Озвученные выше факты, проблемные для современного дарвинизма, можно переформулировать и другим, более общим способом. Можно сказать, что живая природа в большом числе случаев демонстрирует нам явную дискретность («раздельность») биологических видов. В то время как в рамках современного дарвинизма, такое явление наблюдаться, в общем-то, не должно. По крайней мере, дискретность биологических видов должна наблюдаться существенно реже, чем плавные переходы между видами.
Потому что с точки зрения современной теории эволюции, само понятие биологический вид - достаточно иллюзорно. Это скорее, нечто кажущееся нам реальным на определенном промежутке времени. То есть, существующие сегодня «биологические виды» - это как бы «моментальный снимок» вечно идущего эволюционного процесса. Который мы как бы «поймали» своей «фотографией» в определенный момент. И наблюдаем мы этот эволюционный процесс в виде некоторых «групп организмов», обладающих определенной общностью, и называем эти группы - биологическими видами. Но стоит нам немного передвинуть «планку» времени, как мы увидим уже другие «группы организмов», и соответственно, выделим уже другие биологические виды.
С позиций современной теории эволюции, эволюционируют не виды, а отдельные (изолированные) популяции этих видов. Которые постепенно «уходят» друг от друга за счет совершенно естественных и автоматических процессов: 1) случайных мутаций, 2) естественного отбора, и 3) дрейфа генов. Эти автоматические процессы постепенно приводят к тому, что между разными изолированными популяциями постепенно накапливаются различия (морфологические, экологические, поведенческие, генетические). И в конце концов, уровень этих накопленных различий доходит до того, что соответствующий специалист по этому таксону опишет эти (столь далеко разошедшиеся) популяции – как разные биологические виды. То есть, биологический вид с точки зрения современной теории эволюции – это просто одна из популяций какого-то другого биологического вида (например, предыдущего, или соседнего), которая уже «ушла» (в деле накопления всяких различий) дальше, чем другие популяции этого же (предыдущего) вида.
Но в рамках подобного взгляда на эволюцию, мы должны наблюдать в природе не столько разные биологические виды, сколько непрерывную «кашу» из множества изолированных популяций, всё больше отличающихся между собой (или от какого-то принятого «эталона»), вплоть до различий такого ранга, который специалисты уже будут склоняться классифицировать, как другой биологический вид. И такая картина - плавных, непрерывных, постепенных переходов между разными биологическими видами, должна наблюдаться в подавляющем большинстве случаев. Такая картина сама по себе была бы мощнейшим свидетельством в пользу появления биологических видов именно по сценарию постепенной эволюции разных популяций.
Напротив, если бы во всех наблюдаемых случаях мы видели в природе совершенно явную дискретность биологических видов, где в каждом случае было бы ясно, что вот это - один биологический вид, а вот это - уже другой… такая картина явно опровергала бы тот сценарий эволюции, который предлагает современный дарвинизм.
Какую же из этих (возможных) картин мы наблюдаем в реальной живой природе?
К сожалению, реальность в данном случае (как всегда) сложнее простых теоретических схем. То, что мы видим в живой природе, наверное, можно охарактеризовать фразой «серединка на половинку». То есть, в живой природе мы видим как чрезвычайно многочисленные примеры явной дискретности видов, так и наверное не менее многочисленные примеры (казалось бы) «плавных переходов» между биологическими видами. Таким образом, в данной области, и сторонники, и противники дарвиновской эволюции могут «понадёргать» из живой природы огромное число примеров, как подтверждающих современный дарвинизм (с его сценарием постепенной эволюции разных популяций), так и примеры, которые ставят этот сценарий под большое сомнение. Или даже полностью опровергают его (см. например, ниже про бесполые и партеногенетические виды).
Чуть ниже я приведу целый ряд именно таких фактов - одни из которых будут просто (немного) больше свидетельствовать в пользу дискретности биологических видов, чем в пользу «плавных переходов между видами». Другие же примеры уже будут четко показывать, что биологические виды – это именно реальные, дискретные биологические объекты (а не иллюзорные сущности, выделяемые почти формально). Причем в некоторых случаях эти «сущности» оказываются настолько устойчивыми, что возникает острое желание считать биологические виды – неизменными.
С другой стороны, сторонники дарвиновской теории тоже могут много чего сказать в свою пользу (в этой области).
Во-первых, они могут отметить то обстоятельство, что реально наблюдаемая дискретность (многих) биологических видов – совсем не обязательно опровергает дарвиновский сценарий эволюции. Хотя бы потому, что в природе на разные популяции действует много разных внешних факторов. В том числе таких, которые могут легко нарушить (исходную?) «плавность переходов» между совокупностью разных эволюционирующих популяций. Например, сильно разыгравшиеся (в особо засушливый год) массовые лесные пожары могут уничтожить «серединку» в каком-нибудь плавном ряду (гипотетически эволюционирующих) популяций тех или иных растений. И в результате мы теперь, вместо плавного ряда этих популяций (непрерывно «переходящих» друг в друга) будем наблюдать только два «крайних» (сохранившихся) «куска» этого (прежде непрерывного) ряда, которые уже достаточно резко отличаются друг от друга, чтобы мы четко видели различия между ними, и назвали их разными биологическими видами.
Или другой сценарий – представители какой-нибудь отдельной популяции попадают в новый регион, свободный для расселения. И соответственно, эта популяция быстро расселяется по этому региону. Гораздо быстрее, чем разные представители этой (исходной) популяции начнут эволюционировать на этой территории (постепенно накапливая различия между собой). В результате мы станем наблюдать на всей этой территории – совершенно четкий (дискретный) биологический вид, хорошо отличающийся от других биологических видов на этой территории.
Можно придумать и другие похожие сценарии, каким образом из непрерывно эволюционирующих совокупностей популяций могут выделяться достаточно четкие «куски», чтобы нам это потом казалось именно дискретным биологическим видом. Таким образом, просто факт, что многие биологические виды действительно демонстрируют именно дискретность – еще не опровергает дарвиновский сценарий эволюции.
Кроме того, сторонники дарвиновской теории могут привести в свою пользу не менее многочисленные примеры, когда систематики (специалисты по самым разным биологическим таксонам) бесконечно путаются, сомневаются и вступают в оживленные дискуссии друг с другом - считать ли те или иные биологические совокупности – разными видами, или не считать? Можно ли ограничиться, например, уровнем подвида (в рамках единого биологического вида) или уже нельзя? Иногда такие дискуссии могут тянуться бесконечно. Дело дошло до того, что среди биологов довольно популярным стало такое шутливое определение биологического вида:
Биологический вид – это то, что считает видом специалист по данному таксону.
Причем в этой шутке, на самом деле, присутствует только малая доля шутки. Потому что выделение (или не выделение) биологических видов до сих пор так и происходит – преимущественно, методом экспертной оценки. Причем выводы, сделанные разными специалистами, нередко не совпадают друг с другом (что и порождает соответствующие дискуссии). Такая картина, уже сама по себе, красноречиво намекает на «неуловимость» (иллюзорность) понятия биологический вид. А эта «неуловимость», в свою очередь, свидетельствует в пользу именно эволюционного подхода к биологическому виду. Где разные биологические виды считаются чуть ли не формальными сущностями, выделяемыми лишь в качестве «моментальных снимков» - для того чтобы хоть как-то «запечатлеть» (формализовать) процесс постепенного накопления изменений в разных популяциях.
И теперь, если к этой картине добавить еще и тот факт, что многие биологические виды являются настолько близкими друг к другу, что разницу между ними способны увидеть только специалисты (причем одни специалисты могут считать, что этой разницы достаточно для разделения на разные виды, а другие – что недостаточно). И если сюда же добавить тот факт, что многие биологические виды, в свою очередь, сами изменчивы, и могут разделяться на разное число подвидов (в некоторых случаях число подвидов может достигать многих десятков). А в рамках подвидов, в свою очередь, могут «проглядывать» еще и разные варианты и расы… То вся эта картина в целом, кажется, как раз и выглядит именно так, будто мы пытаемся разделить на разные биологические группы - именно непрерывный процесс постепенного изменения (эволюции) всех живых существ Земли.[105] Тем более что споры систематиков отнюдь не ограничиваются только рангом биологических видов. Совершенно аналогичные дискуссии идут между систематиками и по поводу того, относить ли какой-нибудь биологический вид – к тому или иному биологическому роду, или же следует относить его к другому биологическому роду (или вообще выделить этот вид в рамках нового биологического рода). И тому подобное.
Все такие факты (еще раз) говорят в пользу именно «непрерывности» биологических видов (так же как и более высоких таксономических рангов).
Наконец, в некоторых установленных случаях природа, казалось бы, предоставляет нам совершенно четкие иллюстрации именно непрерывной эволюции разных популяций, где на разных «концах» (непрерывного ряда популяций) дело доходит уже до ранга разных биологических видов. Это примеры так называемых «кольцевых ареалов».
«Кольцевые ареалы» – это когда имеется ясный ряд из географически сменяющих друг друга популяций (того или иного биологического вида) с постепенно усиливающимися различиями, которые идут по некоему «кольцу» (либо вокруг какого-нибудь географического препятствия, либо вообще вокруг Земли). И в конечной точке, где популяции вновь встречаются (допустим, после длительного процесса постепенного расширения ареала этого вида) - эти (конечные) популяции уже настолько отличаются друг от друга, что ведут себя уже как разные биологические виды. Например, представители этих (крайних) популяций уже не скрещиваются друг с другом. Хотя все соседние популяции (на всех других участках этого «кольца») могут свободно скрещиваться между собой. Таким образом, «кольцевые ареалы» - это как раз тот случай, который свидетельствует в пользу образования видов по сценарию, предлагаемому современной теорией эволюции - наилучшим образом.
Что могут возразить на всё это сторонники дискретности биологических видов? Допустим, сторонники раздельного создания биологических видов?
Интересно, но им тоже будет, что сказать.
Во-первых, какую-то часть бесконечных дискуссий систематиков (по поводу ранга тех или иных биологических групп) – можно объяснить просто тем, что мы еще недостаточно хорошо знакомы с теми биологическими объектами, которые пытаемся классифицировать. Действительно, если мы сами, попадая в некоторые страны, первое время путаемся в лицах окружающих людей (лица людей этой страны первое время кажутся нам одинаковыми)… то что уж тогда говорить о попытках систематиков разобраться в разнообразии каких-нибудь круглых червей? Или диатомовых водорослей? Понятно, что в этом случае существенная часть путаницы, и соответственно, дискуссий специалистов, которые пытаются разобраться в тех или иных биологических таксонах – может быть порождена просто недостаточным умением (пока) ориентироваться в этих таксонах.
Дело осложняется тем, что нередко, после того, как тот или иной специалист научится видеть какие-то различия между разными совокупностями одного и того же биологического вида – у этого специалиста возникает чувство, что увиденные им различия очень важны, и на основании этих различий надо непременно выделить наблюдаемые группы в разные биологические виды. Это приводит к тенденции к постепенному дроблению биологических видов (на виды всё меньшего «объема»), по мере погружения в изучение той или иной группы живых существ. И хорошо, если в этом случае вообще найдется какой-нибудь другой специалист по этому же таксону (специалистов по многим биологическим таксонам «в природе» вообще единицы), который попытается «отрезвить» первого специалиста. Например, напомнить ему, что таким образом можно и людей разделить сразу на несколько разных видов. Или даже соседей по лестничной площадке выделить уже в другие «виды». Потому что какие-то различия мы в любом случае обязательно найдем. Причем здесь я почти не шучу – лично был знаком с некоторыми систематиками (ботаниками), которые серьезно считали, что современного человека следовало бы разделить на несколько разных видов. Таким образом, путаница и дискуссии специалистов могут быть порождены еще и такими (во многом, субъективными) причинами.
Кроме того, в последнее время при выделении видов используются еще и молекулярно-генетические критерии. То есть, если теперь еще и генетики начинают «видеть резкие отличия» в генах разных популяций какого-нибудь вида, то только на этом основании эти популяции сегодня могут быть разделены на разные виды. Даже в том случае, если эти популяции мало чем различаются друг от друга во всех остальных отношениях. В результате, новые «скрытые виды» («cryptic species» или «sibling species») сегодня стали плодиться буквально «как грибы». Однако возникает вопрос, стоит ли считать такие «виды» - настоящими («хорошими») биологическими видами? Или не стоит.
Кроме того, некоторые, казалось бы, хорошие примеры «непрерывности видов», на самом деле, серьезно свидетельствуют о том, что механизм образования новых видов, возможно, вообще не дарвиновский. Может быть, этот механизм просто заранее запрограммирован в тех или иных биологических таксонах. Здесь хорошим примером являются те самые биологические таксоны, которые в настоящее время еще как бы «рассыпаются» на разные виды (см. выше). Именно такие таксоны создают впечатление иллюзорности границ вида. То есть, когда в никарагуанских озерах плавает некий (даже не знаю как назвать) комплекс (будущих биологических видов) под названием Amphilophus cf. сitrinellus, который настолько переполнен этими «будущими видами», что норовит тут же «рассыпаться» на них в любом мало-мальски захудалом озере… то если не вдумываться, такой пример кажется нам подтверждением наших идей о видообразовании в рамках современного дарвинизма. А вот если изучить этот пример внимательно (см. выше), то мы увидим здесь явные намеки на появление биологических видов по механизму… заранее запрограммированного процесса.
И наконец, по поводу знаменитых «кольцевых ареалов», как это ни странно, но можно сказать больше всего.
Во-первых (и это самое главное) конкретные примеры таких «кольцевых ареалов» в природе являются крайней редкостью. Если мне не изменяет память, к сегодняшнему дню на всю живую природу (!) найдено аж целых пять штук эти самых «кольцевых ареалов». Что само по себе поразительно. Потому что если бы теория эволюции была права в том, что вся живая природа (которую мы наблюдаем сегодня) получилась именно путем постепенного накопления различий между разными популяциями… То наверное, мы должны были бы ожидать во много раз большей частоты встречаемости «кольцевых ареалов» в природе, чем та, которую мы реально наблюдаем (пять штук на всю живую природу к сегодняшнему дню).
Во-вторых, классические (известные из учебников) примеры «кольцевых ареалов»… уже подверглись пересмотру и критике в более поздних публикациях. Например, в работе (Liebers et al., 2004) критикуется самый знаменитый пример - «кольцевой ареал» серебристой чайки. А в отношении (тоже уже давно ставшего известным) «кольцевого ареала» большой синицы - интересная информация содержится в работах (Packert et al., 2005; Федоров и др., 2006). Например, в последней работе была показана свободная гибридизация тех самых «концевых популяций», которые, как считалось раньше, не скрещиваются друг с другом и таким образом ведут себя как разные виды (в соответствии с концепцией «кольцевого ареала»). А оказалось, вполне себе скрещиваются.
Таким образом, старые примеры «кольцевых ареалов» (из установленных пяти штук) уже, похоже, раскритикованы. А остальные, может быть, просто дожидаются своей очереди.
И тем не менее (несмотря на всё сказанное) вряд ли можно исчерпывающе объяснить (только озвученными причинами) огромное число примеров, когда не совсем ясно (или даже совсем неясно) - где следует «делать талию» (С). То есть, выделять или не выделять разные биологические виды. Таких примеров (еще раз) огромное количество. Поэтому на сегодняшний день, в споре «дискретность vs непрерывность» биологических видов – вряд ли можно уверенно принять чью-нибудь сторону.
Однако мы в этой книге ведь критикуем современный дарвинизм. Значит, должны приводить такие факты, которые не вписываются в сценарий дарвиновской эволюции. Поэтому я сейчас приведу целый ряд конкретных примеров, указывающих именно на дискретность биологических видов. Некоторые из таких примеров вообще из ряда вон. То есть, вообще опровергают дарвиновский сценарий эволюции (по факту). Тем не менее, следует помнить, что и фактов «непрерывности биологических видов» (косвенно свидетельствующих в пользу возможности их свободного превращения друг в друга) при желании, тоже можно насобирать в живой природе сколько душе угодно.
Итак, в значительном числе случаев биологические виды хорошо сохраняют свои видовые признаки на всём ареале собственного распространения. Даже если этот ареал огромен, и включает тысячи идеально изолированных популяций. Именно в таких случаях (когда имеются огромные ареалы) факты наблюдаемой дискретности биологических видов (на таких громадных площадях) начинают серьезно «подтачивать» уверенность в дарвиновских сценариях видообразования.
Например, мы наблюдаем, что всё та же (уже много раз помянутая нами) обыкновенная щука остаётся обыкновенной щукой на всём протяжении своего ареала обитания. Невзирая на то, что этот ареал просто колоссален по площади (и очень изменчив по экологическим условиям). Никакого континуума популяций щук, в котором различия постепенно достигают ранга другого вида – мы почему-то не наблюдаем. А наблюдаем мы именно обыкновенную щуку – по всему её гигантскому ареалу. И еще, конечно, наблюдаем некоторые колебания внутривидовой изменчивости между разными популяциями этих щук, которые, тем не менее, не выходят за рамки внутривидовой изменчивости (иначе бы систематики разделили щуку на разные виды). Ну а весь род щук (Esox), в свою очередь, насчитывает менее десятка других видов, причем ареалы этих видов во много раз скромнее ареала распространения щуки обыкновенной.
То есть, в данном случае мы наблюдаем в природе картину, по сути, противоположную той, которую предсказывает современная теория эволюции.
Другой пример. Всем известный род Panthera включает в Старом Свете такие хорошо отличимые друг от друга виды, как тигр, леопард и лев. Предполагается, что все три перечисленных вида произошли от некоего общего предка, изолированные популяции которого постепенно всё накапливали и накапливали различия между собой. И в конце концов эти различия накопились до такой степени, что в некоторых древних популяциях «выкристаллизовались» три разных современных вида: тигр, леопард, лев.
Но если описанный сценарий верен, то мы должны и сейчас наблюдать аналогичные процессы, где разные изолированные популяции львов, леопардов и тигров образуют некий континуум (непрерывный переход), в котором различия между популяциями доходят до таких величин, когда уже не очень понятно, кто перед нами – тигр, лев, леопард или вообще какая-нибудь «неведомая зверушка».
Кто-нибудь наблюдает подобную картину в рамках этого рода? Нет. Несмотря на обширные ареалы, хорошо изолированные популяции и внутривидовую изменчивость, которая воплощается в известных географических вариациях у всех обсуждамых видов, мы наблюдаем в природе картину противоположную – все три озвученных вида крупных кошек прекрасно различаются между собой на всем протяжении их ареалов. То есть, устойчиво сохраняют свои видовые признаки без каких-либо наблюдаемых «плавных переходов». А ведь ареал, например, льва (Panthera leo) был в недавнем прошлом[106] просто огромен – он охватывал почти всё северное полушарие, проходя по Евразии и Северной Америке, и еще захватывал почти всю Африку. И на всём этом колоссальном пространстве лев разделялся только лишь на подвиды (по мнению большинства специалистов). Ареал обитания леопарда тоже до недавнего времени был громаден - хотя и не достигал Нового Света (в отличие от ареала льва), но занимал Африку и тянулся через всю Азию вплоть до Амура на севере и островов Индонезии на юге. В прошлом (до позднего плейстоцена) леопард был широко распространен еще и по всей Европе.
Интересно, что в Центральной и Южной Америке живет всем известный ягуар, то есть, еще один представитель рода Panthera, который, вроде бы, морфологически близок к леопарду, но при этом отличается от льва. Хотя как раз львы в Новом Свете обитали, а вот леопарды – нет. В связи с этим считается, что предки ягуаров (уже будучи похожими именно на ягуаров), постепенно «притопали» из Азии в Центральную и Южную Америку, совершив миграцию весьма впечатляющей протяженности - через Чукотку и Аляску, которые тогда (предполагаемо) были связаны друг с другом, образуя так называемую Берингию. Через эту же Берингию в Северную Америку (тоже предполагаемо) «притопал» еще и лев, освоившись там, и образовав американский подвид.
Вообще следует отметить, что эта удивительная страна Берингия была в своё время очень оживленным местом. Через неё шёл постоянный поток самых разных живых существ, мигрирующих туда и обратно (из Евразии в Северную Америку и наоборот). Львы, медведи, волки, лисы, лоси, лошади, верблюды, щуки, десятки видов растений и многие-многие другие - все они посчитали своим долгом мигрировать на расстояние в тысячи километров через этот северный сухопутный мост. В том числе, и такие биологические виды, которые сегодня встречаются только в теплом климате. Никак иначе (кроме как миграциями целых толп разных биологических видов и родов) высокую общность фауны и флоры Голарктики в рамках естественной эволюции просто не объяснить. Понятно, что по ходу маршрута этих «великих переселений», абсолютно все популяции многих видов – вымерли. Видимо, специально для того, чтобы полностью заметать следы своих «великих переселений». Например, современные представители семейства верблюдовых сегодня живут только в Африке и Азии (два вида верблюдов) и в Южной Америке (ламы и викуньи), хотя центром их происхождения считается Северная Америка. Где никаких верблюдовых сегодня не наблюдается, а наблюдаются лишь ископаемые останки неких древних (т.е. других) видов верблюдовых. И так далее.
Но давайте вернемся к нашему роду Pant hera.
Итак, несмотря на все эти потрясающие миграции, и соответственно, целую кучу изолированных (и воображаемо эволюционирующих) популяций от Африки до Индонезии - лев, леопард и тигр продолжают хорошо отличаться друг от друга на всем протяжении своих ареалов.
Как такое может быть, если теория эволюции учит нас, что биологический вид – это всего лишь эфемерное (с точки зрения эволюции) понятие, а на самом деле, эволюционируют разные популяции?
Такое может быть только в том случае, если тот или иной биологический вид, воображаемо зародившись где-то в одной местности, потом очень быстро (быстрее, чем его популяции эволюционируют) распространится по всему ареалу, известному сегодня для этого вида.
Но если это так, то получается, что всё сегодняшнее распространение рода Panthera – это история сплошных грандиозных миграций и «великого переселения народов» (в сравнительно недавнем прошлом). Причем совершенно непонятно, почему только лишь одна исходная популяция (воображаемо эволюционировавшая, например, до современного леопарда) произвела затем такую огромную географическую экспансию? Куда делись (с её пути) все остальные (исконные) популяции воображаемо эволюционировавших крупных кошек (или других хищников) на такой огромной территории? Причем эволюционировавших прямо на месте, следовательно, более приспособленных (теоретически) к этому самому «месту». Или нам следует считать, что в то время, когда где-то в Азии появился современный леопард, на огромных территориях от Азии до Южной Африки и до Западной Европы – экологическая ниша этого высшего хищника просто пустовала? Пустовала настолько, что леопард сумел осуществить «марш-бросок» из Азии в Африку и Европу гораздо быстрее, чем сам (в свою очередь) разделился на пару десятков разных видов, в соответствии с единственно верным учением?
В принципе, такой сценарий еще не кажется невероятным в отношении только одного леопарда. Или даже всего рода Panthera. Действительно, бывает же такое, что абсолютно все изолированные популяции воображаемых «общих предков и потомков» в Северном полушарии вымирают в силу разных случайных причин, кроме трех конкретных популяций, которые в итоге и занимают всё северное полушарие в виде: льва, леопарда и тигра (при этом одинаково хорошо отличаясь друг от друга на громадных пространствах). То есть, такой сценарий «великих вымираний» и «великих переселений» еще можно предположить в отношении каких-то отдельных родов для объяснения наблюдаемой дискретности этих биологических видов.
Но когда точно такую же картину демонстрирует огромное число самых разных биологических видов (не только животных, но и растений), то поневоле возникает мысль – а чего это наблюдаемые сегодня биологические виды каждый раз так «мотало» и «швыряло» по свету, что мы теперь хорошо видим границы между разными видами… одновременно сумев обнаружить только пять (!) пресловутых «кольцевых ареалов» (с действительно «плавными переходами между популяциями») на всю живую природу. Причем половина из этих пяти установленных примеров уже раскритикована (см. выше).
Следует отметить, что помимо льва, леопарда и тигра, в Старом Свете сегодня обитает еще одна крупная кошка – снежный барс (Uncia uncia). Снежный барс тоже сохраняет свою видовую индивидуальность на всём ареале – никаких «полубарсов-полутигров» (или «полулеопардов») мы тоже нигде не наблюдаем.
Наконец, абсолютно такая же картина наблюдается в отношении еще одной крупной кошки – пумы (Puma concolor), уже в Северной Америке. Несмотря на то, что ареал пумы является крупнейшим из всех млекопитающих Нового Света, охватывая всю Южную Америку и большую часть Северной.
Или возьмем для примера медведей.
Предполагается, что разные виды медведей произошли от некоего общего предка, путем (конечно же!) постепенного накопления различий между разными популяциями этих медведей. И в результате получились (возьмем четыре современных вида) – бурый медведь (Ursus arctos), белый медведь (U. maritimus), черный американский медведь (U. americanus) и белогрудый (гималайский) медведь (U. thibetanus). Причем срок разделения медведей на белых и бурых методами молекулярной филогении еще совсем недавно оценивался всего примерно в 120 тысяч лет. И только недавно этот срок «удревнили» примерно до 600 тысяч лет. Видимо, специально «по просьбе трудящихся», потому что бурые и белые медведи экологически, морфологически и физиологически отличаются друг от друга слишком сильно, чтобы подобные различия могли возникнуть всего за 100 тысяч лет у таких крупных зверей в рамках единственно верного учения.
Итак, предположим, что изолированные популяции некоего «общего предка всех медведей», «размазанные» по огромной территории всей Голарктики, постепенно накапливали различия между собой. Всё накапливали и накапливали. И в конце концов, некоторые популяции стали белыми медведями, некоторые – бурыми, некоторые черными, а некоторые – белогрудыми. Но если так было на самом деле (а не только в нашем воображении, воспаленном идеями эволюции всего и вся), тогда мы и сейчас должны наблюдать примерно такой же размах изменчивости между разными популяциями озвученных видов медведей. Размах, доходящий до того, что для каких-то отдельных популяций будет уже не очень понятно – это всё еще бурый медведь, или уже белый (или уже черный). Ну и где же мы наблюдаем такую картину? На самом деле, бурый медведь – он и в Африке бурый… пардон, не в Африке, конечно. А, например, и в сибирской тайге, и в американских лесах. Где бурый медведь, кстати, сожительствует с американским черным медведем на больших территориях, но тем не менее, заметно от него отличается… зато схож с бурым мишкой из далекой России. В то время как черный американский медведь, кажется, больше похож на гималайского медведя. Ну а гималайский медведь (видимо, специально для того, чтобы еще раз подтвердить дискретность и устойчивость понятия биологический вид) на всём ареале своего современного распространения (от Ирана на западе до Дальнего Востока, Кореи и Вьетнама на востоке) обязательно имеет на груди хорошо заметное белое пятно характерной формы. За что и получил второе свое название (белогрудый медведь). Несмотря на то, что в целом окраска шерсти у медведей может серьезно варьировать даже в пределах одного вида.
Однако наиболее широко распространен по свету, конечно, бурый медведь. Он остаётся бурым медведем даже в пустыне Гоби! Интересно, почему изолированная гобийская популяция бурых медведей, оказавшаяся в столь экстравагантных (для бурого медведя) природных условиях, тем не менее, никуда не эволюционирует (во всяком случае, вплоть до той минуты, пока я пишу эти строки). Ведь специалисты относят эту популяцию к виду бурый медведь (Ursus arctos). Некоторые специалисты выделяют гобийского медведя в отдельный подвид - Ursus arctos gobiensis. Но при этом (по дружному согласию специалистов) гобийский медведь находится именно в рамках Ursus arctos (бурый медведь).
Итак, бурый медведь остаётся верным себе (бурым медведем) в достаточной степени, чтобы специалисты признавали его именно бурым медведем на всем протяжении громадного ареала обитания этого зверя. Невзирая на то, живет ли он в центрально-азиатской пустыне, или же где-нибудь на берегах острова Кадьяк, вблизи берегов Аляски.
Но стоит нам только взглянуть на белого медведя, как мы сразу же видим целый ряд четких признаков, как морфологических, так и физиологических (и конечно, экологических), по которым можно безошибочно разделить белого медведя и бурого. И это несмотря на то, что данные медведи (воображаемо) разошлись в ходе эволюции совсем недавно. И даже несмотря на то, что они могут свободно скрещиваться друг с другом, образуя плодовитое потомство. Причем северная граница распространения бурого медведя может подходить вплотную к границе распространения белого медведя на огромном протяжении их ареалов. Тем не менее, мы почему-то не наблюдаем никаких популяций «бело-бурых медведей» (или еще каких-нибудь не совсем «понятных» переходных популяций) на всем огромном ареале, где эти медведи могли бы соседствовать.
Таким образом, перечисленные факты можно, конечно, объяснять как угодно (задним числом), но против самого факта здесь уже не попрёшь - разные виды медведей демонстрируют вполне четкую видовую дискретность. Понятно, что такая картина совсем не льёт мельницу на воду дарвинизма. Скорее уж, намекает на нечто противоположное дарвиновской концепции.
Другие примеры непонятного (в рамках дарвинизма) сохранения биологическим видом своей специфичности на гигантских географических пространствах.
Бабочка Hypolimnas misippus из семейства нимфалид – распространена в тропических областях Африки, Азии и Австралии. Понятно, что самки этого вида, живущие в Австралии, сегодня вряд ли выходят замуж за самцов, обитающих где-нибудь в Сьерра-Леоне (Западная Африка). То есть, в данном случае имеется просто замечательная географическая изоляция, которая, в рамках современного дарвинизма, должна приводить к постепенному накоплению различий между изолированными популяциями, и в итоге, к образованию новых видов. К сожалению, мы не знаем, как давно эти бабочки изолированно обитают в Африке, Азии и Австралии.
Но если сравнить этих бабочек с современным человеком, то сегодня считается, что современный человек возник в Африке примерно 200-150 тысяч лет назад, а потом достиг Австралии примерно 50 тысяч лет назад. Получается, что на путешествие в Австралию у современного человека ушло от 150 до 100 тысяч лет (а надо сказать, что современный человек – весьма мобильный биологический вид).
Если принять такие же цифры «достижения Австралии» и для озвученного нами вида бабочки Hypolimnas misippus, тогда, учитывая скорость смены поколений у этих бабочек (6-7 поколений в год, см. Atluri et al., 2013), за 100 тысяч лет у этих бабочек должно было смениться примерно столько же поколений, сколько поколений сменилось бы у людей более чем за 10 миллионов лет!
Даже если предположить, что эта бабочка попала в Австралию всего 20 тысяч лет назад, то даже за это время у неё сменилось столько же поколений, сколько бы сменилось у человека за 2.5 миллиона лет. Но за два с половиной миллиона лет (согласно современным эволюционным представлениям), какие-то древние линии тропических обезьян (австралопитеков?) успели превратиться в современного человека. То есть, приобрели массу морфологических отличий. На уровне разных биологических родов (как минимум). А вот бабочки Hypolimnas misippus за то же самое число поколений (предположительно) почему-то не соизволили разделиться хотя бы на три разных биологических вида (например, африканский, азиатский и австралийский).
Можно, конечно, предположить, что эти бабочки попали в Австралию из Африки (или наоборот) не 20.000 лет назад, а всего, допустим, 5 тысяч лет назад (что равносильно примерно 500 тысячам лет для человека). И по этой причине еще просто не успели разделиться на разные виды. Но такие предположения высказывать очень опасно. Особенно в присутствии коварных креационистов. Потому что, учитывая массовый характер полностью аналогичных примеров (дискретности многих современных видов на огромных ареалах) – нам придется предположить, что примерно половина той картины распределения жизни, которую мы наблюдаем сегодня - на самом деле, сложилась совсем недавно (в недавнем геологическом прошлом). То есть, эта картина не свидетельствует в пользу представлений о «миллиардах лет эволюции» (не свидетельствует от слова совсем), а указывает на гораздо меньшие периоды времени. Поэтому коварные креационисты, наслушавшись таких выводов (о совсем малом возрасте ареалов очень многих биологических видов) начнут говорить, что такая картина, скорее, свидетельствует в пользу младоземельной концепции. Или даже в пользу потопной версии (быстрое занятие биологическими видами территорий, освободившихся после некоей глобальной катастрофы в совсем недавнем геологическом прошлом). Причем вторая половина картины наблюдаемого биологического разнообразия (всякие там эндемики и виды с ограниченными ареалами), в принципе, тоже не противоречит этой версии.
На всякий случай, озвучу пару примеров упорного сохранения видовой специфичности еще и среди растений. Очень многие виды растений имеют ареалы огромной протяженности. Например, ареалы большого числа видов растений протягиваются через всю Евразию. Некоторые заходят еще и в Северную Америку. А отдельные биологические виды вообще являются растениями-космополитами, то есть, распространены чуть ли не по всему свету.
Давайте посмотрим, например, на ареалы некоторых видов из семейства орхидных, живущих в умеренных широтах северного полушария. Выбираю почти случайно, в пределах одного и того же рода растений. Род Башмачок (Cypripedium):
1. Вид Башмачок обыкновенный (Cypripedium calceolus). Ареал распространения – широко распространен в Евразии от Британских островов и Испании до Тихого Океана. И на всей этой территории – башмачок обыкновенный представлен одним и тем же видом.
2. Вид Башмачок крупноцветковый (Cypripedium macranthos). Ареал: Белоруссия, европейская часть России, Северный Казахстан, юг Сибири, Дальний Восток, Монголия, Китай (включая остров Тайвань), Корейский полуостров, юг Камчатки, Курильские острова, Япония.
Как видим, ареалы этих двух видов перекрываются на огромном протяжении. Тем не менее, оба этих вида сохраняют свои видовые признаки (и отличаются друг от друга) на всей этой громадной территории (Рис. 40): [107]

Рисунок 40. Слева башмачок обыкновенный (Cypripedium calceolus), справа башмачок крупноцветковый (C. macranthos).
Или можно взять разных представителей рода Ятрышник (Orchis), которые встречаются по всей Европе, а многие виды распространены еще и в Азии (вплоть до Монголии и Тибета), а также в северо-западной Африке. При этом ареалы разных видов этого рода широко перекрываются. Тем не менее, вместо наблюдаемого «континуума свободно эволюционирующих популяций», специалисты как-то различают (друг от друга) конкретные виды на всем протяжении их естественных ареалов. Возможно, не без трудностей (различают). Но и идея «непрерывного континуума разных популяций» среди систематиков - тоже отнюдь не популярна. Хотя, казалось бы, в свете того механизма образования видов, который предлагается современной теорией эволюции - этой идеей (о «непрерывном континууме разных популяций») современная систематика должна быть буквально пропитана. Более того, должна быть популярной и мысль о бессмысленности (формального) выделения разных видов в этом континууме. Однако такие идеи почему-то почти не слышны. Неужели лишь потому, что систематики боятся потерять работу?
Ну а такой представитель семейства орхидных, как биологический вид Malaxis monophyllos, распространен вот на таких территориях: в Европе: Германия, Италия, Польша, Скандинавия, Украина, Россия; в Азии: Россия, Китай, Япония, Непал и даже Филиппины; в Северной Америке: южная Аляска, южная Канада, Новая Англия и регион Великих озер, изолированные популяции в Колорадо и Калифорнии.
То есть, мы опять видим распространение биологических видов на огромные территории с сохранением их видовой специфичности. Каким образом эта видовая специфичность поддерживается на столь большом географическом протяжении, если с точки зрения современной теории эволюции – биологический вид - лишь иллюзорная сущность, заключающаяся просто в значительном накоплении различий между разными популяциями, изолированными друг от друга, с течением времени?
Интересно, что семейство орхидных выделяется из других семейств растений огромным разнообразием видов (около 25000). Причем высокое число видов орхидей наблюдается во всех тропических регионах Земли (тропики Америки, Африки, Мадагаскара, Азии). Например, в одной только тропической Америке сосредоточено около 8000 видов орхидей. Хотя в высоких широтах орхидные распространены столь же широко (практически по всему миру), как и в тропических. Тем не менее, в высоких широтах число видов орхидей гораздо меньше.
Таким образом, мы видим в тропиках «распад» орхидных на очень большое число родов и видов, многие из которых имеют весьма скромный ареал распространения. Такая картина куда больше вписывается в видообразование по механизмам, предложенным современной теорией эволюции, чем северные виды орхидей с их громадными ареалами (у многих видов).
В связи с этим непонятно, почему так много видов орхидей в тропиках (в том числе, с очень ограниченными ареалами), и одновременно намного меньше видов в более высоких широтах при (одновременно) широком распространении многих из этих видов на огромные расстояния. Какая сила так «надирала» разнообразиться орхидеи в тропиках, в то время как в более высоких широтах – не надирала? И самое главное, почему? Или наоборот - если потрясающее воображение разнообразие видов орхидей в тропиках – это нормально с точки зрения теории эволюции, тогда какая же сила удерживает виды орхидных от такого же «распада» на громадных территориях Северного полушария?
В рамках концепции непрерывного творения, факты устойчивости биологических видов на огромных географических территориях могут быть объяснены довольно просто. Можно предположить, что биологический вид – это отнюдь не иллюзорный, а вполне реальный биологический объект. Причем многие такие объекты имеют серьезную степень «помехоустойчивости» («защиты от повреждений временем»). Что и позволяет им существовать на значительных территориях значительное время. После того как над ним соответствующим образом «поработают» (создадут).
В рамках же современного дарвинизма, сохранение единообразия у того или иного биологического вида на огромных территориях может быть объяснено (еще раз) только одним способом – если считать, что этот вид успел распространиться по всей обсуждаемой территории существенно быстрее, чем накапливаются различия в его изолированных популяциях. Поэтому в будущем, этот вид, конечно же, «развалится» на разные виды, но пока еще просто не успел.[108]
В некоторых случаях такое объяснение (задним числом) вполне можно допустить. Например, в случае с орхидеями, мы не знаем точно, сколько времени эволюционировали тропические орхидеи, дробясь на 8000 видов в тропической Америке. Может быть, на это у них ушло 50 млн. лет? А вот орхидеи умеренных широт, может быть, распространились по всей той территории, которую они занимают, допустим, всего за 500 тысяч лет? Вот поэтому мы и не наблюдаем такого множества видов орхидей в высоких широтах? А вот если бы орхидеи в северных широтах «сидели бы на своих местах» те же 50 млн. лет, то может быть, у них бы тоже образовалось 8000 видов?
Что же, такое теоретическое допущение вполне возможно. Выше я уже говорил, что объяснения задним числом имеют такую специфическую особенность – они приводят к успеху практически всегда.
Тем не менее, некоторые биологические факты могут серьезно «подпортить кровь» даже таким объяснениям. Например, если мы возьмем такой род орхидей, как Calanthe (насчитывающий около 200 видов), то, во-первых, обнаружим, что этот род орхидей приурочен к тропическим областям Земли, с преимущественным разнообразием в Юго-Восточной Азии. Таким образом, в рамках эволюционных представлений, следует предположить, что место эволюционного происхождения этого рода – Юго-Восточная Азия. Тем не менее, это обстоятельство отнюдь не мешает разным видам этого рода быть распространенными в тропических и субтропических широтах чуть ли не всего Земного шара: в Китае, Индии, на Мадагаскаре, в Австралии, в Мексике, Центральной Америке, и даже на островах Карибского моря. Это обстоятельство как-то не вписывается в только что сделанное нами предположение, что тропические орхидеи «все 50 млн. лет сидели (эволюционируя) строго на своих местах».
А во-вторых, некоторые виды орхидей из этого рода широко разбросаны по самым разным островам и странам Тихого и Индийского океанов. То есть, эти виды имеют крайне разорванные ареалы (следовательно, представленные превосходно изолированными популяциями). И тем не менее, даже «сидя» на десятках удаленных друг от друга тропических островов (и еще где-нибудь в Индии и Малайзии), разные популяции этих видов почему-то не эволюционировали в разные виды, а сохраняют свои видовые признаки.
Давайте оценим, например, ареал тропической орхидеи Calanthe triplicata. Этот вид орхидеи найден: на острове Маврикий, на Мадагаскаре, на Сейшелах, в Ассаме (восточный штат Индии), в восточных Гималаях, в южной Индии, на острове Шри Ланка, в Бирме, Таиланде, Малайзии, Лаосе, Камбодже, южном Китае, Вьетнаме, на острове Борнео, на Яве, на Малых Зондских островах, на островах Молуккского архипелага, на Филиппинских островах, на Сулавеси, Суматре, архипелаге Бисмарка, в Новой Гвинее, на Соломоновых островах, на острове Норфолк, в восточной Австралии, на островах Фиджи, в Новой Каледонии, на островах Самоа, Вануату, островах Валлис и Футуна, на Маркизских островах (то есть, вообще в центре Тихого океана), на островах Санта-Круз, на Каролинских островах, на Марианских островах, на Тайвани и на островах Рюкю.
Причем этот вид орхидеи отнюдь не привязан к каким-нибудь приморским местообитаниям (для облегчения распространения через тысячи километров морской воды). Эта орхидея произрастает во влажных тенистых лесах. Причем эти леса обычно приурочены к возвышенностям (с высотой над уровнем моря от 500 до 1500 метров).
Ну и где же «эволюция в условиях изоляции» (в том числе, в условиях островной изоляции)? Видимо, этому виду орхидеи просто забыли сказать, что в рамках современной теории эволюции ему уже давно надо разделиться на сотню разных видов?
Наконец, в некоторых случаях озвученное выше объяснение про «50 млн. лет сидения на одном месте vs 500 тыс. лет распространения по всему Земному шару»… такое объяснение в некоторых случаях не подходит еще и по другой причине.
Дело в том, что в некоторых случаях мы точно знаем возраст того или иного биологического вида.[109]
Например, в случае всё тех же цихлид озера Виктория – точно установлено, что им для образования сотен (!) биологических видов понадобилось вовсе не 50 млн. лет… а только 15 тысяч. В свете этого факта, рассуждения о том, что щукам, окуням, ершам, сигам и колюшкам «просто не хватило времени на видообразование» - звучат уже далеко не так убедительно (если не сказать – забавно).
Другой интересный пример. В северном полушарии живет такой вид жука - сибирский морщинник (Helophorus sibiricus). Ареал обитания этого вида столь же огромен, как и в случае уже обсуждавшихся выше видов. Этот жук широко распространен на большей части Голарктики. Его современный ареал начинается от северных частей Скандинавии, проходя по европейской части России, далее через всю Сибирь и Дальний Восток, и продолжается еще дальше, уже в Северную Америку, занимая там практически всю Аляску. В целом, распространение этого вида соответствует таежному биому в Евразии, но превышает его границы, заходя в зону тундры на севере, а на юге доходя до лесов и лугов северного Китая, Монголии и японского острова Хонсю (Fikacek et al., 2011).
Кроме того, было найдено немало ископаемых остатков этого вида из сравнительно недавнего прошлого.[110] Эти находки показывают, что в сравнительно недавнем прошлом данный вид жука имел еще большее распространение (почти кругосветное) – от Британских островов до региона Великих Озер (восточная часть Северной Америки).
Получается, что на всей этой громадной территории данный вид жука сохраняет свои характерные видовые признаки. То есть, мы имеем еще один пример огромного географического распространения с сохранением видовой специфичности.
Однако это только «присказка». «Сказка» началась тогда, когда была сделана еще одна палеонтологическая находка, датируемая уже не сравнительно недавним прошлым, а ранним миоценом (то есть, примерный возраст 20 млн. лет). Обнаруженные ископаемые остатки оказались достаточно подробными, чтобы авторы соответствующей работы (Fikacek et al., 2011) смогли сделать уверенный вывод о принадлежности этих остатков… именно к современному виду жука Helophorus sibiricus. То есть оказалось, что этот вид вообще никак не изменился (морфологически) за последние 20 млн. лет!
Причем и место находки тоже оказалось близким к современному ареалу обитания этого вида (Рис. 41):
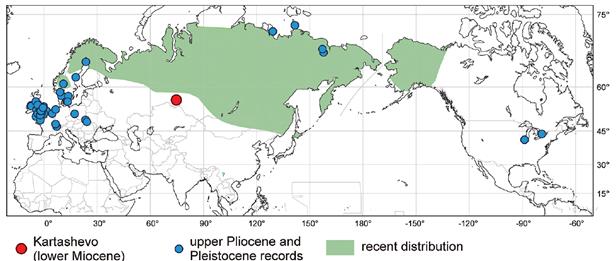
Рисунок 4 1 . Распространение жука Helophorus sibiricus . Зеленым цветом показан современный ареал вида. Синими кружками – находки этого вида жука из верхнего плиоцена и плейстоцена. Красный кружок – обсуждаемая палеонтологическая находка этого вида – морфологически неотличимые от современного вида H. sibiricus ископаемые остатки с примерным возрастом 20 млн. лет. Рисунок взят из работы (Fikacek et al., 2011).
Таким образом, в случае этого вида жука мы наблюдаем отсутствие какой-либо «эволюции изолированных популяций» на огромных географических пространствах, так же как и в предыдущих случаях. Но в этом конкретном случае мы уже не можем привлечь наше предыдущее «объяснение» этого факта - быстрым распространением этого вида по всему современному ареалу. Потому что оказалось, что этот вид жука остаётся неизменным не только на огромных географических пространствах, но еще и на протяжении огромных промежутков времени.
Таким образом, этот конкретный пример предельно наглядно опровергает (по факту) постулат современной теории эволюции, что биологические виды происходят путем постепенного накопления изменений в изолированных популяциях. И наоборот, ярко показывает - биологические виды могут прекраснейшим образом сохранять свою видовую специфичность, несмотря на любое прошедшее время и любое расстояние (между изолированными популяциями).
Следует особо подчеркнуть, что этот жучок живёт отнюдь не в экваториальном лесу Африки. Это обитатель наших северных широт. Следовательно, за прошедшие 20 млн. лет он должен был пройти сквозь «огонь, воду и медные трубы». А если точнее, то сразу через несколько глобальных ледниковых периодов (т.е. кардинальных изменений климата), связанных с этим резких изменений в составе и распространении растительности, многочисленные смены ареала и через многое другое. В общем, по идее (в рамках «единственно верного учения») популяции этого вида должны были за это время раздробиться на такое число разных видов, в которых мы бы неизбежно «заблудились». И выудить из этого «леса эволюционировавших популяций» именно этого жучка (именно с этой морфологией), было бы всё равно, что выудить иголку из стога с сеном.
Действительно, род (Helophorus), к которому принадлежит данный вид, сегодня насчитывает в своем составе почти 200 видов - 150 видов на территории Евразии и Северной Африки, более 40 видов в Северной Америке, еще несколько видов живут в Центральной Америке, тропической Африке и в северной Индии. Казалось бы, этот факт косвенно свидетельствует о том, что биологические виды могут свободно эволюционировать: изменяться, дробиться и разделяться, хотя бы в рамках рода. Однако, несмотря на столь впечатляющее видовое разнообразие внутри этого рода, конкретный вид (этого рода) морщинник сибирский (H. sibiricus) - сохраняет свои видовые особенности на всем протяжении громадного ареала. И как мы теперь выяснили, еще и на огромных промежутках времени. Давайте еще раз прочувствуем всю «интригу» этой ситуации - палеонтологи нашли некоего ископаемого жучка возрастом 20 миллионов лет… и он оказался морфологически неотличим от современного жука, морщинника сибирского. Таким образом, этот героический жучок наглядно демонстрирует потрясающую устойчивость биологического вида не только в пространстве, но еще и во времени. Устойчивость настолько сильную, что это просто необъяснимо в рамках «единственно верного учения», где как известно, «всё течет, всё изменяется» (особенно среди куч изолированных популяций).
И всё бы хорошо, если бы такой жучок был один. Тогда его можно было бы списать на «очень удачное стечение эволюционных обстоятельств». Однако таких «жучков» в настоящее время палеонтология «накопала» уже достаточно много. Приведу соответствующую цитату (Расницын, 2002):
Известны и более прямые указания на большую древность некоторых видов. Так, в фауне эоценового балтийского янтаря (возраст не менее 40 млн. лет) сейчас известно несколько видов насекомых и около десятка видов клещей, на видовом уровне неотличимых от современных; более обычно персистирование на интервалах в 10 — 20 млн. лет (Zherikhin, 1999). Находки разнообразных плиоценовых галлов, неотличимых от галлов, вызываемых современными видами насекомых на тех же растениях, гораздо моложе (3 — 5 млн. лет), но они не менее важны, так как подтверждают широкое распространение не только морфологического, но и биохимического стазиса насекомых в течение миллионов лет (Zherikhin, 2002a). Многие из этих находок происходят из Северной Европы, так что речь идет о видах, переживших ледниковый период, но не изменившихся даже биохимически (поскольку морфологическая специфика галла обусловлена биохимическим воздействием насекомого — галлообразователя на ткани растения — хозяина). Таким образом, стазис длиной в миллионы лет осуществляется не только в отсутствие обмена генами, но и вопреки несомненным глубоким, возможно даже катастрофическим, изменениям условий.
Итак, некоторые виды насекомых и клещей, похоже, чихать хотели на теорию эволюции. И что-то мне подсказывает, что таких насекомых на самом деле, гораздо больше. Просто палеонтология – это такая наука, которая находит далеко не всех насекомых, существовавших когда-либо, и существующих до сих пор. Действительно, упомянутый в цитате балтийский янтарь возрастом 40 млн. лет сегодня является отнюдь не единственным источником ископаемых видов насекомых, которые… неотличимы от современных видов.[111]
Но самым большим монстром в этом отношении является, конечно, уже упоминавшийся нами выше щитень летний (Triops cancriformis). Всё-таки 230 млн. лет вообще без всяких изменений, несмотря на обитание во временных водоемах – это уже абсолютная наглость с точки зрения единственно верного учения.
В связи с этим у меня есть предложение по щитню летнему. С одной стороны, пример щитня летнего (сам по себе, без каких-либо других примеров) уже хоронит теорию эволюции по факту. Потому что по факту отсутствуют какие-либо изменения биологического вида вот уже на протяжении 230 млн. лет, несмотря на бесчисленную смену экологических условий. С другой стороны, щитень, несмотря на свою полную безобидность для человека, имеет весьма устрашающую внешность, напоминающую личинку «чужого» (из одноименного фантастического фильма). Поэтому у меня есть предложение - назвать щитня летнего Чудовищем Теории Эволюции (сокращенно, ЧТЭ).
Действительно, при появлении ЧТЭ все постулаты современной теории эволюции сразу же куда-то деваются. Видимо, микроэволюционные факторы (случайные мутации, дрейф генов и естественный отбор, принятые в этой теории в качестве «двигателей эволюции») - просто шарахались от щитня летнего врассыпную (все эти 200 млн. лет) из-за его устрашающей внешности?
В лице ЧТЭ я дарю читателям поистине страшное оружие против верующих дарвинистов. Особенно против тех из них, которые разбираются в биологии. Для тех, кто хорошо понимает, что такое биологический вид, и чем биологический вид отличается, например, от биологического семейства, щитень летний – это как удар ядерной баллистической ракеты (по теории эволюции).
Попробую пояснить, почему это так.
Дело в том, что разные биологические виды отличаются друг от друга, как правило, мелкими морфологическими признаками, часто не имеющими какого-либо особенного биологического значения. Поэтому с чисто биологической точки зрения, эти признаки – нейтральны (для выживания).
Можете посоветовать верующему дарвинисту поизучать, какими конкретно признаками щитень летний (Triops cancriformis) отличается от других представителей рода Triops. Или, например, от щитня весеннего (Lepidurus apus). И после соответствующего изучения, пусть теперь дарвинист попробует придумать биологически целесообразные причины этих различий. Такой приём гарантировано поставит верующего дарвиниста в ступор.
Давайте посмотрим, в чем же состоят различия.
Вообще, в семействе щитней имеется два разных рода. Род Triops (к которому относится щитень летний), и род Lepidurus. И в роде Triops, и в роде Lepidurus насчитывается несколько разных видов. Итак, чем же щитень летний (Triops cancriformis) отличается от других видов щитней, например, от Triops granarius?
Открываем «Определитель пресноводных беспозвоночных России и сопредельных территорий» и читаем: [112]
Ключ для определения видов
1(2) На сегментах аподуса отсутствуют дополнительные шипы, на дорсальной стороне тельсона имеется 1-4 крупных серединных шипа, располагающихся строго в ряд ……….. T. cancriformis Bosc, 101 (Табл. 7) Палеарктика.
2(1) На сегментах аподуса наряду с крупными основными имеются мелкие дополнительные шипы, на дорсальной стороне тельсона имеется не менее 5 мелких серединных шипиков, располагающихся в неровный ряд ……………… T. granarius ( Lucas, 1864) ( Tabl. 7) Забайкалье.
Как видим, различие между этими видами производится на основании весьма мелких морфологических признаков, которые вряд ли имеют большое биологическое значение. Я понимаю, что всякие там «аподусы» и «мелкие шипы на тельсоне», на слух звучат для неспециалиста весьма туманно. Поэтому привожу еще и соответствующий рисунок из этого же определителя, чтобы читатель сам смог оценить масштабы этой разницы, между вооружением тельсонов[113] двух озвученных видов щитней (см. на рисунке 42 изображения 9 и 11-12):
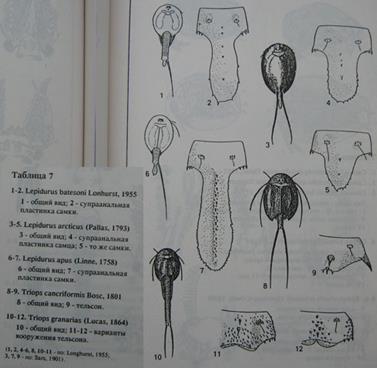
Рисунок 4 2 . Страница из определителя с рисунками диагностических признаков разных видов щитней. Под цифрами 9,11,12 показаны варианты вооружения тельсона у двух разных видов щитней. Вот этими шипиками и различаются T. cancriformis (9) от T. granarius (11,12).
Кроме того, на этом рисунке я хорошо вижу разницу в общих пропорциях тела между двумя озвученными видами (на рисунке 42 изображения 8 и 10). T. granaruis на этом рисунке имеет существенно более удлиненное тело, чем T. cancriformis. Впрочем, я не являюсь специалистом по щитням, поэтому отнюдь не настаиваю на этой разнице. Может быть, это просто так на рисунке получилось, а на самом деле, более «длинные» и более «короткие» особи встречаются и среди индивидуальных представителей обоих видов. В любом случае, эта разница в общих пропорциях тела, тоже, наверное, биологически нейтральна. Ну а про отличия в вооружении тельсонов, выражающиеся в микроскопической (для стороннего человека) разнице между числом и размером шипиков, я уже вообще не говорю. Понятно, что эта разница вряд ли играет сколько-нибудь заметную биологическую роль. Тем более что и тот и другой вид – оба живут, по сути, в одних и тех же условиях - временных пересыхающих водоемах (больших лужах, канавах, всяких заболоченных низинах и тому подобное).
Еще более интересно, что не только разные виды рода Triops, но даже и разные роды щитней (Triops и Lepidurus) различаются между собой столь же «важными» биологическими признаками. Давайте опять заглянем в определитель и прочитаем:
КЛЮЧ ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ
1(2). Супраанальная пластинка имеется …….. Lepidurus Leach, 1819 (стр. 28)
2(1). Супраанальная пластинка отсутствует ……. Triops Schrank, 1803 ( cтр. 29)
Вот так. Получается, что даже биологические роды иногда (а на самом деле, очень часто) различаются между собой такими морфологическими признаками, разница между которыми никак не сказывается на выживаемости организмов.
Чтобы понять, что такое «супраанальная пластинка» - достаточно посмотреть на рисунок из определителя, который я уже привел выше. Там с 1 по 7 изображения показаны разные представители рода Lepidurus, а с 8 по 12 изображения нарисованы два разных представителя рода Triops.
Можно даже самим теперь потренироваться в определении щитней (до уровня рода). Давайте попробуем (Рис. 43):

Рисунок 43. Представители двух разных родов щитней (Triops и Lepidurus). Пояснения в тексте.
Слева никакой супраанальной пластинки нет (показано стрелкой), значит, это представитель рода Triops. А справа имеется хорошо выраженная супраанальная пластинка (показана стрелкой), следовательно, это представитель рода Lepidurus.
Понятно, что придумать какое-нибудь особенное биологическое значение этому признаку вряд ли получится. Ведь стоит только предположить, что представители рода Lepidurus имеют эту пластинку потому, что она им для чего-то нужна, как тут же с протестом выступят представители рода Triops, которые скажут, что эта пластинка «нафиг никому не нужна», потому что они, представители рода Triops, прекрасно без неё обходятся, обитая, по сути, в таких же экологических нишах, что и представители рода Lepidurus. В любом случае, даже если продолжать настаивать, что у этой пластинки, на самом деле, имеется какой-то глубинный биологический смысл, то всё равно нам придется признать, что во временных водоемах можно спокойно выживать и без этого «глубинного признака». Потому что представители рода Triops именно так и делают по факту. Следовательно, данный морфологический признак либо вообще биологически нейтрален, либо его роль очень незначительна. Поскольку можно выживать как с ним, так и без него.
Точно так же можно обратить внимание на некоторую разницу в форме панцирей у этих двух щитней (Рис. 44):

Рисунок 44. Представители разных родов щитней. Triops c лева, Lepidurus справа.
Кроме того, рельеф панциря Lepidurus тоже выглядит несколько иначе и на вид более «гладкий», чем рельеф панциря Triops.[114] Понятно, что и эти различия тоже, скорее всего, биологически нейтральны.
Практически все виды щитней обитают примерно в одинаковых условиях – в мелких временных водоемах: больших лужах, канавах, заливных лугах (редко в постоянных водоемах), где у них почти нет естественных врагов (за исключением птиц, «выхватывающих» щитней сверху, и самих щитней, не чуждых каннибализма). Щитни могут различаться временем своего появления. Например, щитень весенний (Lepidurus apus) специализируется на ранних весенних лужах с холодной водой, а щитень летний (Triops cancriformis) предпочитает летние лужи, с водой более теплой. Вряд ли подобные экологические различия требуют от щитней именно разных панцирей, с разной формой и рельефом. Скорее всего, что эти морфологические различия между щитнями – просто биологически нейтральны.
Итак, давайте повторим еще раз - разные биологические виды часто различаются между собой мелкими, биологически нейтральными признаками (в том числе, и разные виды щитней).
Но нейтральные признаки не находятся под действием естественного отбора. Поскольку они безразличны для выживания.
Зато на такие признаки должны (теоретически) воздействовать другие эволюционные факторы: случайные мутации, дрейф генов и эффект основателя. Эти эволюционные факторы, в отличие от естественного отбора, имеют случайную природу. Например, постоянный поток случайных мутаций обязан постепенно изменять нейтральные биологические признаки. На этом, кстати, построена методика «молекулярных часов» (на уровне генов и белков).
В результате случайного дрейфа генов и эффекта основателя, разнообразные нейтральные мутации должны случайным образом закрепляться, или наоборот, удаляться из популяций. И чем больше этих популяций, тем быстрее будут изменяться нейтральные признаки. Чем больше популяций попадают в условия длительной изоляции – тем быстрее изменяются нейтральные признаки. Чем больше популяций попадает в ситуацию, когда новую популяцию «зарождает» всего парочка родителей (эффект основателя) - тем быстрее должны изменяться нейтральные признаки. Наконец, если часто происходит массовая гибель особей (и целых популяций) из-за колебаний условий среды, и при этом выживает считанное число отдельных особей (так называемые «бутылочные горлышки»)… тем быстрее должны изменяться биологически нейтральные признаки. Таким образом, биологически нейтральные признаки видов должны всё время как бы «эволюционно дрейфовать» в случайных направлениях.
В таких условиях вряд ли следует ожидать сохранения биологически нейтральных признаков в неизменном состоянии в течение хотя бы десятков тысяч поколений. А уж на протяжении 200 миллионов поколений это вообще невозможно.
Тем не менее, щитень летний именно так и поступает:
1. Он отличается от других видов щитней именно мелкими, биологически нейтральными признаками.[115]
2. Он обитает в настолько непостоянной среде обитания, насколько это вообще возможно.
3. Однако щитень летний возрастом 230 млн. лет – морфологически неотличим от щитня летнего, плавающего в современной луже.
В качестве конкретного примера возможного разнообразия форм панцирей, можно посмотреть на других (недавно обнаруженных) ископаемых щитней (Hegna & Ren, 2010). Хорошо видно, что и форма панциря, и общие пропорции тела у этих представителей щитней серьезно отличаются от форм и пропорций, характерных для щитня летнего (Рис. 45):
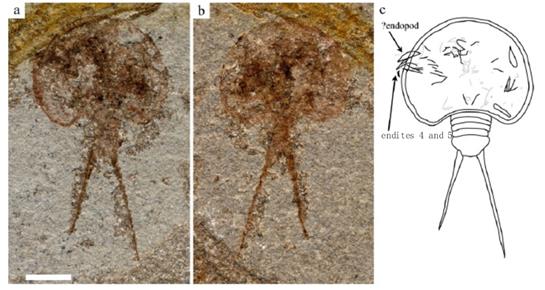
Рисунок 4 5 . Фотографии ископаемых остатков и соответствующий рисунок ископаемого вида Jeholops hongi, предположительно относящегося к щитням (Notostraca), из работы (Hegna & Ren, 2010РHHРвввв).
Представленный на рисунке вид (Jeholops hongi) был обнаружен в более поздних отложениях (примерно 125 млн. лет назад), чем остатки щитня летнего (230-220 млн. лет назад).
Таким образом, казалось бы, нет никаких особенных причин, из-за которых у щитня летнего должны непременно сохраняться именно его пропорции тела, его форма панциря и другие признаки. Тем не менее, щитень летний упорно сохраняет свои видовые признаки по факту, на протяжении всех 230 млн. лет с момента своего первого появления в палеонтологической летописи. Таким образом, щитень летний - это «шах и мат» современной теории эволюции.
Причем этому чудовищу дарвиновской теории мало опровергать эволюцию видов по факту. Жуткое создание при этом еще и нагло издевается над нашим (горячо любимым) учением.
Дело в том, что сегодня щитень летний демонстрирует впечатляющую внутривидовую изменчивость. Как между разными особями, так и между разными популяциями. Например, на сегодняшний день, вроде бы, признаны три разных подвида щитней летних: T. c. mauritanicus, живущий в Северной Африке (и еще в некоторых районах Иберийского полуострова); T. c. simplex, живущий, в основном, в Испании; и наконец, T. c. cancriformis, «типичный» щитень летний, живущий на огромной территории от Западной Европы (и Северной Африки) до Японии включительно.
Причем авторы, например, вот этой работы (Zierold et al., 2007) считают, что целый ряд эволюционных изменений у щитней летних[116] случился уже в ходе последнего ледникового периода, в позднем плейстоцене, то есть, максимум, за последние десятки тысяч лет. Таким образом, можно предположить, что щитень летний сейчас вроде бы… интенсивно эволюционирует!
Что и говорить, в страшное время мы с Вами живем. Щитень летний, не эволюционировавший вообще никак на протяжении 230 млн. лет… вдруг начал эволюционировать прямо сейчас. Наверное, близок Конец Света?
В общем, если бы Вы подвели к коллекции, составленной из разных экземпляров щитня летнего (да еще и собранных в разных удаленных районах), такого верующего дарвиниста, который бы не знал, кто такой этот щитень летний, то верующий дарвинист непременно ткнул бы в эту коллекцию пальцем, воскликнув при этом:
– Посмотрите, разные экземпляры, и разные популяции этого животного серьезно различаются между собой! Поэтому совершенно очевидно, что не пройдет и 1 млн. лет, как это животное изменится, и превратится в другой вид, или даже разобьется на несколько разных видов, под действием животворящей силы эволюции!
Однако по факту, несмотря на впечатляющую внутривидовую изменчивость, это чудовище теории эволюции так и не соизволило сэволюционировать хотя бы куда-нибудь ни за 1 млн. лет, ни даже за 230 млн. лет. Несмотря на то (повторяю еще раз специально для верующих дарвинистов), что обитая в крайне нестабильной среде, оно просто обязано было эволюционно изменяться.
Таким образом, получается полный эволюционный анекдот:
1). Смотрим на сегодняшние популяции щитня летнего. И видим именно такую картину, о которой нам рассказывает современное учение о микроэволюции – наблюдаемый размах внутривидовой изменчивости, доходящий на разных территориях даже до разных подвидов. Так и кажется - вот же она, «зарождающаяся» эволюция новых биологических видов в результате изоляции разных популяций.
2). А теперь «отодвигаемся» на 230 млн. лет назад, в триасовый период. И видим, что щитни летние триасового периода, найденные в ископаемом состоянии где-нибудь в североамериканском штате Вирджиния (Gore, 1986)… неотличимы от щитней летних, плавающих сегодня в лужах какой-нибудь Челябинской области.
Вот такая вот она, «очевидная эволюция».
Понятно, что когда мы говорим о «неотличимости», мы можем говорить только о наблюдаемых морфологических признаках. О каких-либо молекулярно-генетических фактах здесь говорить не приходится, поскольку сложная органика не может сохраняться сотни миллионов лет (впрочем, об этом мы еще подробно поговорим ниже).
Поэтому верующий дарвинист вполне может начать утверждать, что щитни летние из триаса ничем не отличаются от современных щитней летних… только морфологически. А вот генетически, щитень летний, может быть, ого-го как изменился!
В этом случае Вам следует сказать дарвинисту, что Вас совершенно не волнуют молекулярно-генетические вопросы. Вас волнует вопрос эволюции живых форм. И вот в этом отношении, щитень летний не изменился вообще никак. Хотя в рамках современной теории эволюции, щитень летний просто не смог бы пронести в неизменном состоянии (через все 230 млн. лет), те мелкие морфологические признаки, которыми он определяется именно как биологический вид Triops cancriformis.
Впрочем, в некоторых случаях, современную теорию эволюции можно «поймать» аналогичным образом даже на молекулярной генетике. Наверное, первым, кто это сделал, были Ю.П. Алтухов и Ю.Г. Рычков (Алтухов, Рычков, 1972). Авторы установили, что некоторые белки являются одинаковыми для разных популяций конкретного биологического вида, одновременно резко отличаясь от таких же белков у других, даже очень близких биологических видов. Это лучше всего продемонстрировать соответствующей иллюстрацией (Рис. 46):
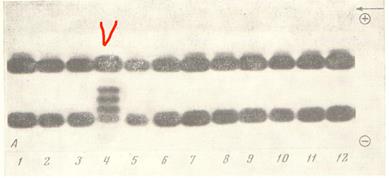
Рисунок 4 6 . Специфичность на видовом уровне гемоглобинов у близких видов рыб сем. S almonidae . 1-3, 5-12 – Кета (Oncorhynchus keta ); 4 – Кижуч ( O . kisutch ). Рисунок взят из работы (Алтухов, Рычков, 1972).
Если посмотреть на рисунок, то хорошо видно, что у разных особей кеты (Oncorhynchus keta) из разных популяций (?) электрофорезный «отпечаток» гемоглобинов примерно одинаков (цифры 1-3, 5-12). В то же время, видно, что у близкого вида - кижуча (Oncorhynchus kisutch) «отпечаток гемоглобинов» резко отличается от «отпечатка гемоглобинов» кеты (цифра 4 на рисунке, отмеченная галочкой).
Теория эволюции (СТЭ) говорит нам, что эти близкие виды (кета и кижуч) произошли от общего предка, благодаря постепенному расхождению разных признаков в разных популяциях. Именно такое, постепенное расхождение в изолированных популяциях, в конце концов, и привело к тому, что гемоглобины этих близких видов рыб столь резко различаются при электрофорезе (согласно современному эволюционному учению).
Но тогда эта теория предсказывает, что мы должны наблюдать сходные процессы еще и между разными популяциями (пока еще) единого вида. Однако мы этого не наблюдаем . А наблюдаем нечто противоположное - почти идентичность обсуждаемого признака в разных популяциях кеты. И одновременно, резкий «скачок» этого же признака при переходе от одного биологического вида к другому биологическому виду.
Создаётся даже ощущение некоего «отпечатка пальца» для конкретного биологического вида.
Таких белков, похожих на «отпечаток пальца» биологического вида, в данной работе был приведен целый ряд (Алтухов, Рычков, 1972). Понятно, что здесь нет ничего похожего на «образование вида за счет постепенного накопления различий между разными популяциями». Очевидно, что факты, опубликованные авторами, не вписываются в рамки СТЭ. Под влиянием этих результатов Ю.П. Алтухов даже сформулировал «закон генетического мономорфизма», в котором предположил, что гены организмов можно разделить на «полиморфные» (подверженные изменчивости) и «мономорфные», которые вообще не подвержены какой-либо изменчивости. К этой (мономорфной) части генома Алтухов предположительно отнёс и гены, отвечающие за некоторые видовые признаки. А под конец жизни Алтухов, кажется, вообще отказался от идеи изменяемости биологических видов.
«Генетический мономорфизм» Алтухова впоследствии был подвергнут критике на том основании, что автору не удалось собрать достаточно обширный материал в пользу своей гипотезы, то есть, доказать, что «генетический мономорфизм» является в природе правилом, а не исключением (найденным, допустим, только для озвученных Алтуховым белков).
Впрочем, какие-то гены, ответственные за видовые признаки – просто обязаны быть «мономорфными» (т.е. неизменяемыми). Это понятно чисто логически. Иначе бы мы не наблюдали в природе устойчивых биологических видов. То есть (опять возвращаемся к истокам) для того чтобы обыкновенная щука выглядела именно как обыкновенная щука на всем протяжении её громадного ареала – какие-то её гены по определению должны быть «мономорфными», отвечая именно за видовую принадлежность. Это тривиальный вывод, который прямо следует из факта видового постоянства на огромных ареалах.
Другое дело, дарвинизм утверждает, что со временем, это «видовое постоянство» должно постепенно исчезать под действием соответствующих эволюционных факторов (случайных мутаций, дрейфа генов, естественного отбора). Но, как известно, в науке утверждения принято доказывать фактами. А факты таковы, что дарвинизму здесь вряд ли позавидуешь. Достаточно взять некоторые из тех примеров, которые я уже приводил выше.
Например, изолированные популяции (сразу нескольких видов) бессяжковых насекомых, удаленные друг от друга на колоссальные расстояния (Калимантан, Япония, Южная Африка, Европа) – неотличимы друг от друга на видовом уровне. Хотя вероятность их трансконтинентальных миграций крайне мала (см. выше). Поэтому такие ареалы приходится объяснять пресловутым «дрейфом континентов» (Расницын, 2002). Отсюда следует, что возраст этих изолированных популяций должен быть никак не меньше 100 млн. лет.
Но для того, чтобы популяции, разметанные по всему Земному шару, оставались неотличимыми друг от друга на видовом уровне в течение сотни миллионов лет (если не в два раза больше) - для этого какая-то часть их генов должна быть просто потрясающе-бронебойно-«мономорфной» именно на видовом уровне.
Или (в последний раз) возьмем всё того же щитня летнего. Этот пример показывает, что характерные признаки вида, сами по себе являясь мелкими и нейтральными в биологическом отношении, тем не менее, сохраняются неизменными на протяжении 200 миллионов поколений. Я не знаю, что еще нужно для того, чтобы, наконец, признать, что СТЭ опровергнута фактами.
Впрочем, на этом факты, убийственные для синтетической теории эволюции, отнюдь не заканчиваются. СТЭ можно спокойно убить еще одной группой фактов. Как известно, понятие «популяция» в строгом смысле можно применить только к организмам, размножающимся половым путем. А вот к организмам, размножающимся бесполым способом (или, например, партеногенезом), понятие «популяция» в строгом смысле неприменимо. Потому что в случае бесполых организмов - нет общего генофонда популяции, который всё время поддерживается за счет свободного скрещивания между особями этой популяции.
В бесполой популяции, по сути, каждая новая особь, получившаяся, допустим, почкованием, теперь навсегда отрывается от всех своих «сотоварищей» по данному биологическому виду. Точно так же будет обстоять дело и в партеногенетических популяциях – каждая новая особь, вышедшая из яйца, отложенного партеногенетической самкой, навсегда «изолируется» от всех других особей этого вида. Поэтому каждую бесполую (или партеногенетическую) особь можно рассматривать сразу и как «бутылочное горлышко популяции», и как «предка-основателя новой популяции» в одном флаконе. То есть, считать родоначальницей целой линии организмов, которые будут содержать только гены, доставшиеся им от этой конкретной особи (с возможными случайными мутациями и дрейфом генов). И так каждый раз, после каждого акта размножения каждой такой особи.
Поэтому для бесполых или партеногенетических организмов (с точки зрения современного дарвинизма) понятие биологический вид должно быть еще более размытым, чем в популяциях организмов, размножающихся половым путем. Никакой дискретности биологических видов мы у таких живых существ наблюдать вообще не должны. А должны наблюдать некую кашу из разных линий организмов, с беспорядочными различиями между этими линиями (от небольших различий до самых серьезных). В результате, должно быть непонятно, где здесь один биологический вид, а где уже другой, третий или четвертый.
Это четкий прогноз теории эволюции в отношении живых существ, не имеющих полового размножения (или размножающихся строго партеногенетически).
А теперь давайте посмотрим, как обстоят дела на самом деле (Расницын, 2002):
С точки зрения синтетической теории эволюции совершенно необъяснима дискретность видов у партеногенетических и бесполых организмов, лишенных обмена генами, но она имеет место. Сравнение близких обоеполых и партеногенетических видов у коловраток (Майр, 1974) и жуков-долгоносиков (Иванова, 1978; В.В. Жерихин, личное сообщение), обоеполых и бесполых видов у протистов (Ю. Полянский, 1957; Poljansky, 1977), низших водорослей (В. Полянский, 1956) и папоротников (Farrar, 1990) показало, что однополые и бесполые виды столь же дискретны, как и обоеполые.
Итак, мы видим, что бесполые и партеногенетические живые существа из совершенно разных таксономических групп ведут себя одинаковым образом, дружно опровергая прогнозы дарвиновской теории эволюции. А именно, эти виды ничем не отличаются от видов, размножающихся половым путем по соотношению «дискретность/непрерывность».
Таким образом, что мы получаем в очередной раз? Мы получаем (в очередной раз) факты, не укладывающиеся в теорию эволюции по дарвиновскому сценарию.
Зато эти же факты совершенно нормально укладываются в рамки теории непрерывного творения. Ведь биологические виды здесь являются продуктом дизайна, независимо от того, каким способом они размножаются (половым или бесполым). Причем этот дизайн достаточно надёжен, чтобы созданный таким образом биологический вид смог просуществовать потом 230 млн. лет без каких-либо изменений. Хотя, конечно, даже в рамках теории непрерывного творения столь потрясающее долгожительство биологического вида тоже достойно большого удивления (см. ниже).
ПРОБЛЕМА КРАСОТЫ
Еще одна группа фактов, которая не укладывается в современный дарвинизм, зато идеально укладывается в теорию непрерывного творения (и даже предсказывается этой теорией) - это так называемая проблема красоты.
В чем заключается эта проблема, наверное, ясно уже из названия. Все мы знаем, что очень многие живые существа на нашей планете имеют такие признаки, которые трудно объяснить острой необходимостью успешного выживания. Зато эти признаки нередко откровенно эстетичны. То есть, всё выглядит так, как будто целью существования таких признаков является не успешное выживание организма, а чисто эстетические мотивы.
Однако этого не может быть в рамках дарвинизма.
С позиций теории Дарвина, любой признак организма является продуктом естественного отбора (более приспособленных) в условиях борьбы за существование в ходе длительной эволюции жизни на Земле. Точнее, допускается возможность, что признак может быть и нейтральным, следовательно, может возникнуть под действием чисто случайных эволюционных факторов. Но только при условии, что этот признак не отнимает у организма слишком много ресурсов на свое создание и поддержание. Если же признак ярко выражен, то из самой логики теории следует, что этот признак должен иметь какое-то полезное, приспособительное значение. Иначе он был бы давно отсеян естественным отбором (хотя бы в целях экономии ресурсов).
Понятно, что если биологический признак ярко выражен, например, признак является громоздким, или очень сложным, то его появление у живых существ в результате чистой случайности – практически исключено. Над появлением этого признака должна была поработать главная творческая сила дарвиновской теории эволюции - естественный отбор более приспособленных.
Но естественный отбор может работать над созданием только таких признаков, которые полезны для выживания и размножения организмов. А над созданием признаков, бесполезных для выживания (нейтральных), и уж тем более, вредных для выживания – естественный отбор не может трудиться просто по определению (естественный отбор - это автоматический процесс преимущественного выживания более приспособленных).
Таким образом, в рамках теории естественного отбора, ярко выраженных признаков, не полезных для выживания или размножения – у организмов просто не может быть. Максимум, что следует ожидать - это наличие каких-нибудь рудиментов (ранее полезных признаков, а сейчас бесполезных), находящихся в стадии деградации.
Однако, несмотря на столь строгий запрет дарвиновской теории, очень многие живые существа имеют ярко выраженные признаки, которые, вроде бы, совершенно бесполезны для их прямого выживания (или даже вредны). То есть, многие живые существа (причем из самых разных биологических таксонов) имеют разнообразные «архитектурные излишества», которые нередко выглядят еще и эстетичными (хотя это и не обязательно).
Одним из самых одиозных примеров в этом отношении является, конечно, знаменитый хвост павлина, в котором роскошная расцветка сочетается со столь буйной гипертрофией самого «хвоста» (на самом деле, надхвостья), что это просто не может не доставлять павлину неудобств при выживании в дикой природе. И павлин поражает нас не только своим хвостом. Внешность павлина содержит целый ряд признаков, имеющих откровенно эстетическое значение. Начиная от знаменитых переливающихся глазков на хвосте, и кончая красивым венцом и «маской» на голове (Рис. 47):

Рисунок 4 7 . Самец обыкновенного павлина (Pavo cristatus).[117]
Создаётся ощущение, что весь облик павлина преследует цель максимизации именно эстетической составляющей - многие признаки согласованно вносят свой вклад в усиление общего эстетического эффекта. То есть, в случае самца павлина мы наблюдаем явную эстетическую целесообразность.
Эстетическую целесообразность можно рассматривать, как аналогию биологической целесообразности.
О явной биологической целесообразности живых существ я уже говорил выше. Биологическая целесообразность - это когда мы видим, что определенные признаки живого существа явно предназначены для какой-то определенной биологической цели. Например, никто не будет спорить, что цель наличия глаз (у того же павлина) – это обеспечение способности видеть. Крылья – предназначены для полета. Ноги – для передвижения по земле. И так далее. Биологическая целесообразность живых существ настолько очевидна, что не оспаривается ни кем.
Эта биологическая целесообразность сегодня может быть объяснена двумя принципиально разными способами:
1. Это продукт разумного дизайна.
2. Это результат длительной естественной эволюции в ходе естественного отбора более приспособленных.
Второй вариант объяснения в отношении глаз, крыльев и ног - теоретически вполне возможен. Так как глаза, крылья и ноги, действительно, полезны живому организму, повышая его приспособленность.
Далее по аналогии - мы ясно видим, что целью специфической внешности павлина является максимизация эстетической составляющей. То есть, мы явно наблюдаем эстетическую целесообразность.
Но в отличие от предыдущей (биологической) целесообразности, эстетическая целесообразность может быть объяснена только одним способом:
1. Это продукт эстетического дизайна.
Вторым способом (эволюцией более приспособленных) эта целесообразность объяснена быть не может, потому что чистая эстетика ничем не полезна её обладателю (а в случае павлина – даже вредна).
Причем мы все знаем, что откровенно эстетичная внешность павлина – далеко не единственный такой факт в живой природе, а скорее, лишь крошечная часть общего, огромного массива таких же фактов. Сюда в первую очередь относится яркая, красивая окраска многих живых существ (птиц, рыб, рептилий, насекомых, ракообразных, моллюсков и даже червей). А так же (помимо окраски) другие «архитектурные излишества», которые затрудняют жизнь своим обладателям.
Приходится признать, что проблема красоты родилась вместе с самой дарвиновской теорией. Ведь как только появилась идея, что живые организмы приобретают свои признаки благодаря естественному отбору более приспособленных, так и возникла необходимость объяснять, зачем же множеству живых существ такие признаки, которые неполезны для их выживания. Можно сказать, что дарвинизм родился на свет сразу с этой «родовой травмой» (проблемой красоты). И эта «родовая травма», похоже, неизлечима. Во всяком случае, сегодня факты складываются именно так.
На мой взгляд, эта группа фактов имеет очень большое значение для теоретической биологии. Потому что такие факты не только опровергают дарвиновские представления о развитии живой природы, но еще и (одновременно) свидетельствуют в пользу существования интеллектуальной силы, которая была «повинна» в создании разнообразия живых существ. Действительно, нам (людям) чисто эстетические мотивы (при создании тех или иных вещей) настолько знакомы и понятны, что обнаружение таких же фактов в живой природе (эстетических мотивов) почти единодушно воспринимается нами, как свидетельство в пользу разумного дизайна. Более того, позволяет судить о тех целях, которые преследовала разумная сила при создании живых существ. Вне всякого сомнения, чисто эстетические мотивы при создании разных групп живых существ прослеживаются достаточно часто. А в определенных биологических таксонах – очень часто.
В связи с чрезвычайной важностью (на мой взгляд) этой группы фактов для понимания реальных принципов, по которым создавалась живая природа, я сейчас не буду останавливаться на проблеме красоты подробно. Я посвящу этому вопросу свою следующую книгу. Ведь (еще раз) факты этой категории не только опровергают дарвинизм, но одновременно свидетельствуют в пользу разумного дизайна. А мы сейчас, в основном, занимаемся тем, что критически разбираем «доказательства эволюции», которые насобирали в свою копилку дарвинисты.
Поэтому здесь мы ограничимся лишь тем обстоятельством, которое уже отметили выше – многочисленные факты наблюдаемых «архитектурных излишеств» в живой природе – превосходно укладываются в рамки концепции разумного дизайна (в любой из его вариантов). И одновременно со страшным «скрипом» сопротивляются дарвиновской концепции. Верующие дарвинисты упорно пытаются всё-таки впихнуть эти факты (явной эстетики в живой природе) в свой несчастный «чемодан» дарвинизма, который просто не в состоянии их принять, а факты всеми силами упираются, возмущаясь подобным насилием.
Если коротко, то дела на этом «фронте» сегодня обстоят так:
1. Как правило, верующие дарвинисты пытаются запихнуть все те случаи, когда живые организмы (явно) эстетичны, в свой пресловутый «половой отбор». То есть, предлагается считать, что это просто самцы так перед самками стараются (отбираются), поскольку самки предпочитают именно эстетичных самцов.
А вот почему самки предпочитают именно самых эстетичных самцов (а не, допустим, самых энергичных) – над этим дарвинисты обещают подумать потом («требуются будущие исследования»).
То, что дарвинисты стараются избежать ответа на этот вопрос, вовсе не удивляет. Потому что здесь имеется логическое противоречие. В рамках самого дарвинизма, те самки, которым нравятся биологически бесполезные (и уж тем более, вредные) черты самцов - должны проигрывать (в размножении собственных генов) тем самкам, которые предпочитают самцов с биологически полезными признаками. По сути, самка, предпочитающая самца с вредными признаками – является просто носителем вредной мутации, и должна быть уничтожена естественным отбором. Это логическое противоречие дарвинисты пытаются преодолеть с помощью специальных (чисто теоретических) моделей полового отбора: модель «убегания» Фишера, и модель гандикапа.[118] Однако эти модели, в свою очередь, не могут ответить на первоначальный вопрос – почему же конечный результат этого (воображаемого) отбора самцов под давлением (воображаемых) предпочтений самок - столь часто получается именно красивым. А не, допустим, безобразным. Или просто эстетически нейтральным.
Пожалуй, приведу характерную цитату из соответствующего обзора, посвященного «успехам» теории полового отбора (Jones & Ratterman, 2009):
« Почему украшения, используемые птицами и другими животными, как правило, кажутся столь поразительно красивыми людям - это уже другой вопрос, но это тайна, которую не обязательно решать для понимания полового отбора».
То есть, как я и говорю - что такое половой отбор - понять можем. А вот решить, собственно, проблему красоты – не можем. Это пока «тайна», видите ли. Но если это «тайна», тогда зачем нам вообще теоретические модели полового отбора, если они как раз и создавались (изначально) для того, чтобы объяснить именно «тайну» наличия красоты в живой природе.
В общем, дарвиновские объяснения проблемы красоты получаются, во-первых, слишком громоздкими. Где одна теоретическая модель полового отбора взгромождается на другую модель полового отбора, при этом погоняя третьей моделью полового отбора.[119]
А во-вторых, в конечном итоге, все эти теоретические нагромождения (бесценных дарвиновских идей) всё равно не дают ответа на исходный вопрос – почему обсуждаемые признаки живых существ так часто кажутся эстетически привлекательными? Тем не менее, дарвинисты продолжают упорно цепляться за гипотезу полового отбора. Потому что цепляться больше не за что.
2. Для подтверждения своей гипотезы про «половой отбор», верующие дарвинисты проводят полевые исследования, где пытаются «поймать» соответствующие (эстетические) предпочтения самок. И естественно, много исследований (особенно выполненных ближе к концу 20 века) действительно находили то, что хотелось найти.
3. Гораздо хуже, когда такие (положительные) результаты, полученные в более ранних исследованиях, потом не воспроизводятся в более поздних независимых исследованиях. Что наводит на грустные мысли о том, что «ученые тоже люди» и что подгонку результатов (под нужную теорию) пока еще никто не отменял.
Дело дошло до того, что несмотря на огромное число проведенных исследований, сегодня всё еще нельзя назвать ни одного биологического вида, в отношении которого можно было бы уверенно сказать (на основании нескольких независимых исследований с согласующимися результатами), что данный вид подтверждает теорию полового отбора (Roughgarden & Akcay, 2010).
4. Еще хуже дело становится тогда, когда самец какого-либо биологического вида имеет явно вредный признак, но тем не менее, какую-либо роль этого признака в пресловутом половом отборе так и не удаётся показать. Не удаётся показать сразу в нескольких независимых исследованиях. В случае некоторых животных, даже множество проведенных исследований всё равно оказались не в состоянии найти ожидаемую связь между эстетичными признаками самцов и их успехом в размножении.
Именно такая скандальная ситуация сложилась сейчас, например, с птицами отряда куриных. В том числе, и со знаменитыми павлинами. Это, конечно, вообще анекдот - когда «икона полового отбора» оказалась не в состоянии подтвердить теорию полового отбора (сразу в трех последних независимых исследованиях). Но как я уже сказал выше, подробный анализ этих исследований - тема для следующей книги.
5. Еще хуже дела становятся, когда «архитектурные излишества» демонстрирует не только самец, но и самка тоже. То есть, сразу оба пола. Такие вещи теория полового отбора объяснить вообще не может. Конечно, выдвигаются определенные предположения, но настолько слабые, что их даже лень критически разбирать.[120]
6. И наконец, совсем плохо становятся дела, когда живое существо демонстрирует явно «радикальную» внешность, совершенно не полезную для выживания… но при этом имеет такие органы чувств, что просто не в состоянии оценить радикальные особенности собственной внешности.
Может быть, Вы помните, как мы разбирали выше «проблему белых бабочек». Для тех, кто уже не помнит, вкратце повторю, о чем речь. На рисунке 48 слева показана ночная бабочка березовая пяденица, которая имеет прекрасную маскирующую окраску, помогающую ей избегать внимания дневных насекомоядных птиц.
А справа мы видим тоже ночную бабочку, и тоже пяденицу. Но эта бабочка хорошо заметна на том фоне, где она «спряталась». Одновременно её внешность очень удачно совпадает с нашими эстетическими предпочтениями. Неслучайно название этой бабочки - пяденица великолепная (Iotaphora admirabilis).
Причем мы видим в её внешности много очень тонких деталей (обратите внимание на «проработку» цветных линий на крыльях), которые сама эта бабочка не в состоянии оценить чисто физически (Рис. 48):

Рисунок 48. Слева березовая пяденица, справа пяденица великолепная. Пояснения в тексте.
Если мы предполагаем, что окраску березовой пяденицы под фон дерева так хорошо подобрал естественный отбор (под давлением хищных птиц), тогда какая сила подбирала наряд пяденицы великолепной?
В следующей книге мы обязательно вернемся к этой теме еще раз. И познакомимся еще с парочкой «открытий чудных» в этой области. Здесь же я приведу только три конкретных примера, которые одновременно соответствуют озвученным выше пунктам 5 и 6 (Рис. 49):

Рисунок 49. Eudaemonia troglophylla . Бабочка из трибы Urotini, подсемейство Saturniinae.[121]
Обладательницей этих поразительно длинных «хвостов» на крыльях является бабочка Eudaemonia troglophylla (подсемейство Saturniinae). Одновременно, сотни видов бабочек из этого же подсемейства вообще не имеют ничего похожего на такие «хвосты». В качестве всего пары наглядных иллюстраций – например, бабочки Antherina suraka и Maltagorea fusicolor, относящиеся к этой же трибе бабочек, не имеют даже намека на какие-либо «хвостики» (Рис. 50):

Рисунок 50: Antherina suraka ( самец ) и Maltagorea fusicolor. Бабочки из трибы Urotini, подсемейство Saturniinae.[122]
Тем не менее, эти бабочки, вроде бы, не собираются вымирать из-за полного отсутствия каких-либо хвостов на крыльях (как и еще сотни видов бабочек из этого же подсемейства). Следовательно, в данном случае мы вряд ли можем сказать, что гигантские «хвосты» на крыльях бабочки Eudaemonia troglophylla - имеют страшную биологическую важность (для этого вида бабочки). Скорее уж, наоборот, здесь напрашивается вывод о несомненной биологической помехе. Это ведь надо, чтобы так угораздило – сначала отращивать, а затем еще и всё время таскать с собой такой вот дополнительный вес (и объем).
Похоже, это как раз тот случай, как нам надо «доставать из широких штанин» (С) именно версию полового отбора. Иначе объяснить столь радикальные особенности дизайна этого вида бабочки – вряд ли удастся. Но с позиций теории полового отбора, для того чтобы отобрать «хвосты» такой длины (и потом стабильно поддерживать этот признак из поколения в поколение), самки этого вида бабочек должны упрямо предпочитать самцов с более длинными «хвостами», жестоко отказывая самцам с более короткими хвостами. Но как же самки сделают это, если для этого нужно видеть разницу между «хвостами» разных самцов? Если самки не будут видеть тонких различий между длиной «хвостов» у разных самцов, тогда ничего не мешает этим «хвостам» постепенно уменьшиться (до биологически оптимальных размеров) незаметно для самок.
Я уже писал выше, что зрение бабочек не настолько хорошее, чтобы визуально оценивать подробные детали своего возможного полового партнера. Например, длину его «хвостов». Уже на расстоянии в два десятка сантиметров они этого сделать не смогут. А на очень близкой дистанции (в несколько сантиметров) такое «чудо природы», наверное, невозможно окинуть оценивающим взором уже чисто «технически».
Так какая же сила отбирала столь длинные «хвосты» бабочке Eudaemonia troglophylla, при этом не отобрав совершенно никаких хвостов бабочке Maltagorea fusicolor, хотя эти виды бабочек принадлежат к одной и той же трибе?
Причем у многих «длиннохвостых» бабочек хвосты имеют и самец, и самка. То есть, сразу оба пола. Поэтому о половом отборе здесь речи, похоже, вообще нет. А речь здесь идет (давайте честно признаемся) об определенном стиле дизайна, в соответствии с которым был создан данный вид бабочек. И этот стиль дизайна выдерживается и в отношении самца, и в отношении самки. Правда, в отношении самки этот стиль дизайна всё-таки выглядит более умеренным – как правило, у самок подобных бабочек «хвосты» заметно короче, чем у самцов. Оно и понятно – на самку ложится более серьезная нагрузка по жизни – ей надо создать много питательных яиц. Поэтому слишком увлекаться «архитектурными излишествами» самкам нельзя.
И это правило широко распространено в природе. Там, где мы видим у животных «архитектурные излишества», самки носят такие излишества в более умеренном варианте (если вообще носят). Помните, выше мы разбирали жирафов? Там самки тоже имели более «здравые» размеры тела, чем самцы. И соответственно, выживали, действительно, лучше (см. выше). То есть, здесь всё продумано – стили дизайна разных живых существ могут быть поразительными и даже радикальными, но в таком случае самки демонстрируют эти стили дизайна в «щадящем режиме». А у многих видов «архитектурные излишества» демонстрируют вообще только самцы. В рамках концепции разумного дизайна, здесь действительно всё разумно. А именно, и одна из основных целей дизайна достигается – создание максимального разнообразия форм жизни на Земле. И в то же время, конкретные биологические виды не поставлены на грань вымирания из-за слишком накладных «архитектурных излишеств». Наиболее накладные «архитектурные излишества» вообще несут только самцы. Самки же полностью высвобождаются для успешного продолжения рода. Например, у тех же индийских павлинов самки в период высиживания яиц и выращивания птенцов гибнут от хищников гораздо чаще, чем самцы (Takahashi et al., 2008), несмотря на то, что вообще не имеют никаких «украшений». А имеют скромную окраску, выполняющую классические функции маскировки. Понятно, что тут уже не до «архитектурных излишеств».
Но наивные дарвинисты, не понимая истинных причин красивой окраски самца павлина, объявляют индийских павлинов… классическим примером полового отбора. Они заявляют, что павлин такой красивый потому, что это его так отобрала павлиниха (так эстетично). Здесь перепутано вообще всё, что только можно, и поставлено с ног на голову. Более того, в качестве гениев эстетического дизайна выступают… курицы.[123]
К счастью, последние полевые исследования успешно провалили теорию полового отбора именно в отношении павлинов. И теперь верующим дарвинистам стало совсем плохо. Теперь им приходится рассуждать о воображаемых прошлых половых предпочтениях самок павлинов, которые уже успели измениться (Takahashi et al., 2008). То есть, соответствующие предпочтения самок «уже улетели, но обещали вернуться» (С). Как в том мультфильме про неуловимого Карлсона. И поэтому никаких связей между размером «хвоста» павлина и успехом его размножения никак не удаётся обнаружить (Loyau et al., 2005; Takahashi et al., 2008; Dakin et Montgomerie, 2011). Эту поучительную историю мы тоже подробно разберем в следующей книге.
А пока второй пример «архитектурных излишеств» (Рис. 51):

Рисунок 51. Жук арлекин (Acrocinus longimanus). Вверху самка , внизу самец .[124]
Это знаменитый жук-арлекин (Acrocinus longimanus). Как видим, здесь всё повторяется снова. Опять «архитектурные излишества» - непропорционально длинные передние лапы, которыми этот жук, собственно, и выделяется среди всех своих коллег - сотен тысяч (!) других видов жуков. И опять видим, что и самка (вверху) «выполнена» в соответствующем стиле дизайна, схожим с дизайном самца. Но одновременно хорошо видно, что дизайн самки гораздо более скромный (более благоразумный), чем «буйно-фантастический» дизайн самца. И снова не очень понятно, для чего же это «архитектурное излишество» вообще нужно.
Была идея, что такие длинные лапы нужны самцу этого вида для того, чтобы бороться за самку (и чтобы охранять те места, где самка откладывает яйца). Поскольку самцы этих жуков борются между собой, в том числе, и с помощью передних лап. Кроме того, самцы еще и кусают друг друга, и бодаются головами (Zeh et al., 1992). Однако проведенные исследования показали, что успех в этой борьбе способствует тем самцам, которые просто имеют большие размеры тела (Zeh et al., 1992). Какую-то особую роль именно длинных передних лап в поединках самцов выявить не удалось. В связи с этим, авторы исследования написали, что «фактор длины ног» маскируется корреляцией с общим размером тела (Zeh et al., 1992). Действительно, если большие размеры тела благоприятствуют победе, а длина передних ног, в свою очередь, коррелирует с размером тела, то длина передних ног тоже будет коррелировать с успехом. Вот только здесь получается такая «связь», которая аналогична «успеху боксеров в красных трусах» – если красные трусы надеть на боксеров весом 100 кг, а синие трусы – на боксеров весом 50 кг, то в поединках между этими боксерами чаще станут побеждать боксеры в красных трусах. Следовательно, именно красные трусы помогают побеждать в боксерских матчах?
Ну а если серьезно, то понятно, что бороться с другими самцами, имея такие «ходули» - вряд ли более удобно, чем имея «среднестатистические» ноги, как у сотен тысяч других видов жуков (в том числе, и у тех, кто любит бороться между собой). Здесь преимущество даёт общий размер тела, а не длина передних ног, тем более, такая «запредельная». В конце концов, если уж передние лапы так важны этим жукам для борьбы друг с другом, то в ходе эволюции следовало бы ожидать появления чего-то похожего на передние лапы богомола. Вот передние лапы богомола – это, действительно, крайне эффективное оружие. Но никак не те «ходули», которые имеются у жуков-арлекинов. Похоже, столь удлиненные передние лапы жукам-арлекинам вообще не нужны (ни для чего биологически полезного). Иначе из почти 400 тысяч (!) видов жуков мы непременно нашли бы еще тысяч десять аналогичных примеров. Вместо этого имеется лишь несколько видов жуков с длиннющими передними лапами на всё огромное «жучиное царство».[125]
В общем, самая правдоподобная и простая версия в данном случае напрашивается сама собой - у жуков-арлекинов просто такой уникальный стиль дизайна.
И уникальных стилей дизайна в природе великое множество. Одни только «стебельчатоглазые» мухи чего стоят (Рис 52):

Рисунок 52. Разные представители «стебельчатоглазых» мух. Да-да, не удивляйтесь, глаза всех этих мух находятся именно на концах длинных стебельков. Хотя в случае верхней мухи слева (Plagiocephalus latifrons) это уже больше похоже на огромные рога.
Остаётся только удивляться, как эти мухи всё еще умудряются летать.
Причем этот уникальный стиль дизайна характерен для многих видов мух сразу из нескольких семейств. А вот степень выраженности этого стиля различается даже среди близких биологических видов (Рис. 53):
Рисунок 53. 1,2,3 – самцы разных видов рода Plagiocephalus: 1 - P. latifrons; 2 - P. lobularis; 3 - P. intermedius. 4,5 – головы самок: 4 - P . latifrons ; 5 — P . intermedius . Изображения взяты из работы (Kameneva, 2004).
То есть, мы можем наблюдать в природе такие радикальные стили дизайна, в которых «выполнены» целые таксономические группы живых существ (вплоть до уровня семейств). Несмотря на отсутствие биологической необходимости в соблюдении подобного стиля. А внутри этого общего стиля дизайна, в свою очередь, наблюдается разнообразие его вариантов.
Кстати, в приведенном случае самки вообще не отягощены соответствующими «архитектурными излишествами» (см. рис. 53, изображения 4,5).
В тех же случаях, когда самки разделяют сходный стиль дизайна вместе со своими самцами, это обычно опять происходит в более «скромном» (биологически более благоразумном) варианте, чем у самцов (Рис. 54):

Рисунок 54. Спаривающиеся стебельчатоглазые мухи. Хорошо видно, что у данного биологического вида стебельчатоглазых мух: 1) самка имеет стиль дизайна, сходный со стилем дизайна самца, 2) но в более «скромном» (биологически более благоразумном) варианте.
Впрочем, среди стебельчатоглазых мух есть и такие виды, у которых и самцы и самки имеют «архитектурные излишества», развитые в совершенно одинаковой степени! Однако у таких видов, в целом, гипертрофия обычно выражена скромнее, чем у тех видов, где самцы по этому признаку отличаются от самок (Рис. 55):
Рисунок 5 5 . Разные виды и роды стебельчатоглазых мух (с предполагаемыми филогенетическими связями). Слева силуэт головы самцов разных видов, справа – самок. Рисунок взят из работы (Husak et al., 2011).
Из рисунка хорошо видно, что стебельчатоглазые мухи представлены как мономорфными видами, то есть такими, где морфология самца и самки вообще не различается. Так и диморфными видами, у которых имеется половой диморфизм по обсуждаемому признаку. Заметно, что в случае мономорфных видов, гипертрофия «стебельков», в целом, выражена слабее, чем в случае диморфных видов. В свою очередь, у самок диморфных видов гипертрофия признака всегда выражена скромнее, чем у самца.
И вот нам предлагается поверить, что этот радикальный стиль дизайна, в котором «выполнены» целые биологические роды в разных семействах мух (данный стиль дизайна повторяется еще и в разных семействах)… так получилось исключительно потому, что самки мух упорно предпочитали самцов с самыми широко расставленными глазами. И в результате этих, поразительно устойчивых половых предпочтений (не изменявшихся на протяжении долгих миллионов лет!) образовались целые роды мух с видами, поголовно обладающими именно данным стилем внешности (в той или иной степени). Получается, что в рамках сразу нескольких биологических родов, имела место упорная «эволюционная движуха» в строго определенном направлении, да еще и вопреки давлению обычного естественного отбора. Ведь никто, надеюсь, не будет спорить, что мухам со «среднестатистической» головой выживать всё-таки легче, чем с такой головой, которую имеют стебельчатоглазые мухи.
Интересно, что имеется ряд публикаций, в которых версия полового отбора у стебельчато-глазых мух… получила подтверждение. Будто бы самки стебельчатоглазых мух, действительно, предпочитают самцов с самыми широко расставленными глазами. И самцы с такими глазами, будто бы, тоже имеют преимущество в поединках с другими самцами.[126]
Такое явление я называю «телекинезом в биологии». Когда очень хочется, чтобы результаты твоих собственных исследований сошлись с прогнозами господствующей теории, то (о чудо!) результаты действительно могут сойтись с тем, с чем тебе очень хотелось, несмотря на реальное положение дел в данной области. Видимо, сила мысли некоторых исследователей является настолько мощной, что сама реальность начинает искажаться, подстраиваясь под теоретические ожидания этих ученых. Такое явление я и называю «телекинезом в биологии». Особенно легко «телекинез в биологии» может происходить там, где исследования связаны с выявлением каких-нибудь статистических закономерностей, то есть там, где несколько лишних точек на графике способны изменить результат работы с формулировки «ожидаемой связи не выявлено» до формулировки «ожидаемая связь установлена».
К сожалению, у научных результатов, добытых таким «телекинетическим способом», обычно начинаются проблемы с воспроизводимостью, как только другие исследователи начинают проводить независимые исследования. Видимо, другие исследователи уже не обладают достаточной силой мысли, чтобы влиять на реальность правильным образом. Поэтому они и не могут воспроизвести результаты, добытые с помощью «телекинеза в биологии». Таким образом, «телекинетические» результаты обычно живут в науке только до того момента, пока не будут проведены другие (независимые) исследования в этой же области.
К сожалению, независимых специалистов часто не хватает для того, чтобы проводить повторные исследования одних и тех же биологических объектов. Поэтому иногда «телекинетические» результаты в биологии могут спокойно «жить» годами. И даже попадать в соответствующие учебники в качестве «доказанных фактов». В итоге складывается ситуация, когда одни «телекинетические» результаты опровергаются независимыми исследованиями. А другие всё еще «живут», дожидаясь своего опровержения. Например, «обнаруженные» в ранних исследованиях половые предпочтения пав (самок павлинов) к самым большим хвостам своих кавалеров – уже опровергнуты в более поздних работах. А вот «половые предпочтения стебельчатоглазых мух», вроде бы, пока еще только дожидаются своего «разрушителя мифов».
Впрочем, некоторые авторы, стараясь подтвердить половой отбор слишком рьяно, иногда неправильно понимают «линию партии»… и в результате получают такие результаты, которые, наоборот, опровергают версию про половой отбор. Например, в работе (Wilkinson & Reillo, 1994) авторы установили, что если начать искусственно отбирать линию мух с коротким расстоянием между глазами, то уже через 13 поколений такой селекции, самки этой линии начинают предпочитать самцов с коротким расстоянием между глазами! То есть получается, что самки всего за 13 поколений самостоятельно изменили свои исходные половые предпочтения не просто на другие, а вообще на противоположные.
По сути, получилось, что половые предпочтения самок стебельчатоглазых мух просто пассивно следуют за тем направлением отбора, которое имеется в данный момент, не демонстрируя никакой устойчивости. Понятно, что такой результат сразу же обесценивает версию о половом отборе, который якобы упорно происходил в строго определенном направлении в рамках целого биологического семейства, на протяжении долгих миллионов лет. Ведь если половые предпочтения самок настолько неустойчивы, что могут изменяться (вообще на противоположные) за 13 поколений, то ни о каком формировании (под действием столь изменчивых половых предпочтений) единого морфологического облика сразу у множества биологических видов (относящихся к целому семейству мух) – не может быть и речи.
В заключение давайте еще раз подчеркнем общую иронию ситуации в случае со стебельчатоглазыми мухами:
1. Стебельчатоглазые мухи имеют явно радикальную внешность.
2. В рамках дарвинизма, эта внешность объясняется тем, что, во-первых, самки этих мух предпочитают самцов с самыми широко расставленными глазами. А во-вторых, самцы с широко расставленными глазами имеют преимущество в поединках с другими самцами. Таким образом, обсуждаемый признак связан исключительно с отбором самцов.
3. Тем не менее, самки стебельчатоглазых мух этого рода – тоже стебельчатоглазы.
4. Более того, чуть ли не у половины представленных видов, самки «стебельчатоглазы» совершенно в той же степени, что и самцы (см. рис. 55).
5. Причем этот поразительный стиль дизайна наблюдается практически поголовно в рамках целого биологического семейства (и не только).
Так какое же объяснение причин подобной внешности (у целых биологических семейств, у самцов и самок (!) тех видов, которые принадлежат к этой группе) звучит более правдоподобно? Что это результат полового отбора самцов? Или, что это такой уникальный стиль дизайна, характерный для соответствующих биологических таксонов?
Итак (заканчиваем), теория разумного дизайна объясняет факты существования подобных «архитектурных излишеств» в живой природе, в целом, более правдоподобно и намного проще, чем дарвиновская теория. А именно, разнообразием стилей дизайна живых существ. Где создаваемое разнообразие, скорее всего, не преследовало никаких других целей, кроме, собственно, самого биологического разнообразия.
Ну а подробно на эту тему мы еще поговорим позже, в следующей книге.
Дата: 2019-02-02, просмотров: 688.