакрученная. Направления от 3'-конца к 5'-концу в двух цепях, из которых состоит молекула ДНК, противоположны (цепи «антипараллельны» друг другу).
Ширина двойной спирали составляет от 22 до 24 Å, или 2,2-2,4 нм, длина каждого нуклеотида 3,3 Å (0,33 нм). Подобно тому, как в винтовой лестнице сбоку можно увидеть ступеньки, на двойной спирали ДНК в промежутках между фосфатным остовом молекулы можно видеть рёбра оснований, кольца которых расположены в плоскости, перпендикулярной по отношению к продольной оси макромолекулы.
В двойной спирали различают малую (12 Å) и большую (22 Å) бороздки. Белки, например, факторы транскрипции, которые присоединяются к определённым последовательностям в двухцепочечной ДНК, обычно взаимодействуют с краями оснований в большой бороздке, где те более доступны.
Образование связей между основаниями.
Каждое основание на одной из цепей связывается с одним определённым основанием на второй цепи. Такое специфическое связывание называется комплементарным. Пурины комплементарны пиримидинам (то есть способны к образованию водородных связей с ними): аденин образует связи только с тимином, а цитозин - с гуанином. В двойной спирали цепочки также связаны с помощью гидрофобных взаимодействий и стэкинга, которые не зависят от последовательности оснований ДНК.
Комплементарность двойной спирали означает, что информация, содержащаяся в одной цепи, содержится и в другой цепи. Обратимость и специфичность взаимодействий между комплементарными парами оснований важна для репликации ДНК и всех остальных функций ДНК в живых организмах.
Так как водородные связи нековалентны, они легко разрываются и восстанавливаются. Цепочки двойной спирали могут расходиться как замок-молния под действием ферментов (хеликазы) или при высокой температуре. Разные пары оснований образуют разное количество водородных связей. АТ связаны двумя, ГЦ - тремя водородными связями, поэтому на разрыв ГЦ требуется больше энергии. Процент ГЦ-пар и длина молекулы ДНК определяют количество энергии, необходимой для диссоциации цепей: длинные молекулы ДНК с большим содержанием ГЦ более тугоплавки.
Части молекул ДНК, которые из-за их функций должны быть легко разделяемы, например, ТАТА последовательность в бактериальных промоторах, обычно содержат большое количество А и Т.
Дата: 2019-03-05, просмотров: 168.