Еще одна группа фактов, которая хорошо укладывается в теорию непрерывного творения (впрочем, как и в любой другой вариант концепции разумного замысла), но одновременно совершенно не укладывается в рамки дарвиновской теории – это результаты современного моделирования генетических процессов в популяциях (Sanford et al., 2007a,b). Эти результаты показывают, что биологические виды должны постепенно генетически деградировать. То есть, генофонд любого биологического вида должен постепенно ухудшаться со временем. Это будет происходить за счет естественных процессов постепенного разрушения геномов, где в качестве разрушающего фактора выступает всё тот же поток случайных мутаций. Как известно, большинство случайных мутаций являются вредными или слабо-вредными. Еще какая-то часть мутаций являются нейтральными (или «условно нейтральными»). А вот полезные мутации гораздо более редки. Таким образом, генофонд реальных популяций в природе постоянно подвергается, в целом, разрушающему «дождю» из случайных мутаций. И в этом «дожде» особенно опасны слабо-вредные мутации. Ведь естественный отбор реагирует на них слабо, потому что они оказывают лишь слабый негативный эффект на жизнедеятельность конкретной особи. Поэтому слабо-вредные мутации не успевают удаляться из популяций очищающим отбором, и постепенно накапливаются в этих популяциях, приводя к медленному снижению их приспособленности. В конце концов (теоретически) эти разрушительные процессы должны приводить к полной деградации приспособленности популяций и их вымиранию.
Чтобы ничего особенного от себя не выдумывать, приведу несколько оригинальных комментариев на эту тему автора соответствующей книги («Генетическая энтропия и тайна генома») генетика Джона Сэнфорда: [127]
«Мутации представляют собой ошибки текстовой обработки в инструкции о порядке работы клетки. Мутации постоянно разрушают генетическую информацию так же, как ошибки текстовой обработки разрушают записанную информацию. Несмотря на то, что иногда происходят полезные мутации (так же, как иногда бывают полезными орфографические ошибки), вредные мутации намного превосходят их по числу. На одну полезную мутацию приходится около миллиона вредных мутаций. Поэтому, даже если учитывать полезные мутации, конечный их результат очень вреден. Чем больше мутаций, тем меньше информации – в этом заключается суть мутационного процесса».
«Естественный отбор ничем не может помочь. Отбор лишь помогает избавиться от наиболее вредных мутаций. Он замедляет мутационное вырождение.
Кроме того, очень редко возникает полезная мутация, имеющая достаточно влияния, чтобы быть отобранной. Это приводит к адаптивной радиации или некой корректировке, что также способствует замедлению вырождения. Но отбор удаляет очень небольшую часть вредных мутаций. Подавляющее большинство вредных мутаций неумолимо накапливаются и являются слишком коварными (имеют слишком маленькое влияние), чтобы существенно влиять на свою жизнестойкость. С другой стороны, практически все полезные мутации (в случае, если они происходят) невосприимчивы к процессу отбора, так как они вызывают минимальные увеличения биологической функциональности.
Таким образом, несмотря на интенсивный отбор, большинство полезных мутаций «дрейфуют» (выходят) из популяции и теряются…
…Отбор замедляет мутационное вырождение, но фактически он нисколько его не останавливает. Так что даже при интенсивном отборе эволюция идет в ложном направлении – в направлении вымирания!».
И о самой книге «Генетическая энтропия и тайна генома»:
«Недавно опубликованная мной книга – плод многолетней научной работы. В ней рассказано, как я полностью переоценил все, что, как я думал, мне было известно об эволюционной генетической теории. В книге исследуются проблемы, лежащие в основе классической теории неодарвинизма. Моя цель заключалась в том, чтобы показать проигрыш дарвиновской теории на каждом уровне. Она не эффективна, потому что:
- мутации появляются быстрее, чем отбор может их вывести из генома;
- мутации слишком коварны, чтобы их можно было отобрать;
- «биологический шум» и «выживание счастливчиков» подавляют отбор;
- вредные мутации физически связаны с полезными мутациями, так что при унаследовании их невозможно разделить (т.е. избавиться от вредных мутаций и оставить полезные)»
Итак, мы видим, что все рассуждения здесь идут в рамках обычной популяционной генетики. Однако роль мутационного процесса и естественного отбора кардинально переоценена.
В современной (дарвиновской) теории эволюции естественный отбор не только успешно побеждает разрушающий эффект потока вредных мутаций, но еще и успевает при этом замечать и закреплять изредка появляющиеся полезные мутации. И таким образом, приспособленность популяций постепенно повышается.
А вот у авторов соответствующей компьютерной программы, моделирующей происходящие генетические процессы в популяциях, получилось наоборот (Sanford et al., 2007a,b). Естественный отбор не только не успевает повышать приспособленность популяции, но даже оказывается не в состоянии просто поддерживать качество генофонда популяции на постоянном, приемлемом уровне. Постепенно популяции генетически деградируют, накапливая всё больше и больше слабо-вредных мутаций. Вплоть до того, что генофонд совсем «дряхлеет», и за этим следует вымирание.
Понятно, что если авторы всё правильно смоделировали, то это ставит однозначный крест на дарвинизме. Сам автор книги (он же один из авторов соответствующей компьютерной программы) вообще пришел к выводу, что возраст всей жизни на Земле не может быть больше 100 тысяч лет. Иначе бы всё живое давным-давно уже повырождалось и исчезло с лица Земли. То есть, автор стал придерживаться младоземельных взглядов на развитие жизни.
Впрочем, в теорию непрерывного творения (с большими возрастами Земли) подобные результаты моделирования генетических процессов в популяциях тоже вполне укладываются. Более того, может быть, становится даже понятно, зачем требовалось создавать всё новые и новые биологические таксоны. Возможно, именно затем, что «старые» таксоны постепенно деградировали и вымирали (вследствие только что описанной генетической энтропии). И периодическое «обновление» биологического разнообразия Земли было просто необходимой мерой.
Неуменьшаемая сложность. Эту термин появился с легкой руки Майкла Бихи (Behe, 1996). Сегодня мы знаем, что устройство многих биологических признаков (у самых разных живых существ) может иметь просто потрясающую сложность.
Давайте, например, еще раз полюбуемся, сколько нужно взаимодействующих генов, чтобы мы с Вами имели возможность говорить (Рис. 56):
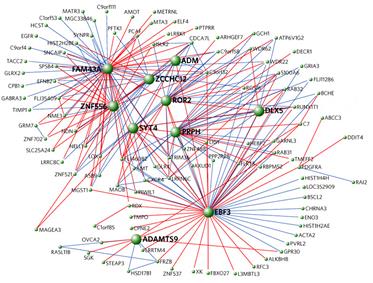
Рисунок 5 6 . Комплекс связанных генов, так или иначе взаимодействующих с геном FOXP 2, одним из генов, ответственных за формирование речи у человека ( Konopka et al., 2009).
Число взаимодействующих генов здесь весьма впечатляет. Очень трудно представить себе естественную (постепенную) «самосборку» признака такой степени сложности - за счет того, что включение каждого нового гена в этот комплекс обязательно приносило организму какую-то гипотетическую пользу.
Это всё равно, как если бы мы искренне верили, что имеется реальная возможность создать турбореактивный двигатель, просто добавляя к этому двигателю по одной детали поочередно, причем таким образом, что этот турбореактивный двигатель каждый раз (после добавления каждой новой детали) становился бы для нас все полезнее и полезнее.
На самом деле, мы понимаем, что такого способа сборки реактивного двигателя не существует. Наоборот, подавляющее большинство деталей нам придётся соединять друг с другом без какой-либо немедленной выгоды. Польза от всех этих действий появится только тогда, когда турбореактивный двигатель будет собран в достаточной степени, чтобы начать хотя бы минимальную работу. А до этого момента (неработающий) реактивный двигатель – не более чем груда бесполезного металла. Причем груда весьма обременительная.
Понятно, что какие-то (дополнительные) детали с турбореактивного двигателя всё-таки можно снять, чтобы этот двигатель продолжал еще хоть как-то работать. Но столь же очевидно, что должен существовать и целый ряд совершенно необходимых деталей, без которых даже минимальная работа этого двигателя будет уже невозможной.
Это и есть знаменитая «проблема неуменьшаемой сложности» - для того, чтобы сложная система оказалась полезной (хотя бы в минимальной степени), для этого она должна быть уже рабочей (хотя бы в минимальной степени). Но для того, чтобы эта сложная система работала – необходимо одновременное наличие (и правильное взаимодействие друг с другом) хотя бы нескольких элементов, составляющих эту систему. Однако по отдельности, эти (разные) элементы – вообще бесполезны. Или даже обременительны (для организма). Поэтому они не могут быть отобраны естественным отбором. Каким же тогда образом эти элементы смогли появиться и закрепиться в популяциях (вопреки естественному отбору), чтобы потом (случайно) образовать сложную систему?
Пожалуй, еще раз приведу красноречивую цитату из работы (Анохин и др., 2007), посвященную именно этой проблеме:
Одной из наиболее острых проблем, стоящих перед современной эволюционной теорией, является вопрос об эволюционном возникновении новых форм поведения. Нейробиологические исследования последних лет показывают, что в основе эволюционно сформированного поведения лежит координированная активность миллионов нервных клеток, образование специфических связей между которыми требует участия сотен генов. С одной стороны, для возникновения такой новой интеграции кажется очевидной необходимость одновременного появления многих комплементарных генетических изменений, в противном случае функция не будет обеспечена и организм не получит адаптивного преимущества в естественном отборе. С другой стороны, вероятность синхронного совпадения такого большого числа благоприятных мутаций чрезвычайно мала. Данное противоречие составляет одну из главных проблем в теории эволюции поведения. Еще более глубоким является вопрос о механизмах эволюционного возникновения сложных поведенческих паттернов. Каким образом в филогенезе выстраиваются сцепленные последовательности поведенческих актов, где животное должно выполнить серию действий, каждое из которых недостаточно для адаптации, но своим удачным завершением запускает активацию следующего поведенческого этапа, ведущего к окончательному приспособительному результату?
Для преодоления этой очевидной трудности, верующие дарвинисты используют ряд теоретических рассуждений. Во-первых, о том, что «пошаговое» совершенствование минимально работающих систем путем постепенного добавления к ним единичных полезных мутаций – всё-таки, возможно. А во-вторых, о том, что «минимально работающие системы» могли возникать, например, для решения других задач выживания, но потом могли изменять свои функции и служить основанием (преадаптацией) для создания уже других органов и тканей.
К сожалению, подобные рассуждения пока (вот уже сотню лет) не выходят за рамки чистых спекуляций, не подтвержденных ничем, кроме, собственно, теоретической возможности подобных сценариев. Причем гипотезы, предлагающие разные эволюционные схемы постепенного появления тех или иных сложных органов (или процессов) - практически всегда имеют самый общий, «расплывчатый» характер. Без детальной проработки подробностей этого процесса.
Такие туманные эволюционные сценарии, наверное, можно придумать для объяснения практически любого сложного органа. Достаточно как следует напрячь эволюционное воображение. Поэтому «проблема неуменьшаемой сложности», кажется, никогда не была особенно суровым испытанием для тех дарвинистов, которые обладают богатым воображением. Ведь «объяснить» чисто теоретически (исключительно на словах, без каких-либо практических подтверждений), да еще и задним числом – можно вообще всё что угодно.
Однако в любом случае, появление сложных органов по таким теоретическим сценариям должно было иметь чрезвычайно постепенный характер. Ведь к (воображаемо) формирующемуся сложному признаку каждый раз должна была добавляться единственная случайная мутация,[128] которая к тому же должна была оказаться непременно полезной (хотя бы немного улучшающей работу сложного признака в целом). Понятно, что такие события должны были происходить весьма редко.
И вот теперь получается, что такой очень постепенно формирующийся орган должен был формироваться еще и в условиях непрерывного «дождя» из слабовредных мутаций, которые всё время норовили (почти незаметно) снижать эффективность работы этого органа («генетическая энтропия»). Весьма трудно представить, что естественный отбор - настолько талантливый товарищ, что способен «вылавливать» из общего мутационного «ливня» редчайшие мутации, которые чуть-чуть повышали эффективность формирующегося сложного признака, одновременно выбраковывая многие тысячи мутаций, которые норовили чуть-чуть снизить эффективность этого же сложного признака (образованного, может быть, сотнями разных генов).
Но и это еще не всё. Имеется еще одна очень серьезная трудность современной теории эволюции, которая находится строго в рамках обычной популяционной генетики. Это знаменитый «кошмар Дженкина». К сожалению, в российских литературных источниках суть аргумента Дженкина была очень серьезно искажена. В результате сложилось впечатление, что аргумент Дженкина по мере развития генетики был снят. А именно, в многочисленных «вольных пересказах» всё выглядит так, будто Дженкин рассуждал о «разбавлении» простого признака (то есть, такого признака, который обусловлен работой только одного гена).
Если же обратиться к оригинальной работе Дженкина (Jenkin, 1867), то становится ясно, что Дженкин говорил о «разбавлении» сложного признака, то есть, образованного удачным сочетанием многих признаков. Насколько мне известно, в русскоязычной печати на это обстоятельство (искажение оригинального аргумента Дженкина) впервые указал Д.А. Шабанов (Шабанов, 2006; Шабанов, 2014).
Если же озвучить аргумент Дженкина в его правильном (исходном) смысле, тогда получится, что «кошмар Дженкина» не то что до сих пор не снят в рамках современной теории эволюции, а наоборот, является одним из критических мест этой теории.
Дело в том, что в популяциях организмов, размножающихся половым путем, в генотипе одной конкретной особи может образоваться удачное сочетание разных генов. И это сочетание разных генов (которые могут находиться даже в разных хромосомах) может оказаться биологически полезным. В то время как каждый отдельный ген (или аллель), входящий в это полезное сочетание – может вообще не иметь биологической пользы (вне этого сочетания). Допустим, такая (удачливая) особь, с удачным сочетанием генов (или аллелей), будет отобрана естественным отбором. Однако это удачное сочетание генов… с высокой вероятностью будет разбито в следующих поколениях - за счет банального расхождения гомологичных хромосом по разным половым клеткам. Кроме того, полезное сочетание аллелей может быть разбито даже в одной хромосоме (!) еще и рекомбинацией (кроссинговер). Получается, что в данном случае естественный отбор, по сути, сработал «вхолостую». Ведь он отобрал такой полезный (сложный) признак, который, по сути, не является наследуемым.
По этому поводу в русском языке даже имеется соответствующая поговорка:
«На детях гениев природа отдыхает» (С).
В качестве наглядного пояснения к этой поговорке могу привести еще более откровенную цитату (из работы Шабанов, 2014):
«…Стоит половому процессу выстроить удачную комбинацию генов, как он тут же рассыпает её. Создатели печально известной, хотя в основном безвредной организации, прозванной «Нобелевским банком спермы», упустили из виду именно это. Когда биохимику Джорджу Уолду предложили сдать свою заслуженную сперму в этот банк, он отказался, отметив, что просителям была бы нужна скорее не его сперма, а сперма таких людей, как его отец, бедный портной-иммигрант, чьи чресла, как ни странно, оказались источником гениальности. «А что дала миру моя сперма? — сокрушался нобелевский лауреат. — Двоих гитаристов!» (Лейн Ник. Лестница жизни. — М.: АСТ, 2013. — С. 187.
Да, ещё надо добавить, что охотникам за «генами успеха» на самом-то деле нужна не вся сперма иммигранта-портного, а один-единственный его сперматозоид, желательный для оплодотворения той самой яйцеклетки его жены, из которой развился его сын. Из остальных половых клеток этой пары могли получиться весьма разные по своим способностям люди. Это — аргумент Дженкина в чистом виде!»
И такие «холостые срабатывания» естественного отбора, кажется, должны происходить весьма часто – ведь многоклеточные организмы обладают массой сложных признаков.
Например, какая будет польза от сложнейшего комплекса из сотен генов, работа которых в совокупности формирует (весьма совершенный) человеческий глаз… если при этом в генотипе этой же особи не будет ряда других генов, которые формируют, допустим, глазной нерв, соединяющий человеческий глаз с человеческим мозгом? Ответ в данном случае очевиден – никакой пользы (не будет).
Поэтому закрепление в ходе эволюции сложных признаков (образуемых сочетанием разных генов) в половых популяциях – крайне проблематично («кошмар Дженкина»). Например, компьютерное моделирование показывает, что если полезно только удачное сочетание из четырех независимых признаков (но не каждый из этих признаков в отдельности), естественный отбор уже оказывается практически неспособным отобрать эти признаки. [129]
В результате, данное полезное сочетание (из четырех признаков) – не закрепляется в популяции. Наоборот, дрейф генов постепенно удаляет эти гены из популяции (по отдельности).
А если для того, чтобы стать полезным, требуется сочетание не четырех, а пяти, десяти, или, допустим, тридцати разных генов? Каким же тогда образом возникали такие сложные признаки в ходе эволюции?
Если учесть «кошмар Дженкина», то образование подобного комплекса эволюционным способом начинает казаться вообще невероятным – потому что получается, что этот сложнейший признак в ходе своего постепенного формирования – норовил еще и всё время «рассыпаться» в каждом новом поколении (за счет расхождения по разным половым клеткам, и еще за счет рекомбинации признаков). То есть, для того чтобы сформировать этот признак, естественному отбору необходимо было (миллионы лет?) следить не только (и даже не столько) за тем, чтобы обсуждаемый генетический комплекс постепенно (очень постепенно) становился всё более сложным - за счет добавления («методом случайного тыка») новых генов, делающих этот признак всё более совершенным (а эти новые гены, в свою очередь, теоретически появлялись за счет чисто случайных мутаций)… Но еще и за тем, чтобы этот сложнейший признак не рассыпался в каждом новом поколении в результате банальных процессов, происходящих с хромосомами любых живых существ, размножающихся половым путем.
Понятно, что в конечном итоге, разумная сборка таких генетических комплексов (в ходе разумного дизайна) выглядит гораздо более правдоподобно, чем предположение о постепенной естественной эволюции признаков подобной степени сложности методом редких, единичных «случайных тыков», в то время как эти признаки:
1) постоянно стремились рассыпаться в каждом новом поколении («кошмар Дженкина»)
2) норовили всё время (незаметно) деградировать из-за накопления слабо-вредных мутаций (генетическая энтропия).
Дата: 2019-02-02, просмотров: 733.