В заключение давайте рассмотрим еще и примеры «эволюции бактерий». Хотя я уже говорил выше - Вы имеете полное право вообще выкинуть такие «примеры» из рассмотрения. По той причине, что нас, прежде всего, интересует, каким образом на Земле появилось потрясающее разнообразие форм организмов. А большинство известных примеров «эволюции бактерий» не имеют к этому вопросу никакого отношения - в этих примерах изменялось что угодно, но только не морфологические признаки бактерий.
Бактерии вообще очень бедны морфологическими признаками. Что там можно особенно анализировать? Шарики, да палочки (кокки и бациллы). Еще цепочки из шариков и изогнутые палочки (стрептококки и вибрионы). И еще гроздья из шариков и палочки, изогнутые спиралью (стафилококки, спириллы и спирохеты). Вот, собственно, и всё «разнообразие» (грубо говоря). Разве сравнится такое морфологическое «разнообразие» всех бактерий с числом морфологических признаков хотя бы только у одного кузнечика (или бабочки), с многочисленными деталями которых можно знакомиться, наверное, неделю, рассматривая их в бинокуляр.
В общем, обсуждать эволюцию форм у бактерий – весьма неблагодарное занятие вследствие крайней бедности этих самых форм. И только актинобактерии с цианобактериями вносят хоть какое-то дополнительное разнообразие в это «царство шариков и палочек».
Но самое главное не это. А то, что бактерии (по крайней мере, некоторые) морфологически никак не изменились за 2-3 млрд. лет их предполагаемой эволюции на планете Земля.
Вот что по этому поводу сказал академик РАН Георгий Александрович Заварзин в своем докладе: [177]
Палеонтологически время появления цианобактерий, морфологически сходных с современными, относят к 2,7 млрд. лет назад. Эти цианобактерии можно определить по современным определителям. Таким образом, цианобактерии представляют персистентную группу и если оценивать эволюцию по численности особей (биомассе) и устойчивости, то они представляют вершину прогрессивной эволюции.
А вот что Г.А. Заварзин написал по поводу целых бактериальных сообществ (Заварзин, 2001):
… Поверхность строматолита (возраст 2.2 млрд. лет) из района Петрозаводска такая же, как у корочки высушенного циано-бактериального мата из лагун Сиваша; другой строматолит (возраст 0.8 млрд. лет) имеет своим аналогом термофильный циано-бактериальный мат с острова Кунашир…
При разнообразии в деталях циано-бактериальные сообщества сохраняют свою архитектуру и состав на протяжении всей геологической летописи. Более того, микропалеонтологи утверждают, что микрофоссилии цианобактерий, возраст которых более 2 млрд. лет, можно классифицировать по современным определителям синезеленых водорослей. Не происходило ни изменений, ни эволюции, выходящей за рамки изменчивости, приуроченной к местообитаниям. Следовательно, изучая современные циано-бактериальные сообщества можно составить представление о микробной биосфере протерозоя…
То есть, мало того, что цианобактерии никак не изменялись на протяжении практически всей истории жизни на Земле. Так они не изменялись еще и целыми бактериальными сообществами.
Помимо поразительных фактов, приведенных академиком Заварзиным, совсем новая научная публикация (Schopf et al., 2015) озвучила полностью аналогичную картину в отношении уже другой группы бактерий. Из этой публикации мы узнаем, что окаменевшие сообщества древнейших серобактерий возрастом 1.8 миллиарда лет, найденные в Западной Австралии, во-первых, оказались идентичны окаменевшему сообществу таких же бактерий возрастом 500 миллионов лет. А во-вторых, эти же (древнейшие) бактериальные сообщества оказались идентичны сообществам современных серобактерий, открытых у побережья Южной Америки в 2007 году (Schopf et al., 2015).[178]
Таким образом, получается пикантная ситуация. Со стороны верующих дарвинистов это весьма зажигательно – доказывать эволюцию живых существ на примере таких биологических созданий, которые вообще не изменялись на протяжении 2 млрд. лет по факту (во всяком случае, каким-нибудь видимым образом).
Кроме этого, имеются и другие серьезные особенности бактерий, которые наводят на размышления - а корректно ли вообще рассматривать примеры с «эволюцией бактерий» в качестве аналогии эволюции любых других живых существ?
Прежде всего, бактерии радикальным образом отличаются от эукариот[179] строением своих клеток. Бактерии устроены гораздо проще и имеют массу отличий от эукариотических клеток, как биохимически, так и морфологически (Рис. 60):
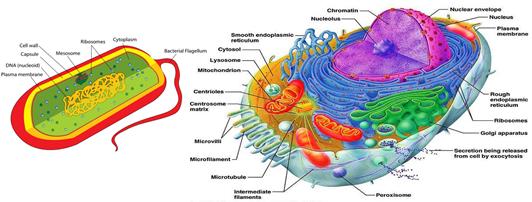
Рисунок 60. Слева схематичное строение бактериальной клетки. Справа схематичное строение эукариотической клетки (пропорции не соблюдены - эукариотические клетки обычно в 10-20 раз больше бактериальных клеток).
Кроме того, бактерии разделяются еще и между собой, и тоже радикальным образом (в биохимическом плане). Настолько, что это потребовало разделения всех бактерий на два отдельных домена – эубактерии и архебактерии, несмотря на их морфологическое сходство друг с другом. Различия между тремя только что озвученными группами живых существ настолько радикальны, что сейчас некоторые авторы предлагают вообще разделять всю существующую на Земле жизнь на три разных формы: эубактерии, архебактерии и эукариоты (Шаталкин, 2004).
Естественно, возникает вопрос, корректно ли переносить те механизмы эволюции, которые мы можем обнаружить у одной формы жизни – на другую форму жизни?
Например, уникальной чертой бактерий является их способность достаточно легко обмениваться между собой разными генами. То есть, осуществлять тот самый горизонтальный перенос генов, который у эукариот напрямую пока еще никто не наблюдал. Зато в мире бактерий горизонтальный перенос является наблюдаемым явлением. Бактерии могут поглощать куски ДНК других бактериальных клеток, например, в ходе процессов коньюгации или трансформации. При этом какие-то отдельные чужие гены вполне могут быть «усвоены» бактерией, поглотившей соответствующую молекулу ДНК, ранее принадлежавшую другой бактерии. Крайне интригующим обстоятельством здесь является то, что поглощенные гены, в принципе, могут быть вообще не от родственной бактерии, а от какой-нибудь удаленной (в таксономическом отношении). Получается, что гены вообще всех видов бактерий, обитающих на каком-нибудь общем участке, в принципе, можно считать единым «генетическим пулом» всех этих бактерий. Особенно те гены, которые находятся в плазмидах, то есть, в тех молекулах ДНК, которыми бактерии обмениваются чаще всего. Стоит ли говорить, что именно в плазмидах, например, нередко сосредоточены гены устойчивости к тем или иным антибиотикам?
Но наверное, самой главной отличительной особенностью бактерий является потрясающая численность их «популяций», которую для подавляющего числа эукариотических организмов даже представить себе невозможно. В одной колонии бактерий может насчитываться миллиарды, десятки или даже сотни миллиардов отдельных особей.
Разве можно сравнивать подобную численность с группами каких-нибудь горных горилл (Gorilla beringei beringei), которые настолько редки, что занесены в Международную Красную книгу? Корректно ли переносить механизмы эволюции, которые теоретически возможны в отношении миллиардных колоний бактерий – на эволюцию горилл?
Но и это еще не всё. Бактерии еще и размножаются очень быстро. В благоприятных условиях у бактерий смена поколений может происходить в течение всего одного часа. Так можно ли сравнивать возможные механизмы эволюции у бактерий, с возможными механизмами эволюции, например, слонов? Если знать, что смена поколений у слонов происходит примерно раз в 17 лет.
Итак, с одной стороны (у бактерий) колоссальная численность особей и фантастическая скорость размножения… а с другой стороны (у эукариот), популяции меньшего размера (сразу на несколько порядков), с гораздо меньшей скоростью смены поколений. Имеем ли мы право ставить знак равенства между этими двумя формами жизни в отношении их возможных механизмов изменений?
Рассмотрим теоретический пример. Допустим, в окружающей среде бактерий возникло такое изменение, которое убьет всех этих бактерий, если у какой-нибудь бактерии срочно не произойдет одной конкретной точечной мутации, которая защищает от этого катастрофического изменения среды. Частота точечных мутаций у бактерий, допустим, одна на миллиард. Тогда весьма возможно, что в многомиллиардной колонии бактерий сразу же и найдется какой-нибудь один (или даже несколько) необходимых мутантов, которые окажутся способными выжить в новых условиях.
А теперь давайте представим себе, что такое же изменение возникло в среде каких-нибудь слонов. Если численность отдельных популяций слонов составляет, допустим, 1000 особей, а всего популяций слонов в этом регионе, допустим, тоже тысяча, тогда общая численность всех слонов, попавших под воздействие новой катастрофической «стихии», составит 1 млн. особей. В этом случае, слонам потребуется примерно 1000 поколений (!), чтобы среди них, наконец, нашлась отдельная особь с нужной мутацией. Поскольку поколения у слонов сменяются раз в 15-20 лет, то получается, что слоны будут находиться под воздействием предположенной нами стихии 10000-20000 лет, пока, наконец, не будет найдена необходимая мутация. Понятно, что за такое время эти слоны просто вымрут, а никуда не эволюционируют. Так можно ли вообще сравнивать механизмы эволюции (адаптации?) бактерий и механизмы эволюции (адаптации?) слонов?
Этот абстрактный пример вполне можно заменить примером, гораздо более близким к реальности. Допустим, появилась какая-то новая линия патогенных вирусов, от которой спасает только та самая, конкретная точечная мутация. Получается, что бактерии вполне могут выжить, понадеявшись на самый тупой вариант адаптации – прямой отбор нескольких особей с нужной мутацией из ста миллиардов других бактерий. А вот слонам такой механизм спасения от новой инфекции уже не подойдет. Если они будут тупо ждать нужную мутацию, они просто вымрут. Поэтому у слонов на этот случай (как и у всех позвоночных животных) имеется специальный адаптационный механизм. При проникновении в организм какой-либо инфекции, в иммунной системе позвоночных начинается специфический ответ. В том числе, происходит гипер-мутагенез (в лимфоцитах), в ходе которого специально подбирается конкретное антитело, наиболее подходящее для уничтожения именно этой инфекции.
Еще один пример. Известно, что среда обитания живых организмов может подвергаться как беспорядочным колебаниям, так и таким изменениям, которые являются более предсказуемыми. Например, одно изменение может предсказуемо следовать за предыдущим. В таком случае живое существо имеет возможность заранее подготовиться к изменению среды, опираясь на уже имеющуюся информацию.
Например, если Вы голодны, а Вам в этот момент протягивают шампур с жареным шашлыком (Вы видите жареное мясо и чувствуете его запах), то в это время было бы полезно, если бы у Вас уже «потекли слюнки». То есть, чтобы Ваш организм немедленно начал готовиться к успешному поглощению пищи. В этом случае Вы работаете как бы на опережение событий – выделяете слюну уже при виде жареного мяса.
Благодаря нашему знаменитому соотечественнику Ивану Петровичу Павлову, все мы уже со школьной скамьи узнаём, благодаря чему наш организм может эффективно предугадывать события. Это происходит благодаря выработке условных рефлексов в нашей нервной системе.
А вот бактерии – не имеют нервной системы. И соответственно, не имеют и условных рефлексов. Однако «работать» на опережение событий они умеют. Без всякой нервной системы. Потому что они имеют громадную численность колоний. Просто благодаря этой численности, бактерии могут находить такие генетические комбинации, которые позволяют им «работать на опережение».
Дадим слово уже многократно помянутому нами Александру Маркову (Марков, 2009б):
…Израильские ученые обнаружили у микроорганизмов (кишечной палочки Escherichia coli и дрожжей Saccharomyces cerevisae) способность к опережающему реагированию, напоминающую классические павловские условные рефлексы. Если в естественной среде обитания микробов один стимул часто предшествует другому, то микробы могут научиться реагировать на первый стимул как на сигнал, предупреждающий о скором появлении второго стимула. В отличие от собак Павлова, микробы приобретают свои «рефлексы» не путем обучения, а за счет мутаций и отбора в длинной череде поколений.
…У организмов, лишенных нервной системы, в том числе у одноклеточных, способности к прижизненному обучению резко ограничены. Но это не значит, что они принципиально не способны к опережающему реагированию. Теоретически, они могут научиться предвосхищать события не хуже павловских собак, но только не за счет прижизненного обучения, а за счет эволюции. Иными словами, вместо «обычной» памяти, которая записывается в структуре межнейронных связей, можно использовать память генетическую, записанную в ДНК. Благодаря гигантской численности популяций микробов, высокой скорости мутирования и очень быстрой смене поколений такое «эволюционное обучение» у микробов теоретически может быть вполне сопоставимо по своей скорости с «обычным» обучением у высших животных…
Как видим, даже самые верующие дарвинисты (в лице Александра Маркова) понимают, что благодаря огромной численности бактериальных колоний, прямой перебор случайных мутаций у бактерий может работать не только в качестве «заменителя» сложной иммунной системы высших животных, но даже в качестве «заменителя» нервной системы, с её механизмами ассоциативного запоминания.[180] И наоборот, высшие организмы, с их крайне малыми численностями популяций (в сравнении с бактериями) не могут позволить себе такую роскошь, как реагирование на быстрые и часто непредсказуемые изменения среды обитания с помощью лишь естественного отбора случайных мутаций. Такое решение было бы гарантированным путем к вымиранию. Поэтому для оперативного реагирования на вызовы среды у высших организмов имеются специальные биологические механизмы.
В связи с этим возникает вопрос. А что мы, собственно, наблюдаем у бактерий, когда они демонстрируют нам очередное приспособление к «сиюминутным» изменениям среды с помощью мелких генетических изменений и естественного отбора? Мы наблюдаем эволюцию этих бактерий? Или же мы наблюдаем просто адаптацию этих бактерий? То есть, может быть, обсуждаемые генетические изменения бактерий и не ведут никуда дальше таких вот мелких приспособлений к локальным особенностям окружающей среды? Возможно, конкретно у бактерий - это просто способ выживания такой – всё время подвергаясь мелким генетическим изменениям… тем не менее, оставаться всё теми же бактериями с незапамятных времен и до сегодняшнего дня. То есть, этакий «бег на месте». Можно ли назвать это эволюцией?
Действительно, получается противоречие – «эволюция» бактерий путем случайных мутаций и естественного отбора идет так быстро, что позволяет бактериальным колониям отвечать на постоянно изменяющиеся вызовы среды чуть ли не в режиме реального времени, почти заменяя в этом отношении формирование рефлексов у высших животных… Но когда мы смотрим на конечные результаты подобной «эволюции», то тут выясняется, что бактерии так и не удосужились измениться хотя бы как-нибудь за 2 миллиарда лет! Этих 2-миллиардолетних бактерий можно определять по современным определителям. Причем некоторые группы бактерий никак не изменялись целыми сообществами (см. выше).
Так может быть, бактерии – это просто такие организмы, которые имеют такую специфическую стратегию реагирования на мелкие изменения окружающей среды - с помощью случайных мутаций и естественного отбора. А вовсе не эволюционируют подобным способом?
Тем более что бактерии отличаются еще одной уникальной особенностью, которая позволяет им, в отличие от высших организмов, свободно использовать в качестве источника полезных инноваций обычный поток случайных мутаций. Дело в том, что бактерии имеют геном очень маленького размера. Поэтому они устойчивы к так называемому «храповику Мёллера». Действительно, проблема «храповика Мёллера» весьма актуальна для организмов с огромным размером генома и слабыми темпами размножения. Представьте себе подобный организм под потоком обычных случайных мутаций, из которых 90% мутаций вредны для выживания, а полезна только одна мутация на миллион. Понятно, что если геном имеет большой размер, а энергия размножения при этом низка, то поток случайных мутаций скорее «разломает» этот геном, чем чего-то там улучшит, даже если естественный отбор будет стараться изо всех сил противодействовать поломке генов. Всё равно поток слабо-вредных мутаций будет постепенно ухудшать качество геномов большого размера, за счет постепенного накопления слабо-вредных мутаций (если темпы размножения данного организма низки).
А вот бактерии, имея крошечный геном и одновременно огромную энергию размножения, вполне могут позволить себе роскошь терять огромное число особей (вплоть до целых линий или даже колоний), оказавшихся чересчур нагруженными вредными мутациями. При этом выигрывать «гонку» (в скорости размножения) будут многочисленные линии, оказавшиеся свободными от вредных мутаций. И уж тем более, линии, которым посчастливилось получить полезную мутацию.
Итак, перефразируя известную поговорку про Юпитера и быка, мы можем с полным основанием сказать:
- Что позволено бактерии, не позволено быку.
То есть, приводить в качестве доказательства возможности эволюции быков по механизму случайные мутации+естественный отбор - примеры мелких генетических изменений у бактерий – не слишком корректно (или даже совсем не корректно).
ТРУДНО БЫТЬ БОГОМ?
А теперь предлагаю немного потренироваться, попробовав себя в роли создателя живых организмов. Представьте, что Вам дали задание – создать животное размером примерно с тигра. Понятно, что популяции таких животных в дикой природе не смогут составлять многие миллиарды особей. Иначе никакой еды на них не напасешься. И так же понятно, что достигать половозрелости всего за несколько часов такие животные тоже не смогут. Животные подобного размера должны будут долго расти, чтобы достигнуть размеров, близких к размеру тигра. То есть, созданное Вами животное подобного размера просто обязано будет иметь низкую численность популяций, и низкую скорость смены поколений.
Но ведь в окружающей среде, в которую Вы должны будете выпустить это животное, наверняка происходят какие-то колебания. И если Вы хотите, чтобы созданное Вами животное, выпущенное в дикую природу, не вымерло сразу же после выпуска, Вы обязаны предусмотреть для этого животного какие-то специальные механизмы, которые позволят ему быстро и эффективно реагировать на неизбежно появляющиеся новые вызовы среды (в том числе, и непредсказуемые). Понятно, что тупой естественный отбор случайных мутаций, с попыткой поиска именно того мутантного «тигра», который сумеет выжить в новой ситуации, здесь не подойдет. Этот механизм в данном случае недопустимо медленный и расточительный.
Здесь можно провести хорошую аналогию с хакерскими атаками. Допустим, имеется задача взломать какую-нибудь систему безопасности, подобрав к ней ключ доступа. Профессиональным хакерам хорошо знаком термин «brute force search», то есть, взлом пароля методом «грубой силы». Это когда необходимый пароль подбирается наиболее тупым способом – путем полного перебора всех возможных вариантов (вплоть до того момента, пока нужная комбинация не совпадёт с одним из вариантов перебора).
Понятно, что если пароль – длинный, то для его взлома методом «грубой силы» требуется большое число попыток. И этих попыток тем больше, чем длиннее пароль. Вплоть до астрономического числа попыток. Чтобы проиллюстрировать, насколько сильно повышается необходимое число попыток при увеличении длины пароля, и сколько времени на этот взлом потребуется компьютеру, приведу соответствующую таблицу: [181]
Таблица 4. Оценочное время полного перебора паролей в зависимости от их длины
| Кол-во знаков | Кол-во вариантов | Стойкость | Время перебора |
| 1 | 36 | 5 бит | менее секунды |
| 2 | 1296 | 10 бит | менее секунды |
| 3 | 46 656 | 15 бит | менее секунды |
| 4 | 1 679 616 | 21 бит | 17 секунд |
| 5 | 60 466 176 | 26 бит | 10 минут |
| 6 | 2 176 782 336 | 31 бит | 6 часов |
| 7 | 78 364 164 096 | 36 бит | 9 дней |
| 8 | 2,821 109 9x1012 | 41 бит | 11 месяцев |
| 9 | 1,015 599 5x1014 | 46 бит | 32 года |
| 10 | 3,656 158 4x1015 | 52 бита | 1 162 года |
| 11 | 1,316 217 0x1017 | 58 бит | 41 823 года |
| 12 | 4,738 381 3x1018 | 62 бита | 1 505 615 лет |
Из таблицы видно - чтобы найти нужный пароль длиной 6 символов, компьютеру Pentium 100 потребуется, в среднем, 6 часов. А вот для того, чтобы найти нужный пароль длиной 12 символов, этому же компьютеру потребуется уже 1.5 млн. лет времени.
Теперь перейдем ближе к нашим «тиграм». Допустим, наши «тигры» в природе столкнулись с новым штаммом вируса, который для них крайне опасен и приводит к 90% гибели в случае заражения. Спасти наших «тигров» может всего одна конкретная точечная мутация (мутация конкретного нуклеотида) в одном из генов, отвечающих за синтез антител к этому вирусу. То есть, вирус немного изменился, став страшно опасным, но спастись от этого вируса вполне можно, если только соответствующее (уже имеющееся) антитело к старому варианту этого вируса тоже немного изменится, подстроившись под новый вариант вируса.
Допустим, частота случайных точечных мутаций у нашего «тигра» составляет примерно 1 к 1 млрд. (на нужный нуклеотид). В этом случае нам потребуется примерно миллиард наших бедных «тигров», чтобы, наконец, «взломать» этот новый вирус с помощью соответствующего (нового) мутантного «тигра». Понятно, что если у нас по всей дальневосточной тайге бегает не более 500 тигров, то тратить миллиард таких тигров для защиты от одной несчастной инфекции – это недопустимая расточительность. То есть, если мы решим, что наши тигры должны адаптироваться к новым инфекциям именно по методике полного перебора, то это гарантированно приведет к вымиранию наших «тигров», над созданием которых мы столь упорно (теоретически) трудились.
Поэтому нам следует позаботиться о том, чтобы снабдить созданных нами «тигров» специальными механизмами, которые будут гораздо более эффективны, чем метод «грубой силы». Хорошим решением здесь будет, например, создание специальной системы гипер-мутирования внутри каждого тигра. То есть, подвергать методу полного перебора следует не самих тигров, а лишь определенные гены в соответствующих клетках, находящихся внутри самих этих тигров. А именно, в таких клетках, которые специально предназначены для борьбы с разнообразными инфекциями (клетки иммунной системы). Таким образом, при появлении новой инфекции, внутри каждого тигра будет происходить быстрый «brute force search». И вполне вероятно, что «тигры» с таким специально встроенным «решателем проблем» успешно преодолеют новую для них напасть. Причем число выживших особей окажется вполне достаточным для будущего устойчивого существования созданного нами вида.
Всё это мы и наблюдаем в реальной природе – сложные и большие организмы с низкими численностями популяций и низкой скоростью смены поколений имеют специальные (встроенные) «решатели проблем» для преодоления огромного числа «сиюминутных» возмущений среды. Начиная от крайне эффективной иммунной системы (в которую как раз встроен тот самый метод «грубой силы») и кончая никак не менее эффективным реагированием на повседневные вызовы среды с помощью механизмов нервной системы у высокоразвитых животных и т.п.
А теперь давайте представим себе, что мы создаем не «тигра», а наоборот – крайне маленькое и очень просто устроенное живое существо. По нашей задумке, таких существ в природе должно быть очень много. И размножаться они должны так быстро, чтобы ими могла питаться вся следующая ступень громадной пищевой пирамиды живых существ, но при этом бы эти мелкие существа не заканчивались.
И вот мы, допустим, создали таких живых существ. И получились эти существа точь в точь похожими на известных нам бактерий.
Понятно, что и перед этими существами тоже будет стоять задача эффективного реагирования на многочисленные мелкие изменения среды (в том числе, и на непредсказуемые). Стоит ли снабжать этих живых существ специальными механизмами решения возникающих проблем? Например, мы можем каким-то образом попытаться «всунуть» в эти существа нервную систему. Но насколько в этом случае замедлится рост и размножение этих микроскопических созданий? Ведь тогда каждой «бактерии» придётся строить и содержать весьма дорогостоящую структуру (нервную систему). А нам была нужна от этих существ максимальная численность и максимальная продуктивность (создание биомассы в единицу времени).
И после небольших раздумий на эту тему мы понимаем, что никаких особо сложных механизмов выживания в такие существа «встраивать» не надо. Вполне достаточно минимума процедур, необходимых для выживания.
Но что же делать со средствами быстрого реагирования на вызовы окружающей среды? Ведь нашим «бактериям» всё-таки нужно как-то бороться с постоянными колебаниями природных условий. И опять после небольших раздумий мы понимаем, что никаких особенных средств в этом случае – тоже не нужно. Поскольку наши микроскопические создания имеют колоссальную численность (в колониях), и не менее колоссальную скорость смены поколений (по сравнению с озвученными выше «тиграми»), то для эффективного реагирования на непредсказуемые изменения среды этим созданиям вполне достаточно того самого метода полного перебора, о котором мы говорили чуть выше. Действительно, если частота точечных мутаций у наших «бактерий» будет такой же, как и у созданных нами «тигров», то есть, 1 к 1 млрд. (на конкретный нуклеотид), тогда поставленная выше задача по «подбору необходимого ключа» к новому смертельно опасному вирусу – может быть успешно решена даже внутри одной единственной колонии бактерий (в первом же поколении). Потому что единственная колония бактерий может насчитывать многие миллиарды особей. Что уж тогда говорить о сотнях и тысячах подобных колоний (которые могут находиться всего на одном метре площади) и о сотнях сменившихся поколений в этих колониях (которые могут смениться в течение одного месяца).
Получается, что механизм случайные мутации+естественный отбор является вполне достаточным для поддержания способности приспосабливаться к мелким изменениям окружающей среды конкретно у бактерий. И способен обеспечить надежное выживание в природе этих микроскопических созданий.
То есть, получается, что Дарвин, когда-то опубликовавший принцип случайные изменения+естественный отбор в качестве механизма глобальной эволюции, на самом деле, может быть, действительно кое-что открыл. Он открыл конкретный адаптивный механизм, с помощью которого бактерии подстраиваются под мелкие изменения окружающей среды. Ну что же, и этот результат – тоже результат.
Итак, давайте подведем предварительные итоги. Ясно, что бактерии и эукариоты – это настолько разные формы жизни, что возникает большой вопрос, насколько вообще корректно сравнивать механизмы эволюции этих двух форм жизни, даже если бы такие механизмы (эволюции) действительно существовали.
Допустим (еще раз) на минуту, что бактерии на самом деле способны эволюционировать, причем именно по механизму случайные мутации+естественный отбор. Ну и какие же результаты этой эволюции? Результаты весьма скромные – отсутствие каких-либо видимых морфологических изменений на протяжении миллиардов лет (см. выше). Этот факт косвенно демонстрирует нам, насколько маломощной эволюционной силой является озвученный механизм эволюции. А вот конкретно слоны (точнее, отряд хоботные), в отличие от бактерий, морфологически изменялись весьма интенсивно (в ходе предполагаемой эволюции). И это несмотря на очень малую численность популяций (в сравнении с бактериями) и несмотря на очень низкую скорость смены поколений (в сравнении с этими же бактериями). Так на каком основании мы должны считать механизмы изменения бактерий и механизмы изменения слонов – одинаковыми?!
Можно, конечно, предположить, что бактерии – это просто такой биологический таксон, для которого именно морфологическая эволюция – крайне затруднена по тем или иным причинам (например, в силу воображаемой исходной «жесткости» организации бактериальных клеток). А вот во всех остальных отношениях бактерии вполне способны эволюционировать. Но это лишь теоретические спекуляции. А по факту у нас получается, что за 3 млрд. лет собственного существования бактерии продемонстрировали очень скромные способности к эволюции форм. И если бы слоны в этом отношении были подобны бактериям, то они бы вообще никуда не сэволюционировали.
Итак, даже если бы сегодняшние бактерии действительно демонстрировали нам совершенно ясную «эволюцию в пробирке», то такие примеры всё равно вряд ли могли бы считаться корректными для объяснения эволюции слонов.
Но мы уже выяснили, что бактерии, скорее всего, демонстрируют нам не «эволюцию в пробирке», а просто «адаптацию в пробирке» (к мелким изменениям окружающей среды), которая конкретно у этих живых существ осуществляется методом «грубой силы» (то есть, собственно, естественным отбором всех возможных случайных мутаций). И никуда дальше этого «эволюция» бактерий не заходит.
Интересно, что для достижения адаптации к локальным изменениям среды, бактерии могут использовать не только метод «грубой силы», но и другие, специальные генетические механизмамы, гораздо более эффективные, чем обычный «полный перебор». Во всяком случае, результаты некоторых исследований в этой области явно свидетельствуют в пользу наличия таких специальных механизмов у бактерий.
Например, весьма впечатляют результаты знаменитых опытов Холла (Hall, 1991). В этих исследованиях Холл получил двойных мутантов, у которых с помощью мутаций было сломано сразу два гена, необходимых для синтеза аминокислоты триптофан. Причем каждая из этих двух мутаций в отдельности - уже лишала бактерию способности синтезировать триптофан. Поэтому мутантные бактерии не могли самостоятельно синтезировать триптофан, а усваивали его только в готовом виде. Далее Холл сажал этих бактерий в среду, в которой отсутствовал готовый триптофан. Триптофан необходим для нормальной жизнедеятельности бактерий, но получить сразу нужного двойного мутанта, то есть, такого мутанта, у которого нужные нуклеотиды восстановлены обратно (необходимой двойной мутацией), вряд ли стоило ожидать, поскольку вероятность такой (двойной) мутации слишком низка. Поэтому будущее этих несчастных бактерий в среде без готового триптофана представлялось совершенно печальным. Однако к удивлению Холла (и всей остальной научной общественности), эти бактерии по-быстрому умудрились отремонтировать себе нужные гены обратно, и снова стали синтезировать триптофан, как ни в чем не бывало. То есть, бактерии продемонстрировали (по факту), что они способны получать жизненно необходимые им двойные мутации примерно в 100.000.000 раз чаще (!), чем это ожидалось при чисто случайном распределении мутаций. Такой результат опытов Холла выглядел чуть ли не мистически. Создавалось ощущение, что бактерии просто знают, что и как им надо отремонтировать у себя в ДНК, чтобы выжить.
Позже были обнаружены такие факты, в свете которых мистичность результатов, полученных Холлом, немного поуменьшилась. Оказалось, что бактерии, помимо обычного метода «полного перебора», всё-таки имеют еще один, специально встроенный «решатель проблем». Видимо, для таких ситуаций, которые совсем уже «из ряда вон».
Бактерии (и не только бактерии) имеют особый генетический комплекс - SOS-систему, которая в нормальных условиях почти бездействует. SOS-система начинает активно работать в случае возникновения множественных повреждений молекул ДНК. Эти повреждения могут быть вызваны самыми разными причинами – начиная от ультрафиолетового излучения и кончая воздействием ряда антибиотиков. В таких случаях SOS-система производит экстренный ремонт молекул ДНК.
И вот (как выяснилось) SOS-система бактерий устроена таким образом, что в совсем уже экстремальных случаях (когда повреждения ДНК становятся особенно массированными), SOS-система начинает ремонтировать ДНК специфическим образом – она начинает специально допускать гораздо больше генетических ошибок, чем в своем обычном режиме работы. По сути, SOS-система бактерий переключается с режима ремонта ДНК в режим гипер-мутагенеза этой ДНК. Переход SOS-системы из режима репарации в режим гипер-мутагенеза обеспечивается включением целого ряда специальных генов, которые в свою очередь, достаточно «умны», чтобы включаться только в самых экстренных случаях (так устроена эта генетическая система).
В результате включения режима гипер-мутагенеза, частота возникновения мутаций в ДНК может повыситься сразу на несколько порядков. Понятно, что в таких условиях быстро находить необходимые варианты одинарных или даже двойных мутаций методом «полного перебора» становится намного легче (учитывая огромную численность бактериальных колоний). Таким форсированным способом можно быстро получить, например, устойчивость к тому или иному антибиотику (или к другому разрушительному фактору среды).
Однако решать проблемы подобным способом – это уже так называемый «вариант атас». Потому что значительная часть бактерий станет получать мутации, либо вредные, либо вообще несовместимые с жизнью. В результате, бактериальная колония в целом станет либо хуже расти, либо вообще сокращаться. Но видимо, иногда проблемы бывают настолько серьезными, что всё-таки требуют включения механизма гипер-мутирования. Пусть большая часть бактерий при этом погибнет от вредных мутаций, но зато какие-то отдельные счастливчики могут стать обладателями именно такого генетического изменения, которое успешно решит возникшую проблему выживания. И в результате, данная линия бактерий сможет продолжить своё существование.
Таким образом, получается, что бактерии для приспособления к среде могут использовать не только обычный вариант «полного перебора», но и его форсированный вариант (во всяком случае, в некоторых ситуациях). Такую способность можно рассматривать, как еще один (дополнительный) механизм, которым кто-то заботливо снабдил (?) бактерий, для того чтобы повысить надежность их выживания при самых экстремальных возмущениях среды. Действительно, трудно представить, каким образом подобный генетический механизм мог бы появиться у бактерий естественным образом. Ведь переход SOS-системы в режим гипер-мутагенеза полезен только иногда, в самых экстремальных условиях (для получения необходимой редкой мутации). В обычных же условиях, такой режим работы будет приводить к снижению конкурентоспособности тех бактериальных линий, в которых SOS-система вздумает начать работать именно в таком режиме. Потому что в этом случае образуется слишком много бактериальных клеток с вредными мутациями. Следовательно, для того чтобы приносить пользу (а не вред!), такой режим работы SOS-системы должен был появиться сразу в готовом виде, то есть, уже уметь надежно «засыпать» в нормальных условиях и просыпаться только в самых экстремальных. Но такая особенность работы SOS-системы обеспечивается несколькими специальными генами. Поэтому представить, как это могло образоваться методом постепенных «случайных тыков» – не очень получается.
Впрочем, проблема естественного возникновения подобного генетического комплекса ничуть не тяжелее проблемы естественного возникновения какой-нибудь системы свертываемости крови, или механизма мейоза, или биохимического пути гликолиза.
А теперь давайте вернемся обратно к результатам опытов Холла (Hall, 1991). Можно предположить, что бактерии, которые в экспериментах Холла поразительно быстро восстанавливали способность синтезировать триптофан - делали это не в силу обладания мистическими способностями, а просто в результате резкого повышения общего мутационного фона (вызванного, допустим, работой озвученной SOS-системы в режиме мутагенеза). Вследствие чего необходимая двойная мутация находилась гораздо быстрее (в 100 млн. раз быстрее), чем это ожидалось в обычном «режиме» жизнедеятельности бактерий.
Однако при ближайшем рассмотрении такое предположение перестаёт выглядеть привлекательным.
Во-первых, потому что той частоты мутагенеза, которую могут обеспечить гены-мутаторы SOS-системы, всё равно не хватает для объяснения результатов опытов Холла. Например, в работе (Maisnier-Patin et al., 2005) исследователи использовали один из генов-мутаторов бактериальной SOS-системы, но даже самая активная работа этого гена обеспечила повышение частоты разных[182] мутаций не более чем на 2 порядка (то есть, всего в 100 раз или немного больше). Однако Холл оценил скорость восстановления «триптофановых способностей» у своих бактерий - в 100 миллионов раз выше, чем это ожидалось бы при случайном распределении мутаций (происходящих в обычном темпе мутагенеза). Получается, что наблюдаемая скорость восстановления бактерий в эксперименте Холла всё равно была в миллион раз выше, чем это ожидалось бы методом «полного перебора», даже после его форсирования в 100 раз.
Во-вторых, в работе (Maisnier-Patin et al., 2005) было показано, что работа гена-мутатора приводит к более-менее равномерному распределению мутаций по всему геному бактерии, при общем повышении частоты разных мутаций в 100 раз. В связи с этим возникает вопрос – а способны ли вообще бактериальные колонии выдержать повышение мощности мутагенеза не в 100 раз, а в 100.000.000? Не получится ли при такой частоте мутаций полного вымирания бактериальных колоний, вследствие мгновенного накопления вредных мутаций практически в каждой бактериальной клетке?
В общем, маловероятно, что быстрое восстановление бактерий (из состояния двойного мутанта) в экспериментах Холла происходило за счет общего повышения уровня мутагенеза. Поэтому не исключено, что «недра» бактериальной клетки скрывают внутри себя еще какой-то (пока не очень понятный) специальный механизм, который в ответ на вызовы среды способен подбирать необходимые генетические изменения гораздо эффективнее, чем обычный мутагенез, и даже эффективней, чем гипер-мутагенез бактериальной SOS-системы.
То есть, бактерии, оказывается, не так просты, чтобы адаптироваться к изменениям окружающей среды только с помощью естественного отбора случайных мутаций в обычном режиме мутирования. Помимо обычных случайных мутаций они могут: 1) обмениваться между собой нужными генами в ходе горизонтального переноса, 2) использовать форсированный вариант «полного перебора» (гипер-мутагенез), 3) возможно, обладают еще какими-то (более «умными») механизмами адаптаций.
Ну а каковы же возможности самого примитивного генетического механизма адаптации бактерий - метода «полного перебора» в обычном режиме? По этому поводу имеется некоторый разнобой фактов.
С одной стороны, имеются данные, что такой способ весьма неплох. Например, в работе (Weinreich et al., 2006) исследователи на практике показали, что «brute force search» в исполнении бактерий, в принципе, способен на многое. В этой работе исследователи продемонстрировали, что у бактерий, с которыми они работали, может выработаться устойчивость к новому антибиотику цефотаксиму. Для этого в ДНК исходных бактерий должны произойти 5 конкретных точечных изменений (мутаций). В этом случае, устойчивость бактерий к новому антибиотику возрастает в 100000 раз. Понятно, что сразу все 5 нужных изменений появиться у бактерий не могут. Для подобной мутации не хватит вообще всех существующих бактерий на планете Земля. Действительно, если частота точечной мутации (с заменой нужного нуклеотида) будет одна к миллиарду, то вероятность того, что сразу 5 нужных нуклеотидов мутируют именно нужным образом внутри какой-нибудь одной бактерии, составит 1 к 1045. Понятно, что такое событие не случится вообще никогда.[183]
Более того, авторы исследования благоразумно рассудили, что даже нужную двойную мутацию (где сразу два конкретных нуклеотида мутируют внутри одной бактерии) тоже вряд ли можно сразу получить методом полного перебора. Потому что такая двойная мутация (вероятность 10-18) потребует количества бактерий (1018 особей), которое вряд ли можно насобирать в пределах воздействия нового антибиотика.
Поэтому авторы работы (Weinreich et al., 2006) решили посмотреть, можно ли получить из исходного немутантного гена, мутантный ген, несущий все 5 нужных мутаций - с помощью цепочки из пяти последовательных одинарных (нужных) мутаций.
То есть, они рассматривали вариант, когда одновременно происходит только одна единственная мутация (из пяти необходимых). И далее исследовали, смогли бы бактерии (в принципе) приобрести конечную устойчивость к новому антибиотику путем «пошагового» (постепенного) приобретения всех пяти мутаций, с величиной шага в одну единственную мутацию. Или не смогли бы. Действительно, это весьма интригующий вопрос. Ведь для того, чтобы естественный отбор последовательно поддержал все пять мутаций, каждая новая мутация, взаимодействуя с предыдущими мутациями, должна повышать, а не понижать устойчивость к новому антибиотику.
Например, одна из пяти мутаций, исследовавшихся в этой работе, не увеличивает, а наоборот, снижает устойчивость к антибиотику, поскольку соответствующий фермент, получив эту мутацию, начинает разрушать данный антибиотик медленнее, чем без этой мутации. Зато эта мутация заметно повышает стабильность («время жизни») этого фермента в цитоплазме самой бактерии. Другая мутация приводит к тому, что соответствующий фермент начинает разрушать антибиотик быстрее. Но зато из-за этой мутации фермент становится менее стабильным (сам быстрее разрушается). А вот двойной мутант по только что озвученным мутациям – уже имеет резко повышенную устойчивость к антибиотику. Потому что соответствующий фермент (с этими двумя мутациями), с одной стороны, быстрее разрушает антибиотик, а с другой стороны, сам «живет» внутри бактериальной клетки дольше. Получается, что например, первая мутация вообще не сможет закрепиться в колонии бактерий до тех пор, пока бактерия не получит вторую мутацию.
То есть, каждая из пяти мутаций определенным образом изменяет конформацию соответствующего белка (фермента, расщепляющего антибиотик). И теоретически, каждое такое изменение может либо «разрешить», либо, наоборот, «запретить» закрепление следующей мутации (вследствие отрицательного отбора).
Поэтому авторы исследования экспериментально проверили все возможные варианты, в каком порядке могли бы появляться 5 нужных одиночных мутаций. Число этих вариантов 120 (факториал из 5). И действительно, обнаружили несколько путей (из возможных 120), с помощью которых каждая из 5 конкретных мутаций, возникая, повышала в той или иной степени устойчивость к антибиотику, пока, наконец, все 5 нужных мутаций не давали бактерии практически полную защиту от антибиотика (повышали устойчивость бактерии к цефотаксиму в 100 тысяч раз).
Таким образом, авторы работы (Weinreich et al., 2006) показали на практике, что метод «полного перебора» даже при величине «шага» всего в одну мутацию - может приводить к весьма впечатляющим результатам, надежно защищая бактерий от непредсказуемых условий среды.
Правда, остался невыясненным вопрос, а что будет, если для достижения результата (возникновения полезного признака или свойства) нужно не 5 одновременных мутаций, а например, 20? Найдется ли в этом случае непрерывная «дорожка» из отдельных «шажков», каждый из которых будет повышать приспособленность? Или уже не найдется?
Впрочем, этот вопрос уже из области знаменитой «проблемы неуменьшаемой сложности» (Behe, 1996). То есть, из серии – а можно ли собрать паровоз путем постепенного присоединения к нему отдельных деталей таким образом, чтобы каждая деталь хотя бы немного повышала функциональность этого паровоза? Задаваться такими теоретическими вопросами мы здесь не будем. А лучше посмотрим, какие результаты и возможности демонстрируют нам сами бактерии, вооруженные методом «прямого перебора».
К сожалению, в целом эти результаты всё-таки не впечатляют. «Дорожка» из 5 мутаций, повышающих в итоге приспособленность в 100000 раз – это конечно круто. Но в большинстве реальных случаев, когда бактериям срочно необходимо к чему-нибудь приспособиться, дело обычно оканчивается всего одной (максимум, двумя) мутациями. И на этом этапе всё и останавливается. Причем в итоге получаются отнюдь не идеальные варианты, а как правило, уже известные нам «полезные поломки». То есть, когда бактериям срочно нужно к чему-нибудь приспособиться, то подхватывается любая мутация, которая может помочь в этой ситуации. Но эта же мутация одновременно серьезно ухудшает что-нибудь другое. И в результате, такие бактерии, содержащие «полезную поломку», проигрывают в борьбе за выживание обычным («не поломанным») бактериям, как только условия среды возвращаются в нормальное русло. То есть, подобные «полезные поломки», полученные методом «грубой силы», можно рассматривать лишь как временные реакции бактерий на какие-то экстремальные возмущения среды. А потом «всё возвращается на круги своя» (и окружающая среда, и живущие в ней бактерии).
Например (допустим) какая-то бактерия использует специальный белок для активного транспорта внутрь собственной клетки какого-нибудь полезного для неё вещества. И тут Вы создаёте антибиотик, молекула которого похожа на это полезное вещество. В результате, бактерии начинают закачивать внутрь себя этот новый яд, и погибают. Однако у какой-то бактерии случилась «полезная поломка» - тот белок, который транспортировал полезное вещество через мембрану – мутировал. И в результате стал плохо выполнять свои функции. В обычных условиях такая бактерия – является калекой. Она будет расти и размножаться хуже своих соседок. Однако в условиях отравления антибиотиком, эта бактерия получает преимущество. Потому что все нормальные бактерии в этих условиях исправно накачивают себя ядом. А больная бактерия этого делать не может.
Вот такие «калеки» обычно и получаются в результате знаменитого «приспособления бактерий к антибиотикам», о котором прожужжали все уши верующие дарвинисты, выдавая такие приспособления за «доказательства эволюции». То есть, те самые «супер-бактерии» из наших больниц, которыми нас так пугают свидетели Дарвина (будто бы эти бактерии настолько «эволюционировали», что стали устойчивы чуть ли не ко всем антибиотикам сразу)… на самом деле, в большинстве своем являются весьма серьезными калеками и инвалидами. И по этой причине в естественной среде не могут быть конкурентами обычным здоровым колониям исходных бактерий.
Наверное, так бывает не всегда. Ведь теоретически, методом «грубой силы» можно получить не только «полезные поломки», но и какие-нибудь более полезные вещи. Но по факту, о появлении у бактерий чего-нибудь полезного и при этом невредного - слышно очень редко. А уж о возникновении методом «грубой силы» каких-нибудь принципиально новых (полезных) генов, и уж тем более, новых биохимических путей - вообще не слышно.
В тех же случаях, когда для борьбы с теми или иными антибиотиками у бактерий обнаруживаются какие-то особые гены (т.е. гены, специально предназначенные для борьбы с тем или иным антибиотиком)… неожиданно оказывается, что ископаемые бактерии возрастом в десятки тысяч или даже в миллионы лет - уже обладали этими генами. Но об этом чуть ниже.
А сейчас давайте разберем один конкретный пример такой «полезной поломки» у бактерий, который, благодаря совместным усилиям всех верующих дарвинистов мира, стал, наверное, самым известным примером «доказательства эволюции» у бактерий. Чуть ли не «притчей во языцех».
Речь идет о «бактериях, научившихся есть цитрат», в экспериментальных исследованиях коллектива Ленски (Blount et al., 2008). Это наиболее модный пример, известный сегодня в области «наблюдаемой эволюции бактерий». Действительно, если рассматривать этот пример «с высоты птичьего полёта», то результат выглядит впечатляюще – бактерии сначала не умели есть цитрат, а потом (в результате случайной мутации) научились его «кушать». То есть, освоили новый вид «корма» в ходе естественной эволюции.
Но дело в том, что если не «пролетать над гнездом кукушки» (С), а рассмотреть этот пример внимательно, то первое, что мы выясним - на самом деле, бактерии кишечной палочки всегда умели кушать цитрат. Просто в природе они делают это только в выгодных для себя (особых) условиях.
В эксперименте, поставленном коллективом Ленски, бактерии искусственно пересаживались в колбы с питательной средой, где они питались глюкозой. Кроме того, в питательной среде содержался еще и цитрат (лимонная кислота).
Цитрат не усваивается бактериями кишечной палочки в присутствии кислорода. В кислородных условиях кишечная палочка использует гораздо более эффективные пути добычи энергии (более эффективные в десятки раз), чем те, которые даёт потребление конкретно цитрата. Поэтому кишечная палочка начинает использовать цитрат только в условиях отсутствия кислорода. Такая способность реализуется за счет того, что ген, который кодирует белок-переносчик цитрата (необходимый для того, чтобы бактерия стала потреблять цитрат), связан с управляющим участком ДНК (промотором), который подавляет работу гена, синтезирующего белок-переносчик цитрата, если в среде присутствует кислород.
И вот, в результате, бедные колонии бактерий на протяжении 30.000 поколений в этом эксперименте практически «сидели» на цитрате, но не могли его использовать. И только примерно на 31.000 поколении произошла соответствующая мутация – описанный выше генный комплекс (оперон), наконец, сломался. А именно, ген, кодирующий только синтез белка-переносчика цитрата, дополнительно скопировался еще и в другую область ДНК, тем самым вырвавшись из того оперона, который блокировал его работу в присутствии кислорода, и попал в оперон, не чувствительный к фактору присутствия кислорода. Поэтому этот ген стал работать всё время, то есть, соответствующий белок-переносчик цитрата начал синтезироваться бактериальной клеткой всё время. В результате, мутантные кишечные палочки начали усваивать цитрат тоже всё время – как в условиях отсутствия кислорода, так и в его присутствии.
Говоря простыми словами, произошло примерно следующее. Раньше у Вас на кухне был работающий кран. Поэтому вода из него текла только тогда, когда Вам это было нужно (когда Вы открывали этот кран). Но в какой-то момент этот кран – сломался. И теперь вода из него течет всё время. Однако в каких-то особых условиях это «новое свойство» Вашего крана может оказаться полезным. Например, теперь Вы точно не умрете от жажды, даже если у Вас связаны руки.
Вот так же и поломка генного комплекса, регулирующего потребление цитрата, оказалась полезной в специфических условиях, заданных экспериментаторами.
Таким образом, в этом эксперименте:
1). Бактерии «научились» есть цитрат не за счет возникновения каких-то новых генов, а за счет поломки уже имевшегося генного комплекса.
2). На то, чтобы сломать соответствующий генный комплекс, у бактерий ушло более 31.000 поколений! Здесь надо вспомнить, что колонии бактерий (в одной пробирке) насчитывают миллиарды особей. Тем не менее, потребовалось ждать 31.000 поколений до нужной поломки.
Что уж тогда говорить об эволюции каких-нибудь тигров или горилл, популяции которых насчитывают всего сотни особей? При этом (еще раз) речь здесь идет не о возникновении какого-нибудь нового гена, а лишь о поломке уже имеющихся генов, конкретно, о разрыве связи между геном, синтезирующим нужный белок, с его исходным опероном. В свете таких интересных подробностей, возможные перспективы воображаемой эволюции бактерии в человека по такому же (!) механизму – начинают выглядеть весьма удручающими. Наверное, можно даже сказать, что эксперимент Ленски наглядно показал невозможность такой эволюции (по механизму случайные мутации + естественный отбор). Ведь для эволюции бактерии в человека, во-первых, требуется возникновение многих тысяч принципиально новых генов. А не поломка уже имеющихся генных комплексов. А во-вторых, воображаемые предки людей были высшими животными, поэтому не могли иметь популяции размером многие миллиарды особей. Что обесценивает возможности механизма «полного перебора» просто критически.
Тем не менее, эксперимент коллектива Ленски на этом не был остановлен. Биологи продолжали наблюдать за теми бактериями, которые в результате озвученной «полезной поломки» приобрели способность потреблять цитрат. Пока они делали это еще слабо, поэтому скорость их размножения еще очень мало отличалась от бактерий, не имевших подобной мутации (Blount et al., 2008; Blount et al., 2012).
Но в следующих поколениях бактерий, мутантный ген (оторвавшийся от своего кислород-регулирующего промотора и «примкнувший» к промотору, безразличному к наличию кислорода), скопировался в ДНК бактерий, возможно, еще несколько раз подряд. В результате, синтез белка-переносчика цитрата тоже резко усилился. И как следствие, серьезно усилилась транспортировка цитрата через бактериальную мембрану и его усвоение кишечной палочкой. Таким образом, в течение еще нескольких тысяч поколений исходная кишечная палочка научилась кушать цитрат, как говорится, «на полную катушку». То есть, здесь имела место не просто (одна) полезная поломка, а этот результат был еще усилен дополнительными мутациями – созданием дополнительных копий нужного гена. Явление, тоже не редкое у бактерий – когда они наращивают в ДНК копии тех генов, которые дают им сиюминутное преимущество в какой-то конкретной среде (например, дополнительные копии тех генов, которые ответственны за синтез белков, разрушающих тот или иной антибиотик).
Интересно, что в эксперименте Ленски мутантные бактерии, научившиеся эффективно усваивать цитрат в любых условиях, так и не смогли до конца вытеснить исходные бактерии (усваивающие цитрат только в отсутствии кислорода). Оно и понятно. Если вокруг есть глюкоза, то ставить на постоянный «поток» синтез белка-переносчика цитрата, является не экономичным занятием. В это время лучше сосредоточиться на синтезе белков, расщепляющих глюкозу. То есть, мы опять наблюдаем, что «бактерию, научившуюся есть цитрат», в принципе, можно считать своеобразным «инвалидом», который в естественных условиях неизбежно проиграет исходным бактериям. Видимо поэтому кишечные палочки и оставили себе «лазейку» (для возвращения к своему нормальному состоянию) в виде некоторого числа исходных (не мутантных) особей?
В этот момент верующий дарвинист может, конечно, начать иронизировать, говоря о том, что таким же образом в качестве «инвалида» можно представить вообще любое живое существо, приспособленное к конкретной среде обитания. Например, гималайского медведя можно представить в качестве «инвалида», проигрывающего белому медведю в условиях Арктики. Однако факт остаётся фактом – ни окончательного разделения ниш, ни окончательного вытеснения в эксперименте Ленски пока не получилось. Как не получается аналогичных разделений и вытеснений и в естественных, «диких» колониях кишечной палочки. Никто пока, вроде бы, не зафиксировал в реальной природе возникновения новых видов кишечной палочки. Да что там кишечные палочки. Озвученные выше цианобактерии не потрудились измениться и за 2 млрд. лет собственной «эволюции». Хотя цианобактерии, наверное, способны к генетическим перестройкам собственной ДНК никак не меньше кишечных палочек. Тем не менее, никакой (видимой) эволюции за 2 млрд. лет.
Более того, имеются и такие факты, которые превращают ситуацию с «эволюцией бактерий» уже в полный анекдот. Дело в том, что периодически ученые находят и «размораживают» бактерий очень древнего возраста. Поскольку «эволюция бактерий» часто доказывается именно указанием на их способность приспосабливаться к воздействию антибиотиков, то в ряде исследований биологи проверили древних бактерий именно на устойчивость к этим самым антибиотикам. И оказалось, что древние бактерии… столь же устойчивы к современным антибиотикам, как и сегодняшние бактерии. Причем устойчивость к антибиотикам древним бактериям обеспечивают… те же самые гены, которые сегодня ответственны за то же самое в ДНК современных бактерий.
Конкретно, устойчивость к современным антибиотикам продемонстрировали «размороженные бактерии» возрастом 30000 лет (D'Costa et al., 2011).
Такую же устойчивость к современным антибиотикам показали бактерии возрастом 4000000 лет (Bhullar et al., 2012).[184] Некоторые штаммы этих бактерий оказались устойчивыми сразу к 14 различным коммерчески доступным антибиотикам. В том числе, к сравнительно новому даптомицину, полусинтетическим макролидным антибиотикам и т.п.
Наконец, еще одно исследование, проведенное совсем недавно, установило, что у индейцев изолированного племени яномами (Южная Америка), которые никогда не сталкивались с антибиотиками - бактерии их кишечной флоры, тем не менее, обладают целым набором специальных генов устойчивости к разным современным антибиотикам. В том числе, и к современным синтетическим антибиотикам (Clemente et al., 2015).
Спрашивается, ну и какие же изменения в «жизни бактерий» мы наблюдаем? Бактерии, оказывается, испокон веков обладают способностью сопротивляться воздействию антибиотиков. Вот тебе и «эволюция бактерий».
Причем благодаря широко распространенному у бактерий горизонтальному переносу генов (за счет захвата ДНК других бактерий), эти маленькие создания, в принципе, способны к распространению уже имеющихся (испокон веков?) генов устойчивости к разным антибиотикам чуть ли не по всему бактериальному царству. В этом отношении, бактерий, наверное, можно рассматривать, как некий сверх-организм, эдакий «океан-Солярис», с громадным совокупным генофондом, включающим в себя гены «на все случаи жизни». Может быть, бактерии так и выживают, испокон веков отбивая атаки антибиотиков с помощью существующей (тоже испокон веков) «общей базы данных» генов устойчивости к этим антибиотикам?
Но тогда при чем здесь вообще «эволюция»?
В общем, в деле «эволюции бактерий» всё обстоит примерно так же, как и в деле «эволюции» всех остальных живых организмов (эукариот). За «доказательства эволюции» эукариот верующие дарвинисты пытаются выдать, как правило, наблюдаемые факты внутривидовой изменчивости (за исключением нескольких интересных примеров изменений, произошедших явно не по дарвиновским механизмам). А за «эволюцию бактерий» они выдают наблюдаемые факты адаптации этих бактерий к локальным особенностям и мелким колебаниям условий среды обитания.
И потом предлагают Вам поверить, что вот точно по такому же механизму постепенно появился и человек из бактерии. Несмотря на то, что никаких фактических оснований для столь смелых обобщений – даже близко не существует.
Действительно, давайте решим простую задачку:
Исходные условия:
Известно, что за 30.000 поколений бактерии могут сломать уже имеющийся генный комплекс (см. выше).
Вопрос:
За какое число поколений может возникнуть биохимический цикл гликолиза, который включает в себя десять разных белков-ферментов (соответственно, 10 разных генов, кодирующих эти белки), где каждый из ферментов работает только на одной стадии, передавая «эстафету» на следующую стадию следующему ферменту (то есть, эти ферменты бессмысленны друг без друга). Причем первые этапы гликолиза происходят с затратами энергии. Поэтому первые стадии были бы не только бесполезны, но даже вредны, если бы не было заключительных стадий гликолиза, где происходит уже накопление энергии, с итоговым положительным балансом.
Итак, за какое время возникнет подобный комплекс, состоящий из тысяч необходимых нуклеотидов, если бактерии могут сломать один (уже имеющийся) ген за 30.000 поколений?
Правильный ответ:
Результаты экспериментов Ленски вообще не позволяют судить о возможности (или невозможности) возникновения гликолиза методом «полного перебора». Поскольку, как говорится, «ломать не строить». Но и так понятно, что методом «полного перебора» мы будем получать работающий гликолиз дольше, чем всё время существования Нашей Вселенной.
Вот на этой «оптимистической» ноте я и предлагаю закончить эту книгу.
Здесь я попробовал критически рассмотреть (и в ряде случаев переосмыслить) те фактические свидетельства, которые насобирал в свою пользу дарвинизм со времени первого опубликования этой теории Дарвиным в 1859 году. И как оказалось, этих (фактических) свидетельств, во-первых, крайний дефицит, а во-вторых, практически все они либо недостаточно строги (надежны), либо недостаточно впечатляющи (можно объяснить другими способами), либо вообще не имеют никакого отношения к дарвиновским механизмам. Мне кажется, это печальное обстоятельство верующим дарвинистам просто необходимо учитывать в проповедях своего любимого и «единственно верного» учения.
Ну а следующую книгу я посвящу, во-первых, фактам, которые опровергают дарвинизм. Некоторые из таких фактов я уже озвучил в этой книге. Но мне кажется, необходим более подробный перечень таких фактов. А во-вторых, я посвящу следующую книгу фактам, которые, на мой взгляд, очень серьезно свидетельствуют в пользу разумного дизайна живых организмов, как основной причины наблюдаемого нами разнообразия живых существ.
ЛИТЕРАТУРА
Alford R.A., Brown G.P., Schwarzkopf L., Phillips B., Shine R. Comparisons through time and space suggest rapid evolution of dispersal behaviour in an invasive species // Wildlife Research. 2009. V. 36. P. 23-28.
Allentoft M.E., Collins M., Harker D., Haile J., Oskam C.L., Hale M.L., Campos P.F., Samaniego J.A., Gilbert M.T.P., Willerslev E., Zhang G., Scofield R.P., Holdaway R.N., Bunce M. The half-life of DNA in bone: measuring decay kinetics in 158 dated fossils // Proc. R. Soc. B. 2012. V. 279. P. 4724-4733.
Annaluru N. et al. Total synthesis of a functional designer eukaryotic chromosome // Science. 2014. V. 344. № 6179. P. 55-59.
Appleton R.D., Palmer A.R. Water-borne stimuli released by predatory crabs and damaged prey induce more predator-resistant shells in a marine gastropod // Proc Natl Acad Sci USA. 1988. V. 85. P. 4387-4391.
Asara J.M., Garavelli J.S., Slatter D.A., Schweitzer M.H., Freimark L.M., Phillips M., Cantley L.C. (б) Interpreting sequences from mastodon and T. rex // Science. 2007. V. 317. № 5843. P. 1324–1325.
Asara J.M., Schweitzer M.H., Freimark L.M., Phillips M., Cantley L.C. (a) Protein sequences from mastodon and Tyrannosaurus rex revealed by mass spectrometry // Science. 2007. V. 316. № 5822. P. 280–285.
Atluri J.B., Samatha B., Bhupathi Rayalu M. Life history, phenology, host plant selection and utilization in the endangered danaid eggfly Hypolimnas misippus // Int. J. Adv. Res. Technol. 2013. V. 2. № 3. P. 117-126.
Attwater J., Wochner A., Holliger P. In-ice evolution of RNA polymerase ribosyme activity // Nature chemistry. 2013. V. 5. P. 1011-1018.
Barluenga M., Stolting K.N., Salzburger W., Muschick M., Meyer A. Sympatric speciation in Nicaraquan crater lake cichlid fish // Nature. 2006. V. 439. P. 719-723.
Beckers O.M., Wagner W.E. Jr. Eavesdropping parasitoids do not cause the evolution of less conspicuous signaling behaviour in a field cricket // Animal Behaviour. 2012. V. 84. P. 1457-1462.
Behe M.J. Darwin’s Black Box: The biochemical challenge to evolution. The Free Press, New York. 1996. - 307 p.
Behm J.E., Ives A.R., Boughman J.W. Breakdown in postmating isolation and the collapse of species pair through hybridization // The American Naturalist. 2010. V. 175. № 1. P. 11-26.
Berlocher S.H., Enquist M. Distribution and host plants of the apple maggot fly, Rhagoletis pomonella (Diptera: Tephritidae) in Texas // Journal of the Kansas entomological society. 1993. V. 66. № 1. P. 51-99.
Bertazzo S., Maidment S.C.R., Kallepitis C., Fearn S., Stevens M.M., Xie H. Fibres and cellular structures preserved in 75-million-year-old dinosaur specimens // Nature Communications 6. 2015. Article number: 7352. doi: 10.1038/ncomms8352
Bhullar K., Waglechner N., Pawlowski A., Koteva K., Banks E.D., Johnston M.D., Barton H.A., Wright G.D. Antibiotic resistance is prevalent in an isolated cave microbiome // PLOS ONE. 2012. V. 7. № 4. P. e34953.
Blount Z.D., Barrick J.E., Davidson C.J., Lenski R.E. Genomic analysis of a key innovation in an experimental Escherichia coli population // Nature. 2012. V. 489. P. 513-518.
Blount Z.D., Borland C.Z., Lenski R.E. Historical contingency and the evolution of a key innovation in an experimental population of Escherichia coli // PNAS. 2008. V. 105. № 23. P. 7899-7906.
Boyce D.G., Lewis M.R., Worm B. Global phytoplankton decline over the past century // Nature. 2010. V. 466. P. 591–596.
Brown G., Kelehear C., Shine R. The early toad gets the worm: cane toads at an invasion front benefit from higher prey availability // Journal of Animal Ecology. 2013. V. 82. P. 854-862.
Brown G.P., Phillips B.L., Webb J.K., Shine R. Toad on the road: use of roads as dispersal corridors by cane toads (Bufo marinus) at an invasion front in tropical Australia // Biological Conservation. 2006. V. 133. P. 88-94.
Buckley M. et al. Comment on “Protein sequences from Mastodon and Tyrannosaurus rex revealed by mass spectrometry” // Science. 2008. V. 319. № 5859, p. 33.
Bush G.L. The taxonomy, cytology, and evolution of the genus Rhagoletis in North America (Diptera: Tephritidae). Bull Mus Comp Zool. 1966. V. 134. P. 431–562.
Cameron E.Z., Du Toit J.T. Social influences on vigilance behaviour in giraffes, Giraffa camelopardalis // Animal Behaviour. 2005. V. 69. P. 1337–1344.
Cameron E.Z., Du Toit J.T. Winning by a neck: tall giraffes avoid competing with shorter browsers // American Naturalist. 2007. V. 169. № 1. P. 130–135.
Cameron P.J., Morrison F.O. Analysis of mortality in the apple maggot, Rhagoletis pomonella (Diptera: Tephritidae), in Quebec // The Canadian Entomologist. 1977. V. 109. № 6. P. 769-788.
Cano R.J., Poinar H., Poinar G.O., Jr. (a) Isolation and partial characterization of DNA from the bee Proplebeia dominicana (Apidae: Hymenoptera) in 25-40 million year old amber // Medical Science Research. 1992. V. 20. № 7. P. 249-251.
Cano R.J., Poinar H.N., Pieniazek N.J., Acra A., Poinar G.O. Jr. Amplification and sequencing of DNA from a 120-135-million-year-old weevil // Nature. 1993. V. 363. № 6429. P. 536-538.
Cano R.J., Poinar H.N., Roubik D.W., Poinar G.O. Jr. (б) Enzymic amplification and nucleotide sequencing of portions of the 18s rRNA gene of the bee Proplebeia dominicana (Apidae: Hymenoptera) isolated from 25-40 million year old Dominican amber // Medical Science Research. 1992. V. 20. № 17. P. 619-622.
Clemente J.C., Pehrsson E.C., Blaser M.J., Sandhu K., Gao Z., Wang B., Magris M., Hidalgo G., Contreras M., Noya-Alarcon O., Lander O., McDonald J., Cox M., Walter J., Oh P.L., Ruiz J.F., Rodriquez S., Shen N., Song S.J., Metcalf J., Knight R., Dantas G., Dominquez-Bello M.G. The microbiome of uncontacted Amerindians // Science Advances. 2015. V. 1. № 3. P. e1500183
Cody G.D., Gupta N.S., Briggs D.E.G., Kilcoyne A.L.D., Summons R.E., Kenig F., Plotnick R.E., Scott A.C. Molecular signature of chitin-protein complex in Paleozoic arthropods // Geology. 2011. V. 39. № 3. P. 255-258. doi: 10.1130/G31648.1
Collins M.J., Gernaey A.M., Nielsen-Marsh C.M., Vermeer C., Westbroek P. Osteocalcin in fossil bones: evidence of very slow rates of decomposition from laboratory studies // Geology. 2000. V. 28. P. 1139-1142.
Colombo M., Diepeveen E.T., Muschick M., Santos M.E., Indermaur A., Boileau N., Barluenga M., Salzburger W. The ecological and genetic basis of convergent thick-lipped phenotypes in cichlid fishes // Mol Ecol. 2013. V. 22. № 3. P. 670-684.
Crothers J.H. Dog-whelks: an introduction to the biology of Nucella lapillus (L.) // Field Studies. 1985. V. 6. P. 291-360.
D’Costa V.M., King C.E, Kalan L., Morar M., Sung W.W.L., Schwarz C., Froese D., Zazula G., Calmels F., Debruyne R., Golding G.B., Poinar H.N., Wright G.D. Antibiotic resistance is ancient // Nature. 2011. V. 477. P. 457–461.
Dakin R., Montgomerie R. Peahens prefer peacocks displaying more eyespots, but rarely // Animal Behaviour. 2011. V. 82. № 1. P. 21-28.
Davies K.L. Duck-bill dinosaurs (Hadrosauridae, Ornithischia) from the north slope of Alaska // Journal of Paleontology. 1987. V. 61. № 1. P. 198-200.
Dawkins R. The extended phenotype. Oxford University Press, 1982. - 307 p.
DeSalle R., Gatesy J., Wheeler W., Grimaldi D. DNA sequences from a fossil termite in Oligo-Miocene amber their phylogenetic implications // Science. 1992. V. 257. № 5078. P. 1933-1936.
Dias B.G., Ressler K.J. Parental olfactory experience influences behavior and neural structure in subsequent generations // Nature Neuroscience. 2013. V. 17. P. 89-96.
Diegisser T., Seitz A., Johannesen J. Morphological adaptation in host races of Tephritis conura // Entomologia Experimentalis et Applicata. 2007. V. 122. № 2. P. 155-164.
Dres M., Mallet J. Host races in plant-feeding insects and their importance in sympatric speciation. Philosophical Transactions of the Royal Society of London B. 2002. V. 357. P. 471– 492.
Du Toit J.T. Feeding-height stratification among African browsing ruminants // African Journal of Ecology. 1990. V. 28. P. 55–61.
Edgell T.C. The geography of inducible defence in a marine snail // The Malacologist. 2008. V. 50. P. 30-31. http://www.malacsoc.org.uk/the_Malacologist/BULL50/Edgell.htm
Edgell T.C., Neufeld C.J. Experimental evidence for latent developmental plasticity: intertidal whelks respond to a native but not an introduced predator // Biol Lett. 2008. V. 4. P. 385-387.
Edwards N.P., Barden H.E., van Dongen B.E., Manning P.L., Larson P.L., Bergmann U., Sellers W.I., Wogeleus R.A. Infrared mapping resolves soft tissue preservation in 50 million year-old reptile skin // Proc. R. Soc. B. 2011. V. 278. P. 3209-3218.
Elmer K.R., Lehtonen T.K., Kautt A.F., Harrod C., Meyer A. Rapid sympatric ecological differentiation of crater lake cichlid fishes within historic times // BMC Biology. 2010. V. 8. P. 60.
Eronen M., Zetterberg P., Briffa K.R., Lindholm M., Merilainen J., Timonen M. The supra-long Scots pine tree-ring record for Finnish Lapland: part 1, chronology construction and initial inferences // The Holocene. 2002. V. 12. P. 673-680.
Feder J.L. The effects of parasitoids on sympatric host races of Rhagoletis pomonella (Diptera: Tephritidae) // Ecology. 1995. V. 76. № 3. P. 801-813.
Feder J.L., Reynolds K., Go W., Wang E.C. Intra- and interspecific competition and host race formation in the apple maggot fly, Rhagoletis pomonella (Diptera: Tephritidae) // Oecologia. 1995. V. 101. № 4. P. 416-425.
Fikacek M., Prokin A., Angus R.B. A long-living species of the hydrophiloid beetles: Helophorus sibiricus from the early Miocene deposits of Kartashevo (Siberia, Russia) // Zookeys. 2011. V. 130. P. 239-254.
Filchak K.E., Roethele J.B., Feder J.L. Natural selection and sympatric divergence in the apple maggot Rhagoletis pomonella // Nature. 2000. V. 407. №. 6805. P. 739-742.
Fisher J.A.D., Rhile E.C., Liu H., Petraitis P.S. An intertidal snail shows a dramatic size increase over the past century // Proc Natl Acad Sci USA. 2009. V. 106. P. 5209-5212.
Forbes A.A., Powell T.H.Q., Stelinski L.L., Smith J.J., Feder J.L. Sequential sympatric speciation across trophic levels // Science. 2009. V. 323. № 5915. P. 776-779.
Freeman A.S., Byers J.E. Divergent induced responses to an invasive predator in marine mussel populations // Science. 2006. V. 313. P. 831-833.
Freeman A.S., Byers J.E. Response to comment on “Divergent induced responses to an invasive predator in marine mussel populations” // Science. 2007. V. 316. P. 53.
Gallant J.R., Traeger L.L., Volkening J.D., Moffett H., Chen P., Novina C.D., Phillips Jr. G.N., Anand R., Wells G.B., Pinch M., Guth R., Unguez G.A., Albert J.S., Zakon H.H., Samanta M.P., Sussman M.R. Genomic basis for the convergent evolution of electric organs // Science. 2014. V. 344. P. 1522–1525.
Garwood R.J., Dunlop J.A., Giribet G., Sutton M.D. Anatomically modern Carboniferous harvestmen demonstrate early cladogenesis and stasis in Opiliones // Nature Communications 2. 2011. Article number: 444. doi:10.1038/ncomms1458
Gibson D.G., Benders G.A., Andrews-Pfannkoch C., Denisova E.A., Baden-Tillson H., Zaveri J., Stockwell T.B., Brownley A., Thomas D.W., Algire M.A., Merryman C., Young L., Noskov V.N., Glass J.I., Venter J.C., Hutchison III C.A., Smith H.O. Complete chemical synthesis, assembly, and cloning of a Mycoplasma genitalium genome // Science. 2008. V. 319. № 5867. P. 1215-1220.
Glass K., Ito S., Wilby P.R., Sota T., Nakamura A., Bowers C.R., Vinther J., Dutta S., Summons R., Briggs D.E.G., Wakamatsu K., Simon J.D. Direct chemical evidence for eumelanin pigment from the jurassic period // PNAS. 2012. V. 109. № 26. P. 10218-10223.
Golenberg E.M., Giannasi D.E., Clegg M.T., Smiley C.J., Durbin M., Henderson D., Zurawski G. Chloroplast DNA sequence from a Miocene Magnolia species // Nature. 1990. V. 344. P. 656-658.
Gore P.J.W. Triassic Notostracans in the newark supergroup, Culpeper Basin, Northern Virginia // Journal of Paleontology. 1986. V. 60. № 5. P. 1086-1096.
Gould S.J. Evolution's erratic pace // Natural History. 1977. V. 86. № 5. P. 14.
Grant B.R., Grant P.R. Evolution of Darwin’s finches caused by a rare climatic event // Proc. R. Soc. Lond. B. 1993. V. 251. P. 111–117.
Grant B.R., Grant P.R. Natural selection in a population of Darwin’s finches // Am. Nat. 1989. V. 133. P. 377–393.
Grant P.R., Grant B.R. Predicting microevolutionary responses to directional selection on heritable variation // Evolution. 1995. V. 49. P. 241–251.
Grant P.R., Grant B.R. Unpredictable evolution in a 30-year study of Darwin’s finches // Science. 2002. V. 296. P. 707–711.
Greenblatt C.L., Baum J., Klein B.Y., Nachshon S., Koltunov V., Cano R.J. Micrococcus luteus – survival in amber // Microb Ecol. 2004. V. 48. № 1. P. 120-127.
Hairston N.G., Ellner S.P., Geber M.A., Takehito Y., Fox J.A. Rapid evolution and the convergence of ecological and evolutionary time // Ecology letters. 2005. V. 8. P. 1114-1127.
Hall B.G. Adaptive evolution that requires multiple spontaneous mutations: mutations involving base substitutions // Proc. Natl. Acad. Sci. USA. 1991. V. 88. P. 5882-5886.
Harris M.P., Hasso S.M., Ferguson M.W.J., Fallon J.F. The development of Archosaurian first-generation teeth in a chicken mutant // Current Biology. 2006. V. 16. № 4. P. 371-377.
Hebsgaard M.B., Phillips M.J., Willerslev E. Geologically ancient DNA: fact or artifact? // Trends in Microbiology. 2005. V. 13. № 5. P. 212-220.
Hegna T.A., Ren D. Two new «notostracans», Chenops gen. nov. and Jeholops gen. nov. (Crustacea: Branchiopoda: ?Notostraca) from the Yixian Formation, Northeastern China // Acta Geologica Sinica. 2010. V. 84. № 4. P. 886–894.
Herrel A., Huyghe K., Vanhooydonck B., Backeljau T., Breugelmans K., Grbac I., Van Damme R., Irschick D.J. Rapid large-scale evolutionary divergence in morphology and performance associated with exploitation of different dietary resource // PNAS. 2008. V. 105. № 12. P. 4792-4795.
Hopkins R., Rausher M.D. Pollinator-mediated selection on flower color allele drives reinforcement // Science. 2012. V. 335. P. 1090-1092.
Husak J.F. Ecological and evolutionary significance of locomotor performance in collared lizards (Crotaphytus collaris). PhD Dissertation. Oklahoma State University, Stillwater, Oklahoma. 2005.
Husak J.F. Does speed help you survive? A test with collared lizards of different ages // Functional Ecology. 2006. V. 20. P. 174-179.
Husak J.F., Ribak G., Wilkinson G.S., Swallow J.G. Compensation for exaggerated eye stalks in stalk-eyed flies (Diopsidae) // Functional Ecology. 2011. V. 25. P. 608-616.
Jahren A.H., Sternberg L.S.L. Eocene meridional weather patterns reflected in the oxygen isotopes of arctic fossil wood // GSA Today. 2002. V. 12. № 1. P. 1-9.
Jenkin F. The origin of species. Art. I. // North Brit. Rev. 1867. V. 46. P. 277-318.
Jones A.G., Ratterman N.L. Mate choice and sexual selection: What have we learned since Darwin? // PNAS. 2009. V. 106. № 1. P. 10001–10008.
Jones Z.M., Jayne B.C. Perch diameter and branching patterns have interactive effects on the locomotion and path choice of anole lizards // The Journal of Experimental Biology. 2012. V. 215. P. 2096-2107.
Kameneva E.P. A new species of the genus Plagiocephalus (Diptera, Ulidiidae) from Central America // Vestnik zoologii. 2004. V. 38. № 4. P. 15-22.
Kaye T.G., Gaugler G., Sawlowicz Z. Dinosaurian soft tissues interpreted as bacterial biofilms // PLoS ONE. 2008. V.3. № 7. P. e2808.
Kelber K.-P. New Triopsids (Crustacea, Notostraca) from the Upper Triassic of Frankonia, Germany / Epicontinental Triassic International Symposium. Hallesches Jb Geowissenschaften Beiheft. 1998. V.5. P 85.
Kim S., Soltis D.E., Soltis P.S., Suh Y. DNA sequences from Miocene fossils: an ndhF sequence of Magnolia latahensis (Magnoliaceae) and an rbcL sequence of Persea pseudocarolinensis (Lauraceae) // Am. J. Bot. 2004. V. 91. № 4. P. 615-620.
Klutsch C.F.C., Misof B., Grosse W.-R., Moritz R.F.A. Genetic and morphometric differentiation among island populations of two Norops lizards (Reptilia: Sauria: Polychrotidae) on independently colonized islands of the Islas de Bahia (Honduras) // Journal of Biogeography. 2007. V. 34. P. 1124-1135.
Kolbe J.J., Leal M., Schoener T.W., Spiller D.A., Losos J.B. Founder effects persist despite adaptive differentiation: a field experiment with lizards // Science. 2012. V. 335. P. 1086–1089.
Larralde R., Robertson M.P., Miller S.L. Rates of decomposition of ribose and other sugars: implications for chemical evolution // Proc. Natl. Acad. Sci. USA. 1995. V. 92. P. 8158-8160.
Lee M.S.Y., Jago J.B., Garcia-Bellido D.C., Edgecombe G.D., Gehling J.G., Paterson J.R. Modern optics in exceptionally preserved eyes of Early Cambrian arthropods from Australia // Nature. 2011. V. 474. P. 631-634.
Leuschner H.H., Sass-Klaassen U., Jansma E., Baillie M.G.L., Spurk M. Subfossil European bog oaks: population dynamics and long-term growth depressions as indicators of changes in the Holocene hydro-regime and climate // The Holocene. 2002. V. 12. P. 695-706.
Leuthold B.M., Leuthold W. Food habits of giraffe in Tsavo National Park, Kenya // East African Wildlife Journal. 1972. V. 10. P. 129–141.
Levy M., Miller S.L. The stability of the RNA bases: implications for the origin of life // Proc. Natl. Acad. Sci. USA. 1998. V. 95. P. 7933-7938.
Liebers D., de Knijff P., Helbig A.J. The herring gull complex in not a ring species // Proc Biol Sci. 2004. V. 271. № 1542. P. 893-901.
Lindgren J., Uvdal P., Engdahl A., Lee A.H., Alwmark C., Bergquist K-E., Nillson E., Ekstrom P., Rasmussen M., Douglas D.A., Polcyn M.J., Jacobs L.L. Microspectroscopic evidence of cretaceous bone proteins // PLoS ONE. 2011. V. 6. № 4. P. e19445.
Lindstrom T., Brown G.P., Sisson S.A., Phillips B.L., Shine R. Rapid shifts in dispersal behavior on an expanding range edge // PNAS. 2013. V. 110. № 33. P. 13452-12456.
Linn C., Dambroski H.R., Feder J.L., Berlocher S.H., Nojima S., Roelofs W.L. Postzygotic isolating factor in sympatric speciation in Rhagoletis flies: Reduced response of hybrids to parental host-fruit odors // PNAS. 2004. V. 101. № 51. P. 17753-17758.
Linn C., Feder J.L., Nojima S., Dambroski H.R., Berlocher S.H., Roelofs W. Fruit odor discrimination and sympatric host race formation in Rhagoletis // PNAS. 2003. V. 100. № 20. P. 11490-11493.
Llewellyn D., Thompson M.B., Phillips B.L., Brown G.P., Shine R. Reduced investment in immune function in invasion-front populations of the cane toad (Rhinella marina) in Australia // Biological Invasions. 2012. V. 14. P. 999-1008.
Logan M.L., Montgomery C.E., Boback S.M., Reed R.N., Campbell J.A. Divergence in morphology, but not habitat use, despite low genetic differentiation among insular populations of the lizard Anolis lemurinus in Honduras // Journal of Tropical Ecology. 2012. V. 28. P. 215-222.
Losos J.B., Schoener T.W., Langerhans R.B., Spiller D.A. Rapid temporal reversal in Predator-Driven Natural Selection // Science. 2006. V. 314. № 5803. P. 1111.
Loyau A., Jalme M.S., Sorci G. Intra- and intersexual selection for multiple traits in the peacock (Pavo cristatus) // Ethology. 2005. V. 111. P. 810-820.
Luck N., Joly D. Sexual selection and mating advantages in the giant sperm species, Drosophila bifurca // Journal of Insect Science. 2005. V. 5. P. 10.
Ma X., Cong P., Hou X., Edgecombe G.D., Strausfeld N.J. An exceptionally preserved arthropod cardiovascular system from the early Cambrian // Nature Communications 5. 2014. Article number: 3560. doi: 10.1038/ncomms4560.
Ma X., Hou X., Edgecombe G.D., Strausfeld N.J. Complex brain and optic lobes in an early Cambrian arthropod // Nature. 2012. V. 490. P. 258-261.
Maisnier-Patin S., Roth J.R., Fredriksson A., Nystrom T., Berg O.G., Andersson D.I. Genomic buffering mitigates the effects of deleterious mutations in bacteria // Nature Genetics. 2005. V. 37. № 12. P. 1376-1379.
Manning P.L., Morris P.M., McMahon A., Jones E., Gize A., Macquaker J.H.S., Wolff G., Thompson A., Marshall J., Taylor K.G., Lyson T., Gaskell S., Reamtong O., Sellers W.I., van Dongen B.E., Buckley M., Wogelius R.A. Mineralized soft-tissue structure and chemistry in a mummified hadrosaur from the Hell Creek Formation North Dakota (USA) // Proc. R. Soc. B. 2009. doi:10.1098/rspb.2009.0812
McHenry C.R., Wroe S., Clausen P.D., Moreno K., Cunningham E. Supermodeled sabercat, predatory behavior in Smilodon fatalis revealed by high-resolution 3D computer simulation // PNAS. 2007. V. 104. № 41. P. 16010-16015.
Mitchell G., Van Sittert S., Skinner J.D. The demography of giraffe deaths in a drought // Transactions of Royal Society of South Africa. 2010. V. 65. № 3. P. 165-168.
Mitchell G., Van Sittert S.J., Skinner J.D. Sexual selection is not the origin of long necks in giraffes // Journal of Zoology. 2009. V. 278. № 4. P. 281-286.
Moczydlowska M., Westall F., Foucher F. Microstructure and Biogeochemistry of the Organically Preserved Ediacaran Metazoan Sabellidites // Journal of Paleontology. 2014. V. 88. № 2. P. 224-239.
Nagl S., Tichy H., Mayer W.E., Takezaki N., Takahata N., Klein J. The origin and age of haplochromine fishes in Lake Victoria, East Africa // Proc. R. Soc. Lond. B. 2000. V. 267. P. 1049-1061.
Nielsen-Marsh C. Biomolecules in fossil remains. Multidisciplinary approach to endurance // The Biochemist (Journal of The Biochemical Society). 2002. P. 12–14.
O'Malley C.E. Crinoid biomarkers (borden Group, Mississippian): implications for phylogeny. Ohio State University. 2006. 122 p.
Packert M., Martens J., Eck S., Nazarenko A.A., Valchuk O.P., Petri B., Veith M. The great tit (Parus major) – a misclassified ring species // Bilogical Journal of the Linnean Society. 2005. V. 86. № 2. P. 153-174.
Palmer J., Lorrey A., Turney C.S.M., Hogg A., Baillie M., Fifield K., Ogden J. Extension of New Zealand kauri (Agathis australis) tree-ring chronologies into Oxygen Isotope Stage (OIS)-3 // Journal of Quaternary Science. 2006. V. 21. P. 779-787.
Park J.S., Vreeland R.H., Cho B.C., Lowenstein T.K., Timofeeff M.N., Rosenzweig W.D. Haloarchaeal diversity in 23, 121 and 419 MYA salts // Geobiology. 2009. V. 7. № 5. P. 515-523.
Pascoal S., Carvalho G., Creer S., Mendo S., Hughes R. Plastic and heritable variation in shell thickness of the intertidal gastropod Nucella lapillus associated with risks of crab predation and wave action, and sexual maturation // PLoS ONE. 2012. V.7. № 12. P. e52134.
Pascoal S., Cezard T., Eik-Nes A., Gharbi K., Majewska J., Payne E., Ritchie M.G., Zuk M., Bailey N.W. Rapid convergent evolution in wild crickets // Current Biology. 2014. V. 24. P. 1-6.
Patel B.H., Percivalle C., Ritson D.J., Duffy C.D., Sutherland J.D. Common origins of RNA, protein and lipid precursors in cyanosulfidic protometabolism // Nature Chemistry. 2015. V. 7. P. 301-307.
Paterson J.R., Garcia-Bellido D.C., Lee M.S.Y., Brock G.A., Jago J.B., Edgecombe G.D. Acute vision in the giant Cambrian predator Anomalocaris and the origin of compound eyes // Nature. 2011. V. 480. P. 237-240.
Pawlicki H., Korbel A., Kubiak H. Cells, collagen fibrils and vessels in dinosaur bone // Nature. 1966. V. 211. № 49. P. 655–657.
Pawlicki R. Histochemical demonstration of DNA in osteocytes from dinosaur bones // Folia Histochem. Cytobiol. 1995. V. 33. № 3. P. 183–186.
Pawlicki R. Metabolic pathways of the fossil dinosaur bones. Part II. Vascular canal in the communication system // Folia Histochem. Cytobiol. (Krakow). 1984. V. 22. № 1. P. 33–41.
Pawlicki R., Nowogrodzka-Zagorska M. Blood vessels and red blood cells preserved in dinosaur bones // Anat. Anz. 1998. V. 180. № 1. P. 73–77.
Pellew R.A. The feeding ecology of a selective browser, the giraffe (Giraffa camelopardalis tippelskirchi) // Journal of Zoology (London). 1984. V. 202. P. 57–81.
Pevzner P.A., Kim S., Ng J. Comment on «Protein Sequences from Mastodon and Tyrannosaurus rex Revealed by Mass Spectrometry» // Science. 2008. V. 319. № 5892, P. 1040.
Pfenning A.R. et al. Convergent transcriptional specializations in the brains of humans and song-learning birds // Science. 2014. V. 346. № 6215. P. 1256846. doi:10.1126/science.1256846.
Phillips B.L., Brown G.P., Greenlees M., Webb J.K., Shine R. 2007. Rapid expansion of the cane toad (Bufo marinus) invasion front in tropical Australia // Austral Ecology. 2007. V. 32. P. 169-176.
Phillips B.L., Brown G.P., Webb J.K., Shine R. Invasion and the evolution of speed in toads // Nature. 2006. V. 439. P. 803.
Poinar H.N., Cano R.J., Poinar G.O. Jr. DNA from an extinct plant // Nature. 1993. V. 363. P. 677.
Poinar Jr. G.O., Marshall C.J., Buckley R. One hundred million years of chemical warfare by
insects // Journal of Chemical Ecology. 2007. V. 33. № 9. P. 1663-1669.
Pose D., Verhage L., Ott F., Yant L., Mathieu J., Angenent G.C., Immink R.G.H., Schmid M. Temperature-dependent regulation of flowering by antagonistic FLM variants // Nature. 2013. V. 503. P. 414–417.
Powner M.W, Gerland B., Sutherland J.D. Synthesis of activated pyrimidine ribonucleotides in prebiotically plausible conditions // Nature. 2009. V. 459. P. 239–242.
Prokopy R.J., Diehl S.R., Coolcy S.S. Behavioral evidence for host races in Rhagoletis pomonella flies // Oecologia. 1988. V. 76. P. 138-147.
Rajakumar R., Sun Mauro D., Dijkstra M.B., Huang M.H., Wheeler D.E., Hiou-Tim F., Khila A., Cournoyaea M., Abouheif E. Ancestral developmental potential facilitates parallel evolution in ants // Science. 2012. V. 335. № 6064. P. 79-82.
Rawson P.D., Yund P.O., Lindsay S.M. Comment on “Divergent induced responses to an invasive predator in marine mussel populations” // Science. 2007. V. 316. P. 53.
Reisz R.R., Huang T.D., Roberts E.M., Peng S., Sullivan C., Stein K., LeBlanc A.R.H., Shieh D., Chang R., Chiang C., Yang C., Zhong S. Embryology of early jarassic dinosaur from China with evidence of preserved organic remains // Nature. 2013. V. 496. P. 210-214.
Ricklefs R.E., Renner S.S. Global Correlations in Tropical Tree Species Richness and Abundance Reject Neutrality // Science. 2012. V. 335. P. 464–467.
Reznick D.N., Shaw F.H., Rodd H., Shaw R.G. Evaluation of the rate of evolution in natural populations of guppies (Poecilia reticulata) // Science. 1997. V. 275. № 5308. P. 1934-1937.
Roughgarden J., Akcay E. Do we need a Sexual Selection 2.0? // Animal behaviour. 2010. V. 79. P. e1-e4.
Sanford J.C., Baumgardner J., Brewer W., Gibson P., ReMine W. (а) Mendel's Accountant: a biologically realistic forward-time population genetics program // SCPE. 2007. V. 8. P. 147-165.
Sanford J.C., Baumgardner J., Brewer W., Gibson P., ReMine, W. (б) Using computer simulation to understand mutation accumulation dynamics and genetic load. / In: Computational Science - ICCS 2007. Ed. by: Shi et al. LNCS, Springer-Verlag, Berlin, Heidelberg, 2007. V. 4488. P. 386-392.
Schmidt A.R., Jancke S., Lindquist E.E., Ragazzi E., Roghi G., Nascimbene P.C., Schmidt K., Wappler T., Grimaldi D.A. Arthropods in amber from the Triassic period // PNAS. 2012. V. 109. № 37. P. 14796-14801.
Schopf J.W., Kudryavtsev A.B., Walter M.R., Kranendonk M.J.V., Williford K.H., Kozdon R., Valley J.W., Gallardo V.A., Espinosa C., Flannery D.T. Sulfur-cycling fossil bacteria from the 1.8-Ga Duck Creek Formation provide promising evidence of evolution’s null hypothesis // PNAS. 2015. V. 112, № 7. P. 2087-2092.
Schweitzer M.H., Horner J.R. Intravascular microstructures in trabecular bone tissues of Tyrannosaurus rex // Annales de Paleontologie. 1999. V. 85. № 3. P. 179–192.
Schweitzer M.H., Marshall M., Carron K., Bohle S., Busse S.C., Arnold E.V., Barnard D., Horner J.R., Starkey J.R. Heme compounds in dinosaur trabecular bone // Proc. Natl. Acad. Sci. U.S.A. 1997. V. 94. № 12. P. 6291–6296.
Schweitzer M.H., Suo Z., Avci R., Asara J.M., Allen M.A., Arce F.T., Horner J.R. Analyses of soft tissue from Tyrannosaurus rex suggest the presence of protein // Science. 2007. V. 316. № 5822. P. 277–280.
Schweitzer M.H., Wittmeyer J.L., Horner J.R. (б) Gender-specific reproductive tissue in ratites and Tyrannosaurus rex // Science. 2005. V. 308. № 5727. P. 1456–1460.
Schweitzer M.H., Wittmeyer J.L., Horner J.R., Toporski J.K. (а) Soft-Tissue Vessels and Cellular Preservation in Tyrannosaurus rex // Science. 2005. V. 307. № 5717. P. 1952–1955.
Schweitzer M.H., Zheng W., Cleland T.P., Bern M. Molecular analyses of dinosaur osteocytes support the presence of endogenous molecules // Bone. 2013. V. 52. № 1. P. 414-423.
Schweitzer M.H., Zheng W., Organ C.L., Avci R., Suo Z., Freimark L.M., Lebleu V.S., Duncan M.D., Vander Heiden M.G., Neveu J.M., Lane W.S., Cottrell J.S., Horner J.R., Cantley L.C., Kalluri R., Asara J.M. Biomolecular characterization and protein sequences of the campanian hadrosaur B. canadensis // Science. 2009. V. 324. № 5927. P. 626–631.
Seeley R.H. Intense natural selection caused a rapid morphological transition in a living marine snail // Proc Natl Acad Sci USA. 1986. V. 83. P. 6897-6901.
Sergievski S.O., Granovitch A.I., Sokolova I.M. Long-term studies of Littorina obtusata and Littorina saxatilis populations in the White Sea // Oceanologica Acta. 1997. V. 20. № 1. P. 259-265.
Shapiro R. Prebiotic cytosine synthesis: a critical analysis and implications for the origin of life // Proc. Natl. Acad. Sci. USA. 1999. V. 96. P. 4396-4401.
Sherman M.Y. Universal genome in the origin of Metazoa: thoughts about evolution // Cell Cycle. 2007. V. 6. № 15. P. 1873-1877.
Shervis L.J., Boosh G.M., Koval C.F. Infestation of sour cherries by the apple maggot: confirmation of a previously uncertain host status // Journal of Economic Entomology. 1970. V. 63. № 1. P. 294-295.
Shilton C.M., Brown G.P., Benedict S., Shine R. Spinal arthropathy associated with Ochrobactrum anthropi in free-ranging cane toads (Chaunus [Bufo] marinus) in Australia // Veterinary Pathology. 2008. V. 45. P. 85-94.
Shine R., Brown G.P., Phillips B.L. (a) An evolutionary process that assembles phenotypes through space rather than time // PNAS. 2011. V. 108. № 14. P. 5708-5711.
Shine R., Brown G.P., Phillips B.L. (b) Spatial sorting, assortative mating, and natural selection // PNAS. 2011. V. 108. № 31. P. E348.
Simmons R.E., Scheepers L. Winning by a neck: sexual selection in the evolution of giraffe // The American Naturalist. 1996. V. 148. № 5. P. 771-786.
Smejkal G.B., Schweitzer M.H. Will current technologies enable dinosaur proteomics? // Expert Rev. Proteomics. 2007. V. 4. № 6. P. 695–699.
Sodera V. One small speck to man: the evolution myth. Vija Sodera Productions. 2009. - 560 p.
Soltis P.S., Soltis D.E., Smiley G.J. An rbcL sequence from a Miocene Taxodium (bald cypress) // Proc. Natl. Acad. Sci. USA. 1992. V. 89. P. 449–451.
Spirin A.S. When, where, and in what environment could the RNA world appear and evolve? // Paleontological Journal. 2007. V. 41. № 5. P. 481-488.
Stahle D.W., Burnette D.J, Villanueva J., Cerano J., Fye F.K., Griffin R.D., Cleaveland M.K., Stahle D.K., Edmondson J.R. Wolff K.P. Tree-ring analysis of ancient baldcypress trees and subfossil wood // Quaternary Science Reviews. 2012. V. 34. P. 1-15.
Stambaugh M.C., Guyette R.P. Progress in constructing a long oak chronology from the central United States // Tree-Ring Research. 2009. V. 65. P. 147-156.
Sutherland J.D. Ribonucleotides // Cold Spring Harbor Perspectives in Biology. 2010. V. 2. P. a005439.
Swallow J.S., Carter P.A., Garland T.Jr. Arttificial selection for increased wheel-running behavior in house mice // Behav Genet. 1998. V. 28. № 3. P. 227-237.
Schwartz A.W. Evaluating the plausibility of prebiotic multistage syntheses // Astrobiology. 2013. V. 13. № 8. P. 784-789.
Takahashi M., Arita H., Hiraiwa-Hasegawa M., Hasegawa T. Peahens do not prefer peacocks with more elaborate trains // Animal Behaviour. 2008. V. 75. № 4. P. 1209-1219.
Tanaka G., Hou X., Ma X., Edgecombe G.D., Strausfeld N.J. Chelicerate neural ground pattern in a Cambrian great appendage arthropod // Nature. 2013. V. 502. P. 364-367.
Tinghitella R.M., Zuk M., Beveridge M., Simmons L.W. Island hopping introduces Polynesian field crickets to novel environments, genetic bottlenecks and rapid evolution // Journal of Evolutionary Biology. 2011. V. 24. P. 1199-1211.
Trussell G.C., Nicklin M.O. Cue sensitivity, inducible defense, and trade-offs in a marine snail // Ecology. 2002. V. 83. P. 1635-1647.
Udintsev G.B. Equatorial Segment of the Mid-Atlantic Ridge: initial results of the geological and geophysical investigations under the EQUARIDGE Program, cruises of R/V 'Akademik Nikolaj Strakhov' in 1987, 1990, 1991. Unesco, 1996. 122 p.
Verheyen E., Salzburger W., Snoeks J., Meyer A. Origin of the superflock of cichlid fishes from lake Victoria, East Africa // Science. 2003. V. 300. № 5617. P. 325-329.
Vervust B., Grbac I., Van Damme R. Differences in morphology, performance and behaviour between recently diverged populations of Podarcis sicula mirror differences in predation pressure // Oikos. 2007. V. 116. № 8. P. 1343-1352.
Vreeland R.H., Rosenzweig W.D., Powers D.W. Isolation of a 250 million-year-old halotolerant bacterium from a primary salt crystal // Nature. 2000. V. 407, P. 897–900.
Weiner S., Lowenstam H.A., Hood L. Characterization of 80-million-year-old mollusk shell proteins // Proc. Natl. Acad. Sci. USA. 1976, V. 73. № 8. P. 2541-2545.
Weinreich D.M., Delaney N.F., DePristo M.A., Hartl D.L. Darwinian evolution can follow only very few mutational paths to fitter proteins // Science. 2006. V. 312. P. 111-114.
Westbroek P., van der Meide P.H., van der Wey-Kloppers, van der Sluis R.J., de Leeuw J.W., de Jong E.W. Fossil macromolecules cephalopod shells: characterization, immunological response and diagenesis // Paleobiology. 1979. V. 5. № 2. P. 151-167.
White A.W., Shine R. The extra-limital spread of an invasive species via ‘stowaway’ dispersal: toad to nowhere? // Animal Conservation. 2009. V. 12. P. 38-45.
Wilkinson G.S., Reillo P.R. Female choice response to artificial selection on an exaggerated male trait in a stalk-eyed fly // Proc. R. Soc. Lond. B. 1994. V. 255. P. 1-6.
Williams G.A. The comparative ecology of the flat periwinkles, Littorina obtusata (L.) and L. marinae Sacchi et Rastelli // Field Studies. 1990. V. 7. № 3. P. 469-482.
Wilson A.M., Lowe J.C., Roskilly K., Hudson P.E., Golabek K.A., McNutt J.W. Locomotion dynamics of hunting in wild cheetahs // Nature. 2013. V. 498. P. 185–189.
Wroe S., McHenry C., Thomason J. Bite club: comparative bite force in big biting mammals and the prediction of predatory behaviour in fossil taxa // Proc Biol Sci. 2005. V. 272. № 1563. P. 619-625.
Young T.P., Isbell L.A. Sex differences in giraffe feeding ecology: Energetic and social constraints // Ethology. 1991. V. 87. P. 79-89.
Zeh D.W., Zeh J.A., Tavakilian G. Sexual selection and sexual dimorphism in the harlequin beetle Acrocinus longimanus // Biotropica. 1992. V. 24. № 1. P. 86-96.
Zhao F., Bottjer D.J., Hu S., Yin Z., Zhu M. Complexity and diversity of eyes in Early Cambrian ecosystems // Scientific Reports 3. 2013. Article number: 2751. doi:10.1038/srep02751
Zierold T., Hanfling B., Gomez A. Recent evolution of alternative reproductive modes in the ‘living fossil’ Triops cancriformis // BMC Evol Biol. 2007. V. 7. P. 161.
Zuk M., Rotenberry J.T., Tinghitella R.M. Silent night: adaptive disappearance of a sexual signal in a parasitized population of field crickets // Biol. Lett. 2006. V. 2. P. 521-524.
Zuk M., Simmons L.W., Cupp L. Calling characteristics of parasitized and unparasitized populations of the field cricket Teleogryllus oceanicus // Behav Ecol Sociobiol. 1993. V. 33. P. 339-343.
Алтухов Ю.П., Рычков Ю.Г. Генетический мономорфизм видов и его возможное биологическое значение // Журнал общей биологии. 1972. Т. 33. №. 3. С. 281-300.
Андреева С.И., Андреев Н.И. Эволюционные преобразования двустворчатых моллюсков Аральского моря в условиях экологического кризиса. – Омск: Изд-во Омского государственного педагогического университета. 2003. - 382 с.
Анохин К.В., Бэйтсон П.П.Г., Бурцев М.С. Обучение, как решающий фактор эволюции функциональных систем / Современные проблемы биологической эволюции: материалы конференции. К 100-летию Государственного Дарвиновского музея. 17-20 сентября 2007, Москва. - М.: ГДМ. 2007. C. 221-223.
Антонов А.Ю. (а) Критический обзор представлений по главным геодинамическим направлениям современной геологической науки в контексте информативности основных рекламных научных изданий России (часть первая) // Вестник Краунц. Науки о Земле. 2007. № 1. Вып. 9. С. 133-144.
Антонов А.Ю. (б) Критический обзор представлений по главным геодинамическим направлениям современной геологической науки в контексте информативности основных рекламных научных изданий России (часть вторая) // Вестник Краунц. Науки о Земле. 2007. № 2. Вып.10. С. 118-127.
Белоусов В.В. Тектоносфера Земли: идеи и действительность. / В кн.: Проблемы глобальной тектоники. - М.: Наука. 1973. C. 60-96.
Биологический энциклопедический словарь / Гл. ред. М.С. Гиляров; редкол.: А.А. Баев, Г. Г. Винберг, Г.А. Заварзин и др. - М.: Сов. Энциклопедия. 1986. 831 с.
Голубева М., Опарина Н. Рожденные ползать могут и летать // «Троицкий вариант – Наука». 2011. №12(81). С. 8.
Гонтарева Н.Б. Синтез органических веществ - предшественников первых живых систем под влиянием физических факторов космической среды / Автореферат диссертации на соискание ученой степени кандидата физико-математических наук. - Санкт-Петербург. 2003. 21 с.
Грант В. Эволюционный процесс. - М.: Мир. 2008.
Докинз Р. Эгоистичный ген. - М.: АСТ:CORPUS. 2013. - 512 с.
Жирнов А.М. Мифы мобилизма и реальная тектоника // Отечественная геология. 2011. № 2. С. 103-107.
Заварзин Г.А. Становление биосферы // Вестник РАН. 2001, Т.71. № 11. C. 988-1001.
Караулов В.Б. Начало конца плейттектонической эпидемии? // Известия высших учебных заведений. Геология и разведка. 2005. № 1. С. 70-71.
Ковалев О.В. Формирование солитоноподобных волн при инвазиях организмов и в эволюции биосферы. / В кн.: Эволюционная биология. Т. 2. Материалы 2-ой Международной конференции «Проблема вида и видообразование». – Томск. 2001. С. 65-81.
Красилов В.А. Нерешенные проблемы теории эволюции. - Владивосток: ДВНЦ АН СССР. 1986. - 138 с.
Красилов В.А. Палеонтология и парадигмы современного естествознания // Экология и жизнь. 2009. № 5. С. 5-10.
Лунный А.Н. (а) Трудный путь признания тканей, сосудов, клеток и фрагментов коллагена в костях динозавров / В сб.: «Современное христианство и естественные науки: материалы докладов научно-богословского семинара», Кировск, 3–4 ноября 2009 г. - Апатиты: «К&М». 2009. С. 41–53. (имеется сетевая версия: http://www.goldentime.ru/hrs_text_040.htm)
Лунный А.Н. (б) ДНК и живые бактерии возрастом в «десятки – сотни миллионов лет» / В сб.: «Православное осмысление мiра и современная наука», Выпуск 5. Материалы XVII международных рождественских образовательных чтений. Отдел религиозного образования и катехизации Русской Православной Церкви. Миссионерско-Просветительский Центр «Шестодневъ». - М.: «НП МПЦ Шестодневъ». 2009. С. 139-182. (имеется сетевая версия: http://publ.lib.ru/ARCHIVES/L/LUNNYY_A._N/_Lunnyy_A.N..html#2009)
Лунный А.Н. Молекулярно-клеточная палеонтология на 2007 год: свидетельство о малом возрасте Земли: обзор / В кн.: Божественное откровение и современная наука. Альманах. Под ред. Н. Колчуринского. - М.: ООО «Три сестры». 2011. С. 98–159. (имеется сетевая версия: http://publ.lib.ru/ARCHIVES/L/LUNNYY_A._N/_Lunnyy_A.N..html )
Марков А. (а) Эволюционный эксперимент на ящерицах показал действенность «эффекта основателя». 2012. Взято с http://elementy.ru/news/431776
Марков А. (б) Параллельная эволюция касты «сверхсолдат» у муравьев получила объяснение. 2012. Взято с http://elementy.ru/news/431738
Марков А. Для видообразования не нужны барьеры. 2006. http://elementy.ru/news/430109
Марков А. (б) У микробов обнаружена способность к предвидению // 2009. http://elementy.ru/news/431107
Марков А. (а) Цепная реакция видообразования. 2009. Взято с http://elementy.ru/news/431001
Наймарк Е. Коллаген из костей динозавров – это уже реальность. 2007. http://elementy.ru/news/430500
Наймарк Е. Цихлиды - живая модель независимой параллельной эволюции. 2012. http://elementy.ru/news/431931
Наймарк Е. Электрические органы у разных групп рыб регулируются сходными генами. 2014. http://elementy.ru/news/432282
Определитель пресноводных беспозвоночных России и сопредельных территорий. Том 2. Ракообразные. / Под редакцией С.Я. Цалолихина. - Санкт-Петербург. 1995. - 629 с.
Расницын А.П. Процесс эволюции и методология систематики // Труды русского энтомологического общества. 2002. Т. 73. 107 с.
Тахтаджян А. Л. Флористические области Земли / Академия наук СССР. Ботанический институт им. В.Л. Комарова. - Л.: Наука, Ленинградское отделение. 1978. - 247 с.
Федоров В.В., Формозов Н.А., Сурин В.Л., Вальчук О.П., Керимов А.Б. Генетические последствия гибридизации большой Parus major и восточной P. minor синиц в Среднем Приамурье // Зоологический журнал. 2006. Т. 85. № 5. С. 621-629.
Фролов В.Т. Наука геология: философский анализ. - М.: Изд-во МГУ. 2004. - 128 с.
Шабанов Д. Обезьяны, догмы и «кошмар Дженкина» // Компьютерра. 2006. № 17(637). С. 44-46.
Шабанов Д. Королева эволюционной биологии: кошмар Дженкина возвращается // Компьютерра. 2014. Онлайн-публикация. Взято 10.11.2014 с сайта: http://www.computerra.ru/91153/koroleva-problem-evolyutsionnoy-biologii-koshmar-dzhenkina-vozvrashhaetsya/
Шапиро Р. У истоков жизни // В мире науки. 2007. № 10. С. 21-29.
Шапошников Г.Х. Морфологическая дивергенция и конвергенция в эксперименте с тлями (Homoptera, Aphidinea) // Энтом. Обозр. 1965. Т. 44. № 1. С. 3-5.
Шапошников Г.Х. Специфичность и возникновение адаптации к новым хозяевам у тлей (Homoptera, Aphidoidea) в процессе естественного отбора (экспериментальное исследование) // Энтом. Обозр. 1961. Т. 40. № 4. С. 739-762.
Шаталкин А.И. Высший уровень деления в классификации организмов. 2. Архебактерии, эубактерии и эукариоты // Журнал общей биологии. 2004. Т.65. № 2. C. 99-115.
[1] http://www.ncbi.nlm.nih.gov/pmc/articles/PMC386617/
[2] На самом деле, упомянутые дарвинистом ссылки на «эволюцию бактерий» здесь не подходили, потому что я просил установленный пример изменения морфологического признака, а не абы какого (кроме того, про «эволюцию бактерий» см. ниже). А пример с «эволюцией мышей» был вообще ужасен - в качестве «установленного факта изменений морфологического признака в природе», верующий дарвинист привел мне исследование, в котором проводился искусственный отбор (!) мышей (Swallow et al., 1998).
[3] http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2663991/
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3521725/
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC280434/
[4] Улитки Littorina obtusata обычно достигают половой зрелости на второй год своей жизни (Williams, 1990). А для Белого моря (вероятно, более холодного?) для этого вида вообще указывается срок достижения половозрелости 3-4 года (Sergievski et al., 1997).
[5] Рисунок взят с сайта лондонского общества малакологов. Режим доступа: http://www.malacsoc.org.uk/the_Malacologist/BULL50/Edgell.htm
[6] Рисунок со свободным доступом, взят с сайта https://commons.wikimedia.org/wiki/File:Nucella_Lapillus.jpg? (автор и лицензия указаны на сайте).
[7] Рисунок со свободным доступом, взят с сайта википедии (автор и лицензия указаны на сайте): https://en.wikipedia.org/wiki/Dog_whelk#/media/File:Nucella_lapillus.jpg
[8] Фотографии слизней взяты с сайта www.pixabay.com (свободный доступ).
[9] Здесь имеются в виду знаменитые «бактерии, научившиеся есть цитрат», в экспериментальных работах коллектива Ленски (Blount et al., 2008; Blount et al., 2012). Эти исследования получили чрезвычайную популярность в среде верующих дарвинистов. Поэтому мы обязательно поговорим о них подробно чуть позже (в разделе про «эволюцию бактерий»).
[10] Это тест, экспериментальным путем демонстрирующий случайный характер мутаций. Заключается он в том, что при воздействии антибиотика на колонии бактерий, число колоний, оказавшихся устойчивыми к антибиотику, очень сильно (непредсказуемо) варьирует в разных сериях экспериментов. Этот сильный разброс объясняется случайным характером соответствующих мутаций.
[11] Richard Dawkins (1982). Книга «The extended phenotype» (Расширенный фенотип), глава 3 «Ограничения на совершенствование», раздел «Запаздывание по времени».
[12] Гликолиз - метаболический путь, то есть, биохимический «конвейер», включающий в себя десяток отдельных этапов, где каждая отдельная стадия немыслима без предыдущей и в свою очередь нужна для следующей. Этот биохимический конвейер чрезвычайно широко распространен в живой природе (у самых разных организмов), являясь одним из основных (из трех имеющихся) путей окисления глюкозы живыми организмами. У некоторых организмов могут наблюдаться определенные вариации этого биохимического пути, однако не известны такие варианты окисления глюкозы, которые были бы существенно проще гликолиза.
[13] С легкой руки Майкла Бихи (Behe, 1996).
[14] http://www.malacsoc.org.uk/the_Malacologist/BULL50/Edgell.htm
[15] Глава 9. «Эволюция современных двустворчатых моллюсков Аральского моря» (Андреева, Андреев, 2003).
[16] Водные растения сравнительно большого размера.
[17] В работе Андреевых такое взаимодействие моллюсков озвучено, скорее, в виде предположения - по аналогии с другими моллюсками из соседнего Каспия.
[18] Данный вид ракушек был внедрён в Аральское море из лиманов Азовского моря (Андреева, Андреев, 2003).
[19] Например, известный рекордсмен в этом отношении, рыба-луна, мечет около 300 млн. икринок.
[20] http://paleoforum.ru/index.php/topic,2231.msg48455.html#msg48455
[21] http://paleoforum.ru/index.php/topic,168.msg5513.html#msg5513
[22] http://paleoforum.ru/index.php/topic,168.msg11221.html#msg11221
[23] http://paleoforum.ru/index.php/topic,168.msg9778.html#msg9778
[24] http://paleoforum.ru/index.php/topic,434.msg5613.html#msg5613
[25] http://paleoforum.ru/index.php/topic,168.msg40100.html#msg40100
[26] http://paleoforum.ru/index.php/topic,2231.msg48456.html#msg48456
[27] Цитата взята с сайта авторов исследований: http://sydney.edu.au/science/biology/shine/canetoad_research/scientific-publications-the-cane-toad-invasion.shtml
[28] Или даже соответствующие аллели могли появиться у этих жаб в Австралии de novo, но в тех пределах, которые характерны для этого вида. То есть, аналогично темным формам березовой пяденицы, которые независимо (но стабильно) появляются в Британии, континентальной Европе и Северной Америке (см. первую часть книги).
[29] Можно предположить и еще более сложный сценарий. Возможно, стремление путешествовать у этих жаб включается только тогда, когда им вообще есть, куда путешествовать. То есть, если жабы «чувствуют» (каким-то образом), что с одной стороны их «подпирает» множество особей собственного вида, а «другая сторона» полностью свободна от подобной конкуренции.
[30] Думаю, не надо объяснять, что стабилизирующий отбор – это нечто, противоположное эволюции (хотя и считается одной из форм естественного отбора). Потому что стабилизирующий отбор поддерживает признаки видов в рамках определенного, неизменного значения. А для примеров эволюции нужны установленные примеры движущего отбора (движущей формы естественного отбора), причем зашедшего так далеко, чтобы признак вышел за пределы значений, наблюдаемых для этого вида прежде.
[31] Вот здесь, например, перечислено 47 известных подвидов данной ящерицы: http://www.cyberlizard.plus.com/lacertids2s.htm
[32] В этих беседках происходит только ухаживание и спаривание, а гнездо самка вьет уже самостоятельно в другом месте.
[33] Возможно, самец считает, что брачная беседка еще не закончена - еще не такая украшенная, какой должна быть, вот и прогоняет самку? То есть, «эстетические чувства» самца (генетически запрограммированные) не позволяют ему спариваться с самкой до тех пор, пока соответствующая «брачная церемония», которая запрограммирована для этого вида, не будет отыграна по всем правилам, даже если самка «уже согласна».
[34] Ну а в самой википедии этот пример появился, скорее всего, благодаря заимствованию из книги Ричарда Докинза «Величайшее зрелище Земли: доказательства эволюции» (Dawkins, 2009). Что не удивительно. Во-первых, потому что книги Докинза давно стали для верующих дарвинистов священными. А во-вторых, потому что «установленных примеров эволюции» критически мало. Поэтому дарвинистам приходится всё время повторять одни и те же примеры, переписывая их друг у друга.
[35] Альтернативная ссылка на сайт с иллюстрацией: http://elementy.ru/lib/431425
[36] Рисунок взят из статьи русскоязычной википедии, посвященной «тлям Шапошникова»:
https://upload.wikimedia.org/wikipedia/commons/6/66/Dysaphis_anthrisci_majkopica.gif
[37] Для краткости цитирую просто из википедии, из статьи «Опыты Г. Шапошникова по искусственной эволюции»:
https://ru.wikipedia.org/wiki/Опыты_Г._Шапошникова_по_искусственной _эволюции
[38] По сути, данная синхронность является нарушением известного теста Лурии-Дельбрука. Этот тест как раз и демонстрирует случайный характер мутаций, именно тем, что соответствующие мутации в разных колониях бактерий возникают в очень разное время, настолько разное, что каких-либо закономерностей (и тем более, синхронности) там вообще не просматривается.
[39] Скорее всего, «эволюция гуппи на водопаде» тоже стала известна широкой массе верующих дарвинистов из их священной книги «Величайшее зрелище Земли: доказательства эволюции» (Dawkins, 2009).
[40] Конкретные значения (Reznick et al., 1997):
1) На участке реки Aripo спустя 11 лет (или примерно 18 поколений): самцы «над водопадом» - 58.2 дня против исходных 48.6 дней (до достижения половозрелости); самки «над водопадом» 93.5 дней против 85.6 дней «под водопадом». Масса самцов «над водопадом» 76.1 мг против 67.5 мг самцов «под водопадом». Масса самок 189.2 мг «над водопадом» против 162.3 мг «под водопадом».
2) На участке реки El Cedro, спустя 4 года (или 6.9 поколений): самцы «над водопадом» 72.7 дня против исходных 60.6 дней у самцов «под водопадом»; самки «над водопадом» 95.5 дней против 94.1 дня «под водопадом». Масса самцов «над водопадом» 62.4 мг против 56.0 мг самцов «под водопадом». Масса самок 118.9 мг «над водопадом» против 116.5 мг «под водопадом».
3) На участке реки El Cedro, спустя 7.5 лет (или 12.7 поколений): самцы «над водопадом» 52.5 дней против исходных 47.3 дней у самцов «под водопадом»; самки «над водопадом» 80.4 дней против 75.8 дней «под водопадом». Масса самцов «над водопадом» 74.4 мг против 71.5 мг самцов «под водопадом». Масса самок 152.1 мг «над водопадом» против 141.8 мг «под водопадом».
[41] Общую приспособленность конкретного организма можно оценить через его генетический вклад в следующее поколение, т.е. через общее число его потомков, в свою очередь, успешно достигших половозрелости.
[42] То есть, результаты этих исследований не подтверждены независимыми работами других авторов.
[43] См. рис. 21, нижний слева график изменения величины клюва – в 90-ых годах 20 века величина размера тела и клюва вьюрков практически полностью вернулась к своим исходным значениям (в 70-ых годах).
[44] Или, если говорить современным языком теории эволюции, «изменения частоты встречаемости аллелей в популяции», причем аллелей, либо уже имевшихся в этой популяции раньше, либо всё время возникающих у этого биологического вида (что тоже можно трактовать, как характерную часть внутривидовой изменчивости этого вида).
[45] Рисунок со свободным доступом, взят с сайта: https://en.wikipedia.org/wiki/Giraffe#/media/File:Giraffe_standing.jpg (автор и лицензия указаны на сайте).
[46] Рисунок со свободным доступом, взят с сайта: https://en.wikipedia.org/wiki/Giraffe#/media/File:Flickr_-_Rainbirder_-_Reticulated_Giraffe_drinking.jpg (автор и лицензия указаны на сайте).
[47] В сети Интернет легко найти соответствующие видеозаписи, где можно посмотреть, как самцы жирафов дерутся с помощью шей.
[48] Теоретические «ястребы» в модели Докинза никогда не уступают друг другу и дерутся до серьезных увечий или даже до смерти.
[49] Докинз рассматривает и другие, более гибкие (и еще более стабильные) стратегии поведения - «отпорщика» и «задиры». И тоже приходит к выводу, что в популяции должно постепенно устанавливаться определенное устойчивое соотношение разных типов поведения («отпорщиков» и «задир»). Такие эволюционно устойчивые равновесия (автоматически получаемые в ходе естественного отбора) Докинз, вслед за Мейнардом Смитом, называет эволюционно стабильными стратегиями.
[50] Удивительно, почему Докинз, рассуждая о борьбе между разными стратегиями (агрессивности) и придя к выводу, что в конечном итоге должно наступать равновесие разных стратегий… после этого не обращается за примерами к живой природе. Чтобы посмотреть, насколько его теоретические выводы соответствуют картине, которая наблюдается в реальности. А в реальности наблюдается разнообразие поведения разных видов животных по этому вопросу. Как ни странно, Докинз (порассуждав о том, к чему должна приводить эволюционная борьба разных стратегий в рамках предлагаемой им концепции «эгоистичного гена»)… затем ни капли не сокрушается по поводу того, что эти теоретические ожидания не подтверждаются картиной, наблюдаемой в природе в очень многих случаях. Например, в случае полового поведения таких биологических видов, где конфликты между самцами всегда имеют менее жесткий характер, чем это могло бы быть. То есть, где все самцы являются «голубями» (в терминологии Докинза).
[51] На всякий случай уточню, что Докинз в своих рассуждениях не делал акцент именно на борьбе самцов за самок. Он говорил, скорее, о борьбе за выживание в целом. В этом случае, его теоретические рассуждения можно и не считать ошибочными. Действительно, если «ястребы» серьезно «проредят» друг друга (в беспощадной борьбе) - тогда для «голубей» могут высвободиться какие-нибудь ресурсы. Например, пищевые. Но подобные рассуждения уже не проходят, как только мы начинаем говорить конкретно о борьбе самцов за самок (т.е. за доступ к размножению). Потому что самки представляют собой ресурс, который может быть полностью освоен даже редкими оставшимися самцами-«ястребами». То есть, сколько бы таких самцов ни осталось, они всё равно используют для размножения всех оставшихся самок. Если не сразу, то в эволюционной перспективе - у самцов должны усиливаться сооветствующие способности (сексуальный аппетит) по мере повышения числа доступных самок (естественный отбор будет поддерживать такие изменения в рамках самой же концепции «эгоистичного гена»). В этом случае для самцов-«голубей» места уже не остаётся (не остается ресурса размножения). Так что в отношении конкретно борьбы самцов за самок – теоретические рассуждения Докинза (о возможности существования стратегии «голубей» в живой природе) не подходят. Тем не менее, значительная часть наблюдаемых фактов «благородства животных» относится именно к области «рыцарских поединков», то есть, турнирных боев самцов за самку (а иногда даже просто «турнирных позирований»), где мы почему-то наблюдаем строгое соблюдение весьма гуманных правил самцами самых разных видов животных по отношению друг к другу.
[52] Стратегия «отпорщика» в терминах Докинза.
[53] Наивысшая скорость, которую продемонстрировал каждый конкретный гепард в исследовании (Wilson et al., 2013) за весь период наблюдений в природе составила: 25.9 м/c, 25.4 м/c, 22.0 м/c, 21.1 м/c, 20.1 м/c. То есть, 93 км/ч, 91 км/ч, 79 км/ч, 76 км/ч и 72 км/ч соответственно.
[54] Данные по величине ускорения и торможения гепардов приведены на всякий случай - в работе (Wilson et al., 2013) было установлено, что гепарды могут развивать очень впечатляющее ускорение и торможение – гораздо выше, чем у других измеренных «скоростных» млекопитающих. Однако столь впечатляющие результаты гепарды показывали тоже лишь в редких случаях. В подавляющем же большинстве случаев они демонстрировали гораздо более скромные показатели ускорения и торможения (см. рис. 25).
[55] Здесь необходимо иметь в виду, что по поводу гепардов мы сейчас рассуждали на основании только одного (последнего) исследования. Поэтому для окончательных выводов лучше подождать результатов аналогичных независимых исследований, выполненных другими авторами.
[56] Телепередача от 13.11.2012; дискуссия богословов и генетиков.
[57] По крайней мере, на восьми разных видах этого рода (Berlocher & Enquist, 1993).
[58] Не путать с кизилом.
[59] Т.е. насекомых, питающихся растениями.
[60] Cкорее всего, Марков прочитал еще статью (Feder, 1995).
[61] Кроме того, в эту группу входит еще несколько обособленных популяций мух с пока еще не до конца выясненным таксономическим положением, которых мы здесь упоминать не будем.
[62] И это, пожалуй, самые поразительные факты из всех озвученных. Как я уже говорил выше, установлено, что яблоневые мухи активно «воротят нос» от запаха боярышника, а боярышниковые мухи, наоборот, от запаха яблок. Хотя эти фрукты, оказывается, прекрасно подходят для развития личинок и тех и других. Вот такая вот она (на самом деле) «беспощадная борьба за выживание». Результат «жесточайшей конкуренции за все доступные ресурсы». Когда мухи в прямом смысле слова воротят нос от этих ресурсов. То есть, это такой (очередной) привет «рожденной на кончике пера» (кабинетной) «теории эволюции» от реальной живой природы.
[63] Абиогенез - это псевдонаучное фэнтези про «естественное самозарождение жизни в грязной луже», где под «грязной лужей» подразумевается то грязная лужа, то зарождение жизни в каких-нибудь геотермальных источниках, то образование «самовоспроизводящихся РНК» в порах каких-нибудь минералов, или какая-нибудь другая подобная дичь, совершенно невозможная с химической точки зрения.
[64] Так называемый «храповик Меллера». Гипотеза «храповика Меллера» первоначально была использована только для организмов, размножающихся бесполым путем. Но ничего не мешает расширить этот же механизм и на организмы с половым размножением. Потому что с одной стороны, организмы с половым размножением могут эффективно избавляться от груза вредных мутаций за счет рекомбинации, но с другой стороны, у организмов с половым размножением (диплоидных) только что возникшие вредные мутации тут же прикрываются здоровой копией этого же гена, что теоретически позволяет вредным мутациям избежать «очищения» отбором. И в результате в популяциях имеют возможность закрепиться не только слабо-вредные мутации, но даже просто вредные.
[65] То есть, сразу целыми системами ландшафтов.
[66] Все четыре изображения – фотографии, взятые с сайта https://commons.wikimedia.org/wiki/Main_Page со свободным доступом.
[67] Из всех наземных хищных млекопитающих, самые длинные клыки (относительно размеров черепа) сегодня, кажется, имеет дымчатый леопард. Но даже и его клыки серьезно отстают, например, от «сабель» травоядного мускусного оленя кабарги или водяного оленя.
[68] За исключением лишь моржей и некоторых видов особо зубастых рыб.
[69] Кажется, что если использовать «саблезубые» клыки, то вследствие их величины ими можно только эффективно бить (колоть), резко опуская голову (см. напр., Wroe et al., 2005). Потом (уже вонзив клыки), наверное, можно еще попытаться вспороть тело жертвы (хотя для слишком длинных зубов при этом более высок риск поломки). В то время как клыками нормальной величины можно спокойно (сильно) кусать, при этом крепко сжимая челюсти (современные хищные млекопитающие, рептилии и большое число хищных рыб). А так же «отрезать» куски (пираньи, акулы) или вспарывать тело жертвы (многие хищные животные с зубами нормальной величины). Авторы соответствующего исследования, в котором моделировалась сила укуса вымершей саблезубой кошки Smilodon fatalis, пришли к выводу, что укус этого крупного саблезубого хищника был примерно в три раза слабее, чем укус льва при сравнимой массе тела (McHenry et al., 2007). Кроме того, саблезубые кошки не могли рисковать, убивая своих жертв в движении – их зубы и даже череп рисковали от этого повредиться (McHenry et al., 2007). То есть, в этом отношении Smilodon fatalis резко отличалась от современных крупных кошачьих, которые без вреда для себя вцепляются зубами в движущиеся жертвы, масса которых может достигать нескольких центнеров.
[70] Рисунок сделан на основе изображения со свободным доступом с сайта https://commons.wikimedia.org/wiki/File:Beutelwolf_fg01.jpg (ссылки на автора и лицензию указаны на сайте).
[71] Следует обратить внимание, что морфология обсуждаемых черепов копирует друг друга всё-таки не абсолютно точно. Черепа отличаются. Например, у волка шесть передних резцов, а у тилацина – восемь. Тем не менее, общая форма (если можно так сказать) повторяется почти «один в один».
[72] Или даже (по другим оценкам) от 0 до 20 млн. лет.
[73] «Относительного» потому, что в более ранних (докембрийских) отложениях обнаружены более ранние фауны многоклеточных животных (так называемые эдиакарская и хайнаньская фауны). Однако они, по всей видимости, не были предковыми для той фауны, которая появилась в кембрии, и не показывают того разнообразия, которое характерно для кембрийской фауны. В связи с этим, российский палеонтолог Кирилл Еськов метко назвал эти ранние фауны «черновиками Господа Бога» («Знание - Сила», 2001, №6).
[74] А по сравнению с целым рядом современных групп членистоногих - даже лучше.
[75] Кстати, отсюда получается (если продолжать упорно рассуждать в рамках теории эволюции), что большая часть огромного разнообразия современных членистоногих, которые устроены проще, чем, например, высшие раки (Malacostraca), получилась не в ходе постепенного (эволюционного) усложнения организации, а наоборот, путем упрощения исходных раннекембрийских животных. Что звучит несколько забавно.
[76] Сингулярность – это воображаемое особое состояние некоего «ничего», из которого воображаемо возникла вся Наша Вселенная примерно 13 млрд. лет назад (с точки зрения современных физиков). Вот и новые типы животных, похоже, тоже «выпали из сингулярности» в раннем кембрии. То есть, из ниоткуда и сразу в готовом виде.
[77] И еще останки какой-то неопознанной мухи.
[78] Альтернативная ссылка на изображение: http://i.blogs.es/c3f65d/defensa_quimica/450_1000.jpg
[79] Глава «Палеонтологическая свинья возрастом 395.000.000 лет» в первой части книги.
[80] «Живые ископаемые» - это такие биологические таксоны, которые просто «забыли» про эволюцию. Они практически не изменились за десятки и даже за сотни миллионов лет, вплоть до наших дней.
[81] Предложенный ряд эволюции разных групп мечехвостов (друг в друга), наверное, выглядит достаточно убедительным. Особенно потому, что демонстрирует еще и определенную направленность изменений (эволюционные тенденции). Но ведь и построенный эволюционный ряд «выхода рыб на сушу» - тоже выглядел ничуть не менее убедительно. До тех пор, пока не был обрушен одним-единственным открытием (см. главу «Палеонтологическая свинья возрастом 395.000.000 лет» в первой части книги).
[82] «…крайняя редкость переходных форм в летописи ископаемых останков продолжает быть профессиональным секретом палеонтологии…» (Gould, 1977).
[83] Глава «Когда молекулярная биология не молчит, о чем она говорит?» в первой части книги.
[84] Глава «Когда молекулярная биология не молчит, о чем она говорит?» в первой части книги.
[85] Из резюме оригинального исследования (Gallant et al., 2014):
Наши результаты показывают, что, несмотря на миллионы лет эволюции и большую дивергенцию в морфологии клеток электрического органа, независимые линии выработали схожие факторы транскрипции и эмбриональные и клеточные пути в эволюции электрических органов.
[86] Глава «Когда молекулярная биология не молчит, о чем она говорит?» в первой части книги.
[87] Что, кстати, наводит на соответствующие аналогии с дарвинизмом в биологии.
[88] Биологический энциклопедический словарь / Гл. ред. М.С. Гиляров; редкол.: А.А. Баев, Г. Г. Винберг, Г.А. Заварзин и др. - М.: Сов. Энциклопедия. 1986. - 831 с.
[89] Да и Антарктида примерно 15 млн. лет назад, согласно существующим представлениям, начала резко «замораживаться». И по этой причине теоретический «мост», только что нарисованный нашим воображением (из Огненной Земли до Кергелена через Антарктиду) должен был прекратить свое существование.
[90] Результаты молекулярно-генетического метода, применяемого для определения эволюционного возраста биологических таксонов, сильно зависят от того, по каким конкретно биологическим объектам (или палеонтологическим находкам) калибровали те «молекулярные часы», которые затем использовались в той или иной работе для определения «возраста» биологического таксона. Стоит ли говорить о том, что такие исследования очень часто приводят к противоречивым результатам (нередко - крайне противоречивым) и к широким разногласиям в результатах разных (независимых) работ, посвященных изучению одного и того же вопроса. В некоторых случаях получается, что скорость работы «молекулярных часов» различается в разных биологических таксонах чуть ли не на два порядка. Это всё равно, что пытаться измерять время по часам, которые то практически стоят на месте, то «летят» со скоростью, в 100 раз превышающей нормальную.
[91] Хотя генетические различия (по идее) призваны отражать эволюционный возраст биологических групп и следовательно (?) общую «продвинутость» процессов видообразования в рамках конкретного таксона.
[92] Или несколько разных (основных) генетических линий примерно за 100 тысяч лет (согласно исследованию Verheyen et al., 2003), которые затем (в свою очередь) успели породить целые «пучки» разных биологических видов уже за гораздо меньшее время.
[93] На основе изображения с сайта википедии: https://ru.wikipedia.org/wiki/%D0%A9%D1%83%D0%BA%D0%B0#/media/File:Distribution_map_of_Esox_lucius.png (ссылки на автора и лицензию указаны на сайте).
[94] То есть, с примерно постоянной скоростью возникновения мутаций, и согласно правилам теории вероятности (например, мутация в одном и том же гене вряд ли случится больше, чем один раз подряд).
[95] Следует помнить, что «теория» полового отбора уже оскандалилась своей хронической невоспроизводимостью. То есть, имеет место низкая воспроизводимость результатов, опубликованных в научных исследованиях, посвященных изучению полового отбора у разных животных.
[96] Coregonus nasus. Рыба из рода сигов, семейства лососёвых. Ареал этого биологического вида тоже огромен - покрывает самые разные водоемы на большей части северной Канады и севера Евразии. Тем не менее, вся эта громадная территория населена одним видом (Coregonus nasus), а не тремястами.
[97] Gymnocephalus ambriaelacus, признанный отдельным видом, эндемик озера Аммерзе.
[98] Причем эти экологические формы то перестают скрещиваться друг с другом (и тогда верующие дарвинисты ликуют, заявляя о возникновении новых видов и «еще одном доказательстве эволюции»). То опять начинают скрещиваться. И тогда дарвинисты печалятся, объявляя, что «процесс эволюции сорван» (Behm et al., 2010). Понятно, что на самом деле (скорее всего) это никакая не эволюция, а просто существование одного и того же вида в двух разных экологических формах, что является весьма распространенным явлением для многих видов рыб. Это весьма интересная и поучительная тема, о которой я еще обязательно как-нибудь расскажу.
[99] Щитни - это жаброногие ракообразные.
[100] Некоторые генетические механизмы, которые могли бы такое сделать, в принципе, известны.
[101] Имеются в виду «хорошие», крупные виды (так называемые «виды в широком понимании»).
[102] Альтернативная ссылка: http://elementy.ru/images/news/neatral_forests-1.jpg
[103] Коэффициент положительной корреляции максимальной силы – единица. Это когда соответствующие ряды чисел полностью идентичны.
[104] Примерно от 0.5 до 0.8 между Америкой и Азией (т.е. от средней до высокой силы связи), и от 0.4 до 0.6 между Америкой и Африкой, и от 0.4 до 0.65 между Африкой и Азией (сила связи от умеренной до средней).
[105] Разделить формально, для собственного удобства, чтобы хоть как-то ориентироваться в этом (огромном) биологическом материале.
[106] До позднего плейстоцена.
[107] Хотя могут образовывать (в рамках вида) несколько разных цветовых вариаций.
[108] То есть, Карлсон существует, и он обязательно прилетит (когда-нибудь). Просто еще не прилетел.
[109] В рамках современных представлений о длительности геологических эпох, конечно.
[110] Период от позднего плиоцена до позднего плейстоцена. То есть, примерно от 2.5 млн. лет назад до 10 тыс. лет.
[111] Ископаемые виды насекомых, неотличимые от современных, к сегодняшнему дню обнаружены не только в балтийском янтаре, но и в янтаре из других географических регионов Земли. Возможно, в будущем я еще разовью эту тему – подробно разберу конкретный список таких биологических видов, которые вообще «забыли про эволюцию». Но не прямо сейчас.
[112] Определитель пресноводных беспозвоночных России и сопредельных территорий. Том 2. Ракообразные. Под редакцией С.Я. Цалолихина. Санкт-Петербург, 1995. 629 с.
[113] Тельсон – анальная лопасть у ракообразных.
[114] Если мне, конечно, не мещерится. Как я уже говорил, я не являюсь специалистом по щитням.
[115] И даже от другого современного рода щитней (Lepidurus) его тоже отличают лишь мелкие, биологические нейтральные признаки.
[116] Вплоть до изменения репродуктивного режима популяций, например, превращения бисексуальных популяций щитня летнего в популяции гермафродитов или в популяции с гермафродитами и самцами (Zierold et al., 2007).
[117] Рисунок со свободным доступом. Взят с сайта: https://en.wikipedia.org/wiki/Indian_peafowl#/media/File:Indian_Peafowl_on_Display.jpg (ссылки на автора и лицензию находятся на сайте).
[118] Подробно знакомиться с этими моделями будем уже в следующей книге.
[119] Разновидности теоретических моделей полового отбора: модель прямой выгоды, модель непрямой выгоды, модель «убегания» Фишера, модель гандикапа Захави (Jones & Ratterman, 2009).
[120] Имеются в виду: 1) версия «обоюдного» полового отбора, 2) версия о «случайном эволюционном заимствовании» (эстетического признака самца – самкой). Наверное, разобрать эти идеи всё-таки придётся, но уже в следующей книге.
[121] Рисунок со свободным доступом. Взят с сайта: https://en.wikipedia.org/wiki/Eudaemonia_troglophylla#/media/File:Exp%C3%A9dition_Sangha_2010_papillon.jpg (ссылки на автора и лицензию находятся на сайте).
[122] Рисунки со свободным доступом. Взяты с сайтов: https://en.wikipedia.org/wiki/Antherina_suraka#/media/File:Suraka_Silk_Moth_%28Antherina_suraka%29,_Vohimana_reserve,_Madagascar.jpg и https://nl.wikipedia.org/wiki/Maltagorea_fusicolor#/media/File:Butterfly_Madagascar_Antananarivo.JPG (ссылки на авторов и лицензию находятся на сайтах).
[123] Надеюсь, никто не будет отрицать, что внешность самца павлина может служить образцом эстетического дизайна высокого уровня. А ведь павлины – это представители куриных. Таким образом, в рамках современного дарвинизма получается, что курицы – это гении эстетического дизайна.
[124] Рисунки со свободным доступом. Взяты с сайтов: https://en.wikipedia.org/wiki/Harlequin_beetle#/media/File:Acrocinuslongimanus.JPG и https://en.wikipedia.org/wiki/Harlequin_beetle#/media/File:Acrocinus_longimanus_MHNT_femelle.jpg (ссылки на авторов и лицензию находятся на сайтах).
[125] Например, жуки-долгоносики из рода Mahakamia тоже имеют поразительно длинные передние лапы.
[126] Чтобы понять, насколько на самом деле «важна» стебельчатоглазость в поединках между самцами этих мух, достаточно просто понаблюдать за такими поединками. Мы увидим, что самцы стебельчатоглазых мух (во всяком случае, некоторых видов) с готовностью пинают друг друга передними ногами. Понятно, что преимущество здесь будет на стороне более крупных самцов (как обычно). А вот роль, собственно, стебельчатоглазости в этих баталиях остаётся не слишком очевидной (или даже совсем не очевидной).
[127] Цитаты приведены с сайта http://www.origins.org.ua/page.php?id_story=1056 Взято 4.06.2015
[128] Потому что вероятность даже двойной полезной мутации – уже слишком мала, чтобы её можно было принимать во внимание. Не говоря уже о тройной полезной мутации и выше.
[129] Ведь до тех пор, пока потенциально полезные аллели (такие варианты генов, которые могут принести пользу, но только в сочетании с другими аллелями) – являются еще редкими в популяции, они почти всегда будут находиться (в генотипе той или иной особи) только в единственной копии. То есть, нести этот аллель будет только одна хромосома (из двойного набора). Поэтому после расхождения гомологичных хромосом (несущих разные аллели) по разным половым клеткам – вероятность разбиения (исчезновения) удачного сочетания этих (пока еще редких) аллелей будет очень высока.
[130] Или даже девяностолетней эпопеи, если отсчитывать время не от знаменитого опыта Миллера-Юри, а от первых публикаций Опарина на эту тему.
[131] Период «полу-жизни» рибозы в воде с нейтральной реакцией (рH 7) при температуре 1000С – всего 73 минуты; при 250С – 300 дней; При 00С – 44 года (Larralde et al., 1995). В кислой и щелочной среде рибоза еще менее устойчива. «Время жизни» других сахаров находится примерно в таких же пределах (Larralde et al., 1995). В связи с этим авторы работы (Larralde et al., 1995) делают вывод, что рибоза и другие сахара не могли быть причастны к возникновению первичного генетического материала (нуклеотидов).
[132] Азотистые основания в горячей воде разрушаются очень быстро. При температуре 100°C аденин и гуанин имеют период «полураспада» около года, урацил приблизительно 12 лет, а цитозин всего лишь 19 дней. При 25°C скорость «полу-гидролиза» аденина и гуанина всё еще остаётся высокой (для геологических промежутков времени) - около 10.000 лет. Скорость же распада цитозина при 25°C продолжает оставаться практически мгновенной (по геологическим меркам) – 340 лет. И даже при 0°C, когда аденин и гуанин становятся уже достаточно устойчивыми, скорость «полу-гидролиза» цитозина всё еще остаётся высокой (для геологических промежутков времени) – около 17.000 лет (Levi & Miller, 1998). Исходя из этих фактов, авторы работы (Levi & Miller, 1998) приходят к выводу, что нестабильность азотистых оснований является очень серьезной проблемой для гипотезы естественного происхождения жизни.
[133] Каждый рибонуклеотид состоит из трех компонентов: азотистого основания, рибозы и фосфата.
[134] Особенно эта проблема актуальна при высоких температурах окружающей среды, что ставит жирный крест на всех предложенных гипотезах происхождения жизни в геотермальных источниках. А также на любых других гипотезах, в которых предполагаются высокие температуры среды. С другой стороны, и при низких температурах скорость распада сахаров и цитозина всё еще остаётся слишком большой. В то время как низкая температура замедляет вообще любые химические реакции (как реакции распада, так и реакции синтеза).
[135] Потому что нуклеотиды, необходимые для синтеза РНК, во-первых, слишком сложны, чтобы самостоятельно образовываться в неживой природе, а во-вторых, синтез этих нуклеотидов требует разных условий (синтез пуриновых и пиримидиновых нуклеотидов).
[136] Уридин под ультрафиолетовым излучением тоже неустойчив (см. напр., Гонтарева, 2003).
[137] Правда, после этого признания Сазерленд далее начинает теоретически рассуждать, каким образом эта проблема могла бы (теоретически) преодолеваться в условиях древней Земли (Sutherland, 2010).
[138] Как видим, даже в нашумевших опытах Сазерленда, на самом деле: 1) на разных стадиях эксперимента одни условия искусственно заменялись другими, 2) при этом промежуточные вещества очищались (см. выше).
[139] Кроме того, опыт Миллера-Юри имеет и другие методические недостатки. В этом опыте небольшой объем изолированной газовой смеси, циркулируя по кругу, в течение недели подвергался непрерывному воздействию электрических разрядов. Видимо, имитируя природную молнию, бьющую в одно и то же место тысячу раз подряд? В результате, и без того очень сомнительная (для земных условий) смесь газов (метан, аммиак, водород и пары воды) быстро превратилась в совсем уже нереалистичную искусственную смесь, с высоким содержанием циановодорода и формальдегида. И уже из этих веществ потом образовалось небольшое количество аминокислот (которые и наделали столько шума среди научной общественности). При этом основной аминокислотой, полученной в эксперименте, оказался глицин. То есть, самая простая из всех аминокислот (всего два атома углерода в составе молекулы), а также аланин (три атома углерода). Более сложные вещества образовывались в подобных экспериментах уже в следовых количествах. Но даже если взять именно глицин, то есть, именно ту аминокислоту, которой в эксперименте получилось больше всего, и вылить весь этот «продукт действия тысяч молний»… нет, не в древний океан, а всего лишь в емкость, содержащую один кубический метр воды, то в результате разбавления мы получим практически дистиллированную воду.
[140] Промежуточная очистка часто сопровождается еще и искусственным повышением концентрации необходимого вещества (до приемлемого уровня).
[141] Главной новостью в предложенном (коллективом Сазерленда) новом химическом пути, действительно, явилось именно преодоление печально известной «проблемы рибозы». Как известно, рибоза в неживой природе, во-первых, слишком нестабильна (см. выше). А во-вторых, она до этого упорно отказывалась соединяться с азотистыми основаниями (с образованием нужных нуклеозидов). Вот эту проблему и решил Сазерленд (просто обойдя её за счет других химических реакций). Однако предложенный им новый путь химических реакций до сих пор продолжает оставаться искусственным, поскольку еще не показано, каким образом этот путь (требующий разных условий) мог бы существовать в реальной природе. Здесь появляются уже новые трудности. О чем и говорит автор приведенной цитаты.
[142] Скорость транскрипции РНК у живых организмов составляет 40-50 пар нуклеотидов в секунду, или 140.000–180.000 нуклеотидов в час. Рибозим tC9Y присоединяет нуклеотиды со скоростью 4 нуклеотида в час. Это просто несоизмеримо с реально требуемыми скоростями синтеза. Рибозимы с такой скоростью работы скорее разрушатся (деградируют), чем доделают свою работу до конца. А чинить рибозимы от разрывов и ошибок некому. В живых клетках действуют специальные белковые комплексы для постоянного ремонта ДНК. А ведь ДНК намного устойчивей РНК.
[143] Сегодня в живой клетке действует сложнейший нанотехнологический белковый комплекс, который обеспечивает почти фантастическую точность копирования ДНК – вероятность ошибки (точечной замены) в живой клетке составляет примерно 10-9 (0.0000001%). И даже с такой точностью копирования вредные мутации (ошибки копирования) имеют тенденцию постепенно накапливаться в геномах. А теперь сравните эту точность с «точностью» работы обсуждаемого рибозима tC9Y, где вероятность точечных замен составляет почти 1%. И это помимо всех других типов ошибок (см. в тексте).
[144] Если для этого использовать высокую температуру (для «плавления» РНК), то эта же высокая температура довольно быстро «порвёт» все полезные молекулы.
[145] Известные научные эксперименты, которые проводил в 17 веке один из крупнейших ученых того времени Ян Баптист ван Гельмонт. В этих экспериментах ученый с помощью потной рубашки и горсти пшеницы успешно самозарождал мышей то в темном шкафу, то в горшках.
[146] И если «да», то приведите конкретные цитаты из соответствующих научных публикаций, пожалуйста.
[147] Например, в самой последней работе, коллектив Сазерленда сумел подобрать условия для синтеза ряда предшественников аминокислот, липидов и некоторых готовых простых аминокислот (Patel et al., 2015). Правда, здесь Сазерленд опять использовал очистку нужных веществ и изменение химических условий на разных стадиях процесса (например, искусственно удалялся кислород и т.д.). То есть, Сазерленд здесь снова задействовал методику, за которую сам же себя и критиковал в предыдущей статье (см. выше). Интересно, что в этой (новейшей) работе был сделан «упор» на циановодород, как на одно из ключевых исходных веществ. Но простейшие аминокислоты (глицин и аланин) из циановодорода получил еще Миллер в своем знаменитом эксперименте с электрическим разрядом в газовой смеси малого объема (см. выше). Так что, похоже, рекламируемые сегодня новейшие результаты (полученные коллективом Сазерленда в последней работе), на самом деле, рекламируются уже по новому кругу. Видимо, следуя принципу: всё новое - это хорошо забытое старое (С).
[148] В зависимости от природы того разума, который создал жизнь.
[149] За подробностями обращаться к физикам.
[150] Явление, когда в половых путях самки идет борьба сперматозоидов от двух или более разных самцов, недавно спарившихся с этой самкой (у тех биологических видов, где самки отличаются половой неразборчивостью). Получены определенные свидетельства, что в таких случаях сперматозоиды разных самцов могут тем или иным способом мешать друг другу оплодотворять яйцеклетки самок.
[151] В исследовании (Luck & Joly, 2005) было установлено, что уже осеменённые самки Drosophila bifurca отказывали самцам в 84% случаев, в то время как девственные самки продемонстрировали поведение отказа всего в 9% случаев. Причем авторы работы предполагают, что те осеменённые самки, которые всё-таки принимали повторные ухаживания других самцов (приблизительно 16% оставшихся случаев) – это были такие самки, первый половой партнер которых оказался с уже истощенным запасом спермы (недостаточным для полного оплодотворения яиц данной самки). Либо это были такие самки, которые по тем или иным причинам уже потеряли сперму предыдущего партнера. И эта потеря стимулировала данных самок на повторные спаривания (Luck & Joly, 2005). Таким образом получается, что самки этого вида мух вполне довольствуются спермой единственного самца (для одного акта оплодотворения яиц), и никакого «соревнования спермы» (от нескольких разных самцов) у этого вида не происходит. По крайней мере, в подавляющем большинстве случаев. Следовательно, использование гипотезы «спермовых войн» в качестве попытки объяснения потрясающего гигантизма сперматозоидов, не подходит для этого вида мух по факту.
[152] Ниже я сосредоточусь, в основном, на наиболее ярких фактах и самых последних открытиях. Для тех, кто дополнительно заинтересуется вопросом «ископаемой органики», рекомендую подробные русскоязычные обзоры на эту тему (Лунный А.Н., 2009а; 2009б; 2011).
[153] Остеоциты - клетки костной ткани.
[154] Белок - это сложное органическое соединение, длинная цепочка (нить) из аминокислот, соединенных между собой пептидными связями. Кроме того, разные части этой цепочки могут быть дополнительно соединены между собой химическими связями (между аминокислотными остатками удаленных друг от друга участков белковой цепи). В этом случае получаются разнообразные петли и спирали, в которые закручивается белковая нить. Нуклеиновые кислоты – это тоже сложные органические соединения, длинные цепочки (нити), но образованные не аминокислотами, а нуклеотидами.
[155] Скорость деградации ДНК в работе (Allentoft et al., 2012), кажется, получилась ниже, чем в работе (Nielsen-Marsh, 2002). В этих двух работах использовались разные методики. Оценка скорости деградации ДНК в работе (Nielsen-Marsh, 2002) основывалась на соответствующих химических экспериментах. А в исследовании (Allentoft et al., 2012) оценка проводилась путем сравнительного анализа сохранности ДНК в ископаемых костях, возраст которых, в свою очередь, измерялся радиоуглеродным методом. Однако в любом случае (по данным обеих работ), ДНК не может сохраняться многие миллионы лет (при сколько-нибудь положительных температурах).
[156] На определенной глубине, в так называемом нейтральном слое, температура земли перестает колебаться в зависимости от времен года и становится постоянно равной среднегодовой температуре этой местности.
[157] В любом случае, климат там был никак не менее чем субтропический, потому что там не могло быть холоднее, чем далеко за полярным кругом в то же самое время.
[158] Поскольку период полураспада изотопа углерода 14С относительно невелик (около 5700 лет), измерять с помощью радиоуглеродного метода можно возраст только таких объектов, которые образовались сравнительно недавно (примерно до 60 тысяч лет). Возможно, авторы озвученной работы применили радиоуглеродный анализ к «многомиллионолетней» кости мозазавра для того, чтобы показать, что изотопа углерода 14С там уже практически не осталось, и следовательно, органика в костях мозазавра действительно древняя. Но неожиданно получилось то, что получилось - радиоуглерода оказалось достаточно много для того, чтобы дать возраст 24600 лет.
[159] http://www.sciencedaily.com/releases/2013/04/130410131216.htm
[160] Белемниты - вымершая группа головоногих моллюсков.
[161] Столь большие оценки, как правило, были сделаны на основании косвенных данных. С помощью примерно таких рассуждений - раз в этом районе когда-то росло дерево, то значит, это было еще до такого-то периода оледенения (или, например, до соответствующего изменения береговой линии моря). А поскольку считается, что (такой-то) ледниковый период (или такое-то изменение береговой линии) произошли сто тысяч лет назад, то значит, этому дереву должно быть более ста тысяч лет. Интересно, что радиоуглеродный анализ некоторых из таких «ископаемых пеньков», тем не менее, показал всего примерно 12.000 лет (Stahle et al., 2012). В то же время, радиоуглеродный анализ других ископаемых стволов и пней показывал возраст более 50.000 лет (Stahle et al., 2012).
[162] Факты обнаружения относительно простых органических веществ в ископаемых останках с предполагаемым возрастом десятки миллионов лет являются менее показательными, чем сохранение крупных фрагментов белковых молекул или ДНК. Потому что многие простые органические вещества способны сохраняться гораздо дольше белков и ДНК. Тем не менее, факты обнаружения простой органики, во-первых, дополняют общую картину, а во-вторых, некоторые из таких находок поражают уже сами по себе, из-за громадного предполагаемого возраста этих находок (см. далее по тексту).
[163] Эдмонтозавры – одни из крупнейших представителей семейства утконосых динозавров (Hadrosauridae).
[164] Цитата из резюме работы (Moczydlowska et al., 2014): «The biogeochemistry of the organic matter that comprised the tube, which was inferred from its properties, composition, and microstructure, is consistent with chitin and proteins as in siboglinids».
[165] Фрамбоиды пирита – минеральные образования серного колчедана.
[166] «Потому что этого не может быть никогда» (С)
[167] Эти кристаллы соли в пермский период, по-видимому, являлись частью «рассола» (очень соленого водоема), где обитали древние галобактерии.
[168] Причем выделили, похоже, вместе с самими бактериями.
[169] Железо входит в состав гемоглобина крови.
[170] Нагревание повышает внутреннюю энергию (движение молекул) и соответственно повышает скорость химических реакций. Благодаря чему можно в реальном времени приблизительно оценить скорость распада белковых молекул при обычных температурах (зная соответствующие зависимости).
[171] Помимо «железной теории», иногда озвучивается столь же произвольная (непроверенная) гипотеза о сохранении белков в костях динозавров за счет образования устойчивых комплексов с биоапатитом оригинальной кости (с образованием неких «внутренних кристаллов»). Впрочем, частично эта гипотеза всё же была проверена – в работе (Collins et al., 2000) сообщается, что сохранность остеокальцина (самого прочного белка из всех здесь озвученных) находится в обратной зависимости от степени кристаллизации апатита кости (зависимость выведена путем сравнения степени сохранности остеокальцина в костях разного возраста). То есть, чем сильнее кристаллизуется биоапатит костей, тем хуже сохранность остеокальцина. Получается, что образование комплексов с кристаллами апатита, не повышает, а наоборот, понижает устойчивость белка (во всяком случае, в приведенном примере).
[172] Возможно, цель состояла не только в создании разных живых существ, но еще и в обеспечении их существования на Земле в течение определенного периода времени. Может быть, такие периоды существования предусматривались для целых специфических биомов. Например, какие-нибудь «моря силура». Или «динозавровые ландшафты» и т.п.
[173] Такое поведение самцов известно как для других (близких) видов сверчков, так и для «поющих» (немутантных) самцов этого же вида (Teleogryllus oceanicus). То есть, нормальные (немутантные) сверчки этого вида тоже могут привлекаться песней других самцов (например, если их собственное пение в течение некоторого времени не имело успеха в деле привлечения самок). Однако нормальные (поющие) сверчки этого вида приближаются к другим самцам в меньшей степени, чем «онемевшие» самцы, которые, как оказалось, сближаются с поющими самцами на расстояние, примерно в 1.5 раза меньшее (Zuk et al., 2006).
[174] На самом деле, называть только что озвученные факты примером эволюции - вряд ли корректно. В лучшем случае, здесь можно говорить о работе естественного отбора. Однако никакой эволюции здесь нет (по факту). Потому что частота встречаемости этого признака, хотя и явно повышена в некоторых популяциях людей, живущих в регионах с риском малярии, тем не менее, во-первых, не слишком высока (до 15%), а во-вторых, она не поднимается дальше. Наверное, не поднимается именно потому, что если эта частота станет выше, то аллели серповидноклеточной анемии начнут слишком часто находить друг друга, то есть, слишком много людей получат серповидноклеточную анемию в гомозиготной форме. А в этом состоянии данная мутация становится настолько вредной, что устойчивость к малярии в качестве полезного «бонуса» уже, наверное, не окупает негативных симптомов болезни. Вот поэтому частота встречаемости аллеля серповидноклеточной анемии дальше не повышается. А раз дальнейшего увеличения встречаемости этого аллеля не происходит (по факту), то значит, здесь нет и никакого примера эволюции.
[175] Правда, есть еще вариант, что «эволюционно онемевшие сверчки» - это, на самом деле, просто еще один пример пресловутого телекинеза в биологии (см. выше). Но, честно говоря, не хотелось бы так думать.
[176] До 6% процентов (Beckers & Wagner, 2012).
[177] Эволюция микробных сообществ. Доклад, прочитанный на теоретическом семинаре геологов и биологов «Происхождение живых систем». 15-20 августа 2003 г., Горный Алтай, стационар «Денисова Пещера». Электронная публикация. http://www.evolbiol.ru/zavarzindok.htm
[178] Интересно, что авторы работы (Schopf et al., 2015) каким-то образом умудрились сделать вывод, что абсолютный эволюционный застой на протяжении почти 2 млрд. лет, который они сами же и обнаружили в сообществах серобактерий, «оказывается»… подтверждает теорию Дарвина! (в качестве какой-то там нуль-гипотезы, которая была сразу же придумана для этого случая). То есть, если бы биологический таксон со временем изменялся, то это бы подтверждало теорию Дарвина. А если биологический таксон никак не изменяется на протяжении вот уже 2.000.000.000 лет… то это тоже подтверждает теорию Дарвина! Что и говорить, мысленным выкрутасам верующих дарвинистов просто нет предела.
[179] Эукариоты – организмы, в клетках которых имеется ядро. Эукариоты обеспечивают подавляющую часть разнообразия форм живых организмов Земли.
[180] Впрочем, в ходе этого исследования была обнаружена одна тонкость, которая может указывать на то, что механизмы образования генетических связей здесь не такие простые, как кажется. Вот этот ньюанс (Марков, 2009б):
…Бактерии, которых в течение 500 поколений выращивали в лактозной среде без добавления мальтозы, полностью «разучились» включать свои мальтозные гены в ответ на присутствие лактозы. Интересно, что при этом они вовсе не утратили способность переваривать мальтозу. Мальтозные гены сохранились в целости, и по-прежнему включались в ответ на появление в среде самой мальтозы. Исчезло только опережающее реагирование.
Не очень понятно, почему в результате случайных мутаций разрушились только генетические связи, отвечающие за обсуждаемый «условный рефлекс», но не сами мальтозные гены? Ведь они же тоже не использовались в течение тех же 500 поколений. Это случайность? Или, может быть, сами мутации здесь не такие уж случайные? Возможно, они происходят лишь на тех участках ДНК, которые ответственны именно за «условные рефлексы», то есть, за взаимодействие с изменяющейся средой. А какие-то важные участки ДНК (определенные гены) мутациями обходятся стороной (или почти стороной)?
[181] Таблица взята из статьи «Полный перебор» русскоязычной википедии. Представлено оценочное время полного перебора паролей в зависимости от их длины. Предполагается, что в пароле могут использоваться 36 различных символов (латинские буквы одного регистра + цифры), а скорость перебора составляет 100 000 паролей в секунду (класс атаки В, типичный для восстановления пароля из кэша Windows (.PWL файлов) на Pentium 100).
[182] В работе (Maisnier-Patin et al., 2005) оценивалась частота разных мутаций, которые сами по себе имели очень разную вероятность, вплоть до разницы в несколько порядков. Наименее частая мутация происходила с вероятностью 10-9, в то время как наиболее частая – с вероятностью 10-7 при максимально подавленной работе гена-мутатора, и ~ 10-5 при максимально активной работе гена-мутатора.
[183] Для простоты я не рассматриваю еще и четыре возможных варианта замены (четырьмя разными нуклеотидами), а считаю, что уже сама замена, как таковая, будет решать проблему.
[184] В последнем случае бактерии были не разморожены, а просто извлечены из подземного резервуара, который согласно имеющимся представлениям стал полностью изолированным уже 4 млн. лет назад.
Дата: 2019-02-02, просмотров: 507.