И Геккель и Мечников придерживались колониальной гипотезы. Они так же считали, что предками Metazoa были гетеротрофные жгутиконосцы.
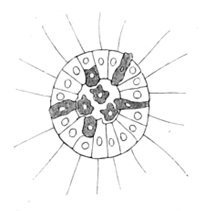 Гипотеза гастреи. Согласно этой гипотезе все многоклеточные животные произошли от одного общего предка — гипотетического организма — гастреи. По мнению Геккеля, гастрея имела овальное мешковидное тело с двухслойной стенкой и ротовым отверстием. Она образовалась из неспециализированной и неинтегрированной колонии (соответствует бластуле). Далее задний полюс впячивается (инвагинация), образуется гастрея. У неё уже есть кишка. Из гастреи Геккель выводил кишечнополостных. У теории гастреи немного сторонников, т. к. нет оснований считать инвагинацию первичным способом образования энтодермы.
Гипотеза гастреи. Согласно этой гипотезе все многоклеточные животные произошли от одного общего предка — гипотетического организма — гастреи. По мнению Геккеля, гастрея имела овальное мешковидное тело с двухслойной стенкой и ротовым отверстием. Она образовалась из неспециализированной и неинтегрированной колонии (соответствует бластуле). Далее задний полюс впячивается (инвагинация), образуется гастрея. У неё уже есть кишка. Из гастреи Геккель выводил кишечнополостных. У теории гастреи немного сторонников, т. к. нет оснований считать инвагинацию первичным способом образования энтодермы.
Гипотеза фагоцителлы. По Мечникову гаструляция происходит следующим образом: хоаноциты с большим количеством пищеварительных вакуолей убирают жгут, мигрируют в мезохилл. Там они претерпевают деление. Т.е. происходит мультиполярная миграция. Далее происходит клеточная специализация: двигательные клетки внешнего слоя теряют способность к миграции. На заднем (оральном) полюсе располагаются трофические клетки.
***
· Инвагинация — происходит путём втягивания стенки бластулы в бластоцель; характерна для большинства групп животных.
· Деламинация (характерна для кишечнополостных) — клетки, находящиеся снаружи, преобразуются в эпителиальный пласт эктодермы, а из оставшихся клеток формируется энтодерма. Обычно деляминация сопровождается делениями клеток бластулы, плоскость которых проходит «по касательной» к поверхности.
· Иммиграция — миграция отдельных клеток стенки бластулы внутрь бластоцеля.
· Униполярная — на одном участке стенки бластулы, обычно на вегетативном полюсе;
· Мультиполярная — на нескольких участках стенки бластулы.
· Эпиболия — обрастание одних клеток быстро делящимися другими клетками или обрастание клетками внутренней массы желтка (при неполном дроблении).
· Инволюция — вворачивание внутрь зародыша увеличивающегося в размерах наружного пласта клеток, который распространяется по внутренней поверхности остающихся снаружи клеток.
Целлюляристкие гипотезы.
Вообще-то предположение, что многоклеточные животные произошли не от колонии жгутиконосцев, а от одиночного простейшего путем одновременного обособления в последнем многих клеток (т. е. путем целлюляризации) впервые выдвинул Иеринг (Iering, 1877), и оно поддерживалось А. А. Тихомировым (1887) и Френцелем (Frencel, 1892) (цит. по: Иванов, 1968). Однако такого рода взгляды получили наибольшее развитие именно в работах Хаджи. Этот исследователь произвел тщательное сравнение многоядерных инфузорий и бескишечных турбеллярий (Acoela) и обнаружил очень много сходств в их строении; он принял, что сходные структуры одновременно являются также гомологичными. По мнению Хаджи, сократительным вакуолям инфузории соответствуют нефридии червя, трихоцистам — рабдиты. И у того и у другого организма при питании образуются пищеварительные вакуоли. Как инфузории, так и бескишечные черви имеют небольшие размеры; они плавают с помощью ресничек, всегда гермафродиты. Хаджи считал, что в процессе эволюции часть многочисленных ядер у предковой формы Metazoa стала располагаться над поверхностью тела, затем здесь произошло обособление цитоплазмы вокруг каждого ядра и образовались клетки. У Acoela процесс целлюляризации еще не зашел далеко, поэтому внутренний фагоцитобласт имеет неклеточное строение. В теле более высокоорганизованных плоских червей внутренняя цитоплазматическая масса также распалась на отдельные клетки, так что процесс целлюляризации завершился полностью. Следовательно, по мнению Хаджи, по своей природе инфузория вполне может быть сравнима с целым организмом низших Metazoa. Теорию целлюляризации поддерживают многие зарубежные ученые.
Наши отечественные исследователи относятся к этой теории отрицательно. Наиболее развернутую и обоснованную критику дает ей академик А. В. Иванов. Современные электронно-микроскопические исследования четко показывают, что тонкое строение покровов, фибриллярных систем, связывающих кинетосомы ресничек, а также строение трихоцистов и рабдит, сократительной вакуоли и нефридиев у инфузорий и бескишечных червей имеют совершенно различное строение. Теория целлюляризации опирается в главном лишь на внешние аналогии и конвергентные сходства, действительно имеющие место в морфологии некоторых Protozoa и Metazoa.
Эмбриологическим основанием гипотезы целлюляризации служит поверхностное дробление яиц членистоногих. При таком типе дробления деление ядер сперва не сопровождается делением цитоплазмы. Клеточные границы появляются одновременно и сравнительно поздно.
Гипотезы целлюляризации подверглись решительной критике со стороны В.А. Догеля, В.Н. Беклемишева, А.А. Захваткина, А.В. Иванова, О.М. Ивановой-Казас и других крупных зоологов. Суть этой критики, вкратце, состоит в следующем.
Во-первых, ее авторы указали на несостоятельность эмбриологического аргумента. Дело в том, что членистоногие – это животные, далеко отстоящие от истоков многоклеточности, и потому едва ли могли сохранить примитивные формы развития. Дробление яйца членистоногих, вне всякого сомнения – это результат далеко зашедшей специализации. Низшие же многоклеточные имеют совершенно иной ход онтогенеза.
Кроме того, исходя из гипотезы целлюляризации, в онтогенезе многоклеточного животного все ткани должны были бы дифференцироваться сразу после синцитиального дробления, и прямо на месте. В действительности же в ходе индивидуального развития многоклеточных (при гаструляции и органогенезе) наблюдаются последовательная дифференциация и значительные перемещения клеточных масс.
Во-вторых, высокоорганизованные простейшие – это слишком специализированные существа, чтобы дать начало животным с организацией принципиально иного типа. Подобное предположение противоречит одному из фундаментальных законов эволюции, гласящем, что у истоков эволюционно молодой группы организмов всегда находится не самый совершенный представитель из числа эволюционных предшественников.
В-третьих, гипотезы целлюляризации не имеют экологического обоснования. В этой связи, разделение тела простейшего на клетки выглядит беспричинным.
4. "Строение" предка Metazoa согласно разным взглядам на его происхождение.
- Гастрея (по гипотезе Геккеля) - двуслойный организм. Наружный слой играл роль кожного покрова, а внутренний слой - роль кишечника. Бластопор функционировал как рот. Среди современных животных ближе всего к этой гипотетической стадии филогенеза стоят наиболее простые представители кишечнополостных, тело которых состоит всего из двух клеточных слоев.
Но. Геккель механически каждую стадию развития механически считал простой копией какой-то эволюционной стадии, не заботясь об экологической и физиологической стороне вопроса. Он не думал о том, какие биологические преимущества должны были дать предполагаемые эволюционные изменения. Поэтому гастрея представляется не живым существом, а отвлеченной схемой.
Критика (Мечников): Образование двухслойной организации путем инвагинации - ошибочное представление. Для самых примитивных животных характерна мультиполярная иммиграция или деляминация. Инвагинация в процессе эволюции появляется позднее. Внутриклеточное пищеварение первично, полостное вторично.
Но! Сама идея колониального происхождения важна.
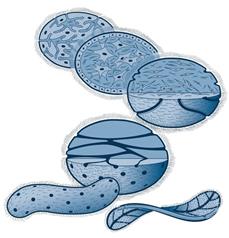 -Фагоцителла (по гипотезе Мечникова) - гипотетический организм, внешне напоминающий личинку многих губок и кишечнополостных. Состоит из двух групп клеток: поверхностные жгутиковые, выполняющие локомоторную функцию - кинобласт, внутренняя масса амебоидных клеток, ответственных за питание - фагоцитобласт. Предки фагоцителлы - простейшие с анимальным типом питания, жгутиконосцы, способные принимать амебоидную форму. Достижение двухслойной организации за счет мультиполярной иммиграции и деляминации (как примитивные формы гаструляции). Образование рта: в результате приспособления для захвата более крупной добычи.
-Фагоцителла (по гипотезе Мечникова) - гипотетический организм, внешне напоминающий личинку многих губок и кишечнополостных. Состоит из двух групп клеток: поверхностные жгутиковые, выполняющие локомоторную функцию - кинобласт, внутренняя масса амебоидных клеток, ответственных за питание - фагоцитобласт. Предки фагоцителлы - простейшие с анимальным типом питания, жгутиконосцы, способные принимать амебоидную форму. Достижение двухслойной организации за счет мультиполярной иммиграции и деляминации (как примитивные формы гаструляции). Образование рта: в результате приспособления для захвата более крупной добычи.
-Галлертоид (по Грасгоффу) -
Галлертоид - это название гипотетическогой реконструкции первых многоклеточных животных, которые могли бы явиться отправной точкой для эволюции всего животного царства. Анатомия и гистология галлертоида была реконструирована в рамках научно-исследовательской работы над Франкфуртской теорией эволюции на основе исследований морфологии и гистологии дошедших до нас организмов. Ведущим для реконструкции этих первых многоклеточных животных был вопрос о том, как могло возникнуть типичное для животных устройство тела.
Что касается животных, типичным является построение тела из соединительной ткани (коллагена) со встроенным в эту сеть клетками. В дополнение к этому волокнистые структуры эпителия, плотного слоя клеток, необходимые для того, чтобы разграничить волокнистую сеть и окружающую среду (воду). Галлертоиды, вероятно, были небольшими, несколько миллиметров величины животными, которые имели различные формы тела и были пронизаны системой более- менее частых каналов. Эти каналы были выстланы реснитчатым эпителием и служили для потребления пищи. Частицы транспортировались через каналы и там поглощались клетками, а извлеченные вещества распределялись по всему организму.
Галлертоид как первое многоклеточное животное сочетает в себе все типичные для животных конструктивные элементы. На основе этой конструкции в конечном счете, могут быть реконструированы основные направления эволюции животного царства. На сегодняшний день есть организмы, которые близки к гистологической структуре галлертоида, гребневики.
Происхождение галлертоида в соответствии с Франкфуртской теорией эволюции было реконструировано и описано как процесс внутренней компартментализации. В многоядерных одноклеточных организмах, чьи тела пересекают широкие каналы и трубочки системы эндоплазматической сети, волокна (полисахариды и полипептиды), первоначально включались в процесс накопления энергии. Посредством включения волокнистых белков, изменяются механические свойства тела, оно затвердевает, его констистенция становится слизеподобной с гелеобразными свойствами. Эта масса поддерживала и стабилизировала тело ранних многоядерных простейших, их организм был способен принимать более крупные формы. Окружающая цитоплазма, таким образом, растягивалась и была отгорожена стенками эндоплазматического ретикулума. Так как эти стенки имели ту же структуру, что и внешняя клеточная мембрана (точнее, эндоплазматический ретикулум содержит блоки клеточной мембраны) образовалась многоклеточная конструкция тела с синцитиальной структурой.
5. Гипотеза "гастреи" и ее эволюционные последствия.
Классическая теория гастреи в том виде, каком ее создал Э. Геккель, сыграла большую положительную роль в развитии эволюционной зоологии. Однако уже многим современникам Геккеля были видны недостатки этой гипотезы. Гипотеза гастреи была лишена физиологического обоснования, а представляла собой голую морфологическую схему. Эта концепция предполагала внезапное возникновение полостного пищеварения, игнорируя факты широкого распространения фагоцитоза у простейших и низших многоклеточных. Гипотеза гастреи основывалась на «буквальном» толковании всех стадий развития как рекапитуляции последовательных этапов филогенеза многоклеточных. При этом за основу принимался онтогенез таких высокоорганизованных форм, как ланцетник или сагитта, в развитии которых двухслойный зародыш формируется путем инвагинации, т. е. путем впячивания стенки бластулы внутрь. Однако у низших многоклеточных (губок, гидроидных полипов) гаструляция путем инвагинации не встречается. Двухслойный зародыш здесь формируется путем иммиграции или деляминации.
Принципиальные трудности классической теории гастреи сохраняются и в тех модификациях, которые предлагали О. Бютчли и Г. Иегерстен, при этом в каждой из этих концепций появляются и специфические противоречия, отсутствующие в построениях Э. Геккеля. Так, гипотеза плакулы О. Бютчли основывается на придании рекапитуляционного значения весьма экзотическому способу гаструляции — изгибанию двухслойной пластинки, — встречающемуся у отдельных представителей довольно высокоорганизованных групп животных (у нескольких видов нематод, некоторых малощетииковых червей, асцидий). Инвагинационная гаструла, послужившая Э. Геккелю моделью формирования двухслойности в филогенезе при том что она не встречается у низших многоклеточных, широко распространена в животном царстве в отличие от редко встречающейся плакулы.
Умозрительная, хотя и весьма детально разработанная, гипотеза билатерогастреи сталкивается с теми же эмбриологическими трудностями, как и классическая гипотеза геккелевской гастреи: она предполагает первичной и исходной инвагинационную гаструлу, не встречающуюся у низших многоклеточных. Однако гипотеза билатерогастреи предполагает уплощенную форму бластул и первичную билатеральнную симметрию в онто- и филогенезе многоклеточных. Это второе предположение особенно находится в противоречии с эмбриологическими данными. В онтогенезе низших многоклеточных (губок и кишечнополостных) ранние стадии развития не имеют никаких следов билатеральной симметрии. Наблюдающиеся у личинок некоторых коралловых полипов элементы билатеральности не могут считаться доказательством справедливости теории билатерогастреи, так как стадии дробления и гаструляции у этих животных всегда радиально-симметричны, а билатеральная симметрия возникает лишь у некоторых личинок в результате их приспособления к ползающему образу жизни. Важно что в онтогенезе билатерально-симметричных животных ранние стадии развития (дробление, гаструляция) имеют выраженную радиальную симметрию. Билатеральная симметрии появляется лишь на более поздних этапах развития. Никаких следов исходной билатеральной симметрии, свойственной еще бластулообразным стадиям филогенеза многоклеточных (если следовать взглядам Иегерстена), мы не находим в онтогенезах ни у губок, ни у кишечнополостных, ни у низших Bilateria. Вообще выведение радиально-симметричных кишечнополостные из билатерально-симметричной билатерогастреи, принимаемое Г. Иегерстеном, вызывает огромные трудности и противоречат всей совокупности фактов строения и развития кишечнополостных (Иванов, 1968).
6. Гипотеза "фагоцителлы".
Поскольку у Metazoa полость дробления обычно появляется очень рано и зародыш быстро превращается в бластулу, предком многоклеточных Мечников считал бластулообразную колонию жгутиконосцев. Он полагал, что тотальное дробление многоклеточных следует выводить из деления жгутиконосцев: первые меридиональные деления дробящегося яйца представляют собой наследие от флагелятных предков, поскольку для жгутиконосцев характерно именно продольное деление. Исходную шарообразную форму колонии Мечников пытается также объяснить исходя из продольного деления жгутиконосцев. Если деление клетки все время происходит продольно, то получается пластинка, но если третье деление изменится, станет поперечным, то в результате возникает шаровидная колония клеток. Именно такое изменение направления деления и произошло в филогенезе. Таким образом, предком Metazoa была колония, у которой направления деления чередовались в трех координатных плоскостях. Мечников думал, что образование двуслойной стадии произошло не путем впячивания, а путем иммиграции — внедрения отдельных клеток в полость бластулы, в результате чего образовался зачаток энтодермы. Эволюционные истоки такой иммиграции он видел в явлении фагоцитоза.
Питание первичного метазоона, по Мечникову, совершалось теми же клетками, которые служили для движения, т. е. жгутиковыми клетками путем внутриклеточного пищеварения (фагоцитоза). И. И. Мечников предположил, что перегруженные пищей клетки легко теряли жгутик и уходили в полость организма, затем они снова могли выходить на поверхность и формировать жгутик. Так получилась первая факультативная дифференциация на наружный слой клеток — «кинобласт» — и на внутреннюю клеточную массу — «фагоцитобласт». Эта дифференциация затем закрепилась в эволюции, и образовался компактный организм — паренхимелла, моделью которого он считал личинку губок — паренхимуллу. Позднее Мечников назвал этот организм фагоцителлой. Это и был общий предок многоклеточных животных.
Дальнейшая судьба фагоцителлы такова. Некоторые ее потомки перешли к сидячему образу жизни и дали начало губкам. Другие стали ползать, приобрели билатеральную симметрию и ротовое отверстие. От них произошли бескишечные плоские черви турбеллярии, у которых еще нет кишечника и пищеварение совершается в лакунах паренхимы и в блуждающих клетках — фагоцитах. Третьи, сохранив плавающий образ жизни, приобрели рот, испытали эпителизацию фагоцитобласта и превратились в первичных кишечнополостных — родоначальников сидячих полипов.
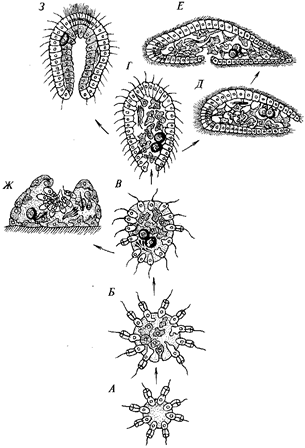 Таким образом, гипотеза И. И. Мечникова объясняла с эволюционной точки зрения все главные этапы онтогенеза Metazoa и предлагала новые филогенетически обоснованные представления о первичных зародышевых листках и их дальнейшей эволюции. На этой основе Мечников нарисовал вполне правдоподобную гипотетическую картину эволюционного становления Metazoa и первых этапов их филогенетического развития, картину, которая хорошо объясняет множество эмбриологических и сравнительно-анатомических факторов, непонятных с точки зрения других гипотез.
Таким образом, гипотеза И. И. Мечникова объясняла с эволюционной точки зрения все главные этапы онтогенеза Metazoa и предлагала новые филогенетически обоснованные представления о первичных зародышевых листках и их дальнейшей эволюции. На этой основе Мечников нарисовал вполне правдоподобную гипотетическую картину эволюционного становления Metazoa и первых этапов их филогенетического развития, картину, которая хорошо объясняет множество эмбриологических и сравнительно-анатомических факторов, непонятных с точки зрения других гипотез.
А. А. Захваткин в 1949 г. выдвинул гипотезу о происхождении многоклеточных от колониальных жгутиконосцев на основе палинтомии — особой формы бесполого размножения путем последовательных делений клетки без стадий роста получающихся дочерних клеток. Такое деление является, по его мнению, прообразом дробления яйца у Metazoa.
Другой путь эволюционного формирования Metazoa предложил А. В. Иванов в конце 60-х гг., считавший, что гипотетические исходные колонии жгутиконосцев не были палинтомическими и вообще мало отличались от шаровидных колоний современных воротничковых жгутиконосцев.
Иванов принимает за основу теорию фагоцителлы Мечникова. Однако прообразом фагоцителлы он считает не личинку губок, а примитивное плоское многоклеточное трихоплакс, являющееся единственным представителем типа Пластинчатые (Placozoa). Схема возникновения многоклеточных, согласно Иванову, представлена на рисунки.
Поскольку в эмбриогенезе низших многоклеточных двуслойных зародыш образуется чаще путем иммиграции, большинство зоологов считают, что именно таким путем и шло преобразование шаровидной колонии жгутиконосцев в первый многоклеточный организм. При этом у предковых форм многоклеточных образование двух клеточных слоев сопровождалось специализацией клеток и колония жгутиконосцев превратилась в единый многоклеточный организм. Наружный слой сохранил двигательную и чувствительную функции, а внутренний — пищеварительную и половую.
7. Гипотеза "галлертоида" (Боник, Грасгофф) и ее эволюционные последствия.
(Похож на паренхимулу, но у той внутри клетки плотно расположены)
В 1976 г. была выдвинута еще одна гипотеза происхождения Metazoa (Bonik et al., 1976), которая получила название гипотезы Галлертоида. «Gallert» в переводе с немецкого означает «желатина», «студень». Галлертоид - гипотетический первичный Metazoa, возникший в процессе эволюции из планктонной шарообразной колонии жгутиконосцев, внутри которой имеются амебоидные клетки. У Галлертоида, устроенного таким же образом, появился студенистый экстрацеллюлярный матрикс. Такой гипотетический организм предлагается рассматривать в качестве предка всех остальных Metazoa. Позднее эта гипотеза была развита и несколько модифицирована одним из ее создателей (Grasshoff, 1993). Описанный выше гипотетический организм был назван ранним Галлертоидом, от которого независимо произошли два других гипотетических животных - Плакулоид и поздний Галлертоид. Последний сохраняет шарообразную или овальную форму, но его клетки, находящиеся под поверхностным эпителиообразным слоем, соединяются длинными псевдоподиеподобными выростами, формируя трехмерную сеть - соединительную ткань. Все пространство между клетками этой сети заполнено экстрацеллюлярным матриксом. Поздний Галлертоид дал начало Spongia, Cnidaria и Ctenophora. Плакулоид перешел к бентосному образу жизни. Тело его стало более плоским, однако его внутренние клетки остались амебоидными и свободно перемещались в студенистом экстрацеллюлярном матриксе. По мнению Грассгофа, плакулоид дал начало древним Placozoa и, вероятно, предкам животных венда.
Возможный эволюционный путь преобразования раннего Галлертоида в трихоплакса подробно рассмотрен в работах Сайда и Шиеруотера (Syed, Schierwater, 2002a, b). Они назвали свою гипотезу галлертоид-плакулоидной моделью. Согласно ей планктонный галлертоидный предок Placozoa перешел к обитанию в бентосе, поскольку стал питаться детритом. Поздний Плакулоид имел уплощенное тело. Внутри он содержал амебоидные клетки и экстрацеллюлярный матрикс. Такой организм, как считал и Грассгоф (Grasshoff, 1993), возможно, дал начало дисковидным формам вендского периода, а также от него произошли первичные Placozoa. Они сохранили экстрацеллюлярный матрикс и амебоидные клетки, которые, поскольку тело ползающего по субстрату животного стало очень плоским, располагались в один тонкий слой. Позднее в процессе эволюции Placozoa они объединились в единый синцитий. Затем возникла морфологическая и функциональная дифференцировка клеток дорсального и вентрального слоев. Наконец, произошла редукция экстрацеллюлярного матрикса. Так сформировался T. adhaerens, который приобрел способность размножаться с помощью бродяжек.
 Следует отметить, что ни один из рассматриваемых этапов эволюции Галлертоида, начиная от колонии жгутиковых протистов и кончая гипотетическими представителями Placozoa не имеют каких-либо аналогий в строении современных животных (или их эмбрионов). Так, не известны колониальные жгутиконосцы, которые содержали бы внутри амебоидные клетки. Никем не описаны метазои (или их эмбрионы), хотя бы частично похожие по своему строению на Галлертоид, Плакулоид или первичные Placozoa. То есть рассматриваемые гипотезы являются сугубо умозрительными. Быть может поэтому они не получили достаточно широкого распространения и признания зоологов. Тем не менее высказанное авторами галлертоид-плакулоидной модели (Syed, Schierwater, 2002a, b) предположение о том, что Placozoa, возможно, являются группой, родственной «Vendobionta» - дисковидным беспозвоночным эдиакарского периода, находит свое подтверждение в данных экспериментальной работы Шварца (Schwartz, 1984), о которой мы говорили ранее, и свидетельствующей о том, что тело T. adhaerens по своей природе является бициклическим.
Следует отметить, что ни один из рассматриваемых этапов эволюции Галлертоида, начиная от колонии жгутиковых протистов и кончая гипотетическими представителями Placozoa не имеют каких-либо аналогий в строении современных животных (или их эмбрионов). Так, не известны колониальные жгутиконосцы, которые содержали бы внутри амебоидные клетки. Никем не описаны метазои (или их эмбрионы), хотя бы частично похожие по своему строению на Галлертоид, Плакулоид или первичные Placozoa. То есть рассматриваемые гипотезы являются сугубо умозрительными. Быть может поэтому они не получили достаточно широкого распространения и признания зоологов. Тем не менее высказанное авторами галлертоид-плакулоидной модели (Syed, Schierwater, 2002a, b) предположение о том, что Placozoa, возможно, являются группой, родственной «Vendobionta» - дисковидным беспозвоночным эдиакарского периода, находит свое подтверждение в данных экспериментальной работы Шварца (Schwartz, 1984), о которой мы говорили ранее, и свидетельствующей о том, что тело T. adhaerens по своей природе является бициклическим.
Дата: 2019-02-02, просмотров: 1164.