1. Энтероцельная гипотеза - целомы образуются в результате обособления "карманов" кишечника. (Вспомнить схему 8-лучевого кораллового полипа, гребневиков).
2. Гоноцельная гипотеза - целом от половых желез. Целом происходит от гонад низших червей. Полость каждого полусегмента аннелиды соответствует полости одного фолликула турбеллярии или немертины. Эпителий целома (перитонеум) соответствует стенке гонады, целомодукты - половым протокам. Наилучшей исходной формой для выведения метамерного целома аннелид являются с этой точки зрения немертины с их часто метамерно расположенными мешковидными гонадами, из которых каждая открывается наружу отдельным коротким протоком. Проток - целомодукт, стенка гонады - эпителий целома.
3. Миоцельная гипотеза - целом возник как полость в мускульной закладке животного, заполнившаяся жидкостью и играющая благодаря этому роль опорного образования для окружающей мускулатуры. (Метамерность, эпителиально-мышечные клетки)
12. Взгляды А. Ремане (архицеломатная гипотеза).
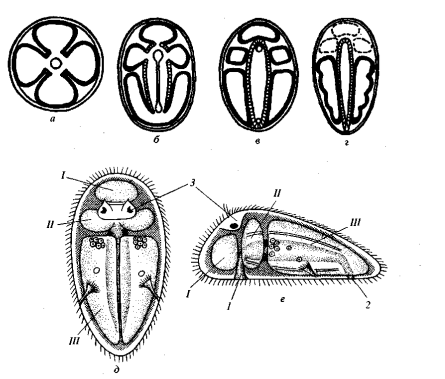
Рис. 2. Происхождение Bilateria согласно архицеломатным гипотезам (по: Remane et al., 1989): а - строение четырех-
лучевого кораллового полипа - предка Bilateria, б - вытягивание первичного рта и начало дифференцировки гаст-
ральных карманов, в - разделение первичного рта на рот и анус (амфистомия) и отделение гастральных карманов
(=целомических мешков) от кишечника, г - редукция протоцеля и мезоцеля Spiralia, д - строение гипотетического
архицеломатного предка, вид со спинной стороны, е - то же, вид слева. Обозначения: 1 - рот, 2 - анус, 3 - нервный
центр, / - предротовой целом (протоцель), // - околоротовой целом (мезоцель), III - туловищный целом (метацель).
Архицеломатные гипотезы. Сторонники этих гипотез (Masterman, 1897,1898; Remane, 1950,1958, 1967; Ulrich, 1950, 1951; Marcus, 1958; Siewing,1967, 1969, 1972, 1973, 1974, 1980, и др.) предполагали, что предками Bilateria были четырехлучевые полипы, гастральная полость которых была разделена перегородками на четыре камеры (рис. 2, а). Такие полипы стали ползать на ротовой поверхности, которая превратилась в брюшную сторону. Первичный рот вытянулся и стал щелевидным (как у современных коралловых полипов), а затем сомкнулся посредине так, что от него осталось только два отверстия: одно на переднем, а другое - на заднем конце тела (рис. 2, б, в). Переднее отверстие стало ртом, а заднее - анусом. Гастральные карманы отделились от центральной части кишечника и дали начало пяти целомическим камерам: непарной предротовой, парным околоротовым и парным туловищным (рис. 2, в). Предротовая и околоротовые камеры произошли непосредственно от карманов гастральной полости, а туловищная пара целомов возникла из заднего гастрального кармана, который оказался разделенным из-за того, что анальное отверстие сместилось на задний конец тела.
Таким путем возникло расчленение тела первичных Bilateria на три архимера. Целом тоже оказался подразделенным на три отдела: предротовой (протоцель), околоротовой (мезоцель) и туловищный (метацель или соматоцель) (рис. 2, д, е).
Среди современных организмов к таким формам ближе всего несегментированные целомические организмы (сипункулиды, плеченогие, форониды, полухордовые). Впоследствии у Spiralia предротовой и околоротовые целомы редуцировались, а туловищный (например, у аннелид) подвергся метамерии (рис. 2, г). У Deuterostomia предротовой целом расщепился на два диссиметричных мешка, а туловищный целом (только у хордовых) подвергся метамеризации (см. Siewing, 1969, 1973, 1974, 1980). По мнению сторонников архицеломатных гипотез, все нецеломические Bilateria утратили целом, а некоторые (например, плоские черви) - и сквозной кишечник.

Дата: 2019-02-02, просмотров: 872.