112
| Гипоталамус |
| ХИМИЧЕСКАЯ КЛАССИФИКАЦИЯ ГОРМОНОВ Гормоны можно разделить на два основных типа: стероидные и нестероидные. Химическая структура первых напоминает структуру холестерина и большинство этих гормонов являются его производными. Поэтому они растворяются в ли- 8 .-из 113 |
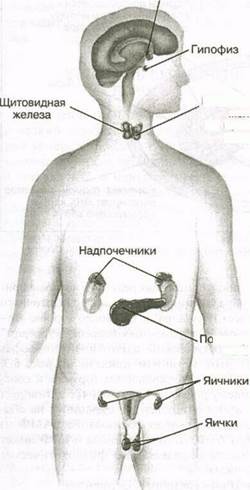
Рис. 6.1. Расположение основных эндокринных органов
-только на опреде-
ют на многие ткани, другие ленные клетки-мишени.
ПРИРОДА ГОРМОНОВ
Гормоны участвуют в большинстве физиологических процессов, поэтому их действие имеет большое значение для многих аспектов мышечной и спортивной деятельности. Прежде чем приступить к изучению функций гормонов, рассмотрим их сущность. Далее мы рассмотрим химическую природу гормонов и общие механизмы их действия.
пидах и довольно легко диффундируют через клеточные мембраны. К этой группе относятся гормоны, экскретируемые
• корковым веществом надпочечника (такие, как кортизол и альдостерон);
• яичниками (эстроген и прогестерон);
• яичками (тестостерон);
• плацентой (эстроген и прогестерон). Нестероидные гормоны не являются жирорастворимыми, поэтому они не могут легко пересекать клеточные оболочки. Группу нестероидных гормонов можно разделить на два класса: белковые, или пептидные, гормоны и производные аминокислот. К последнему классу относятся два гормона щитовидной железы (тироксин и три-йодтиронин) и два гормона мозгового вещества надпочечников (адреналин и норадреналин). Остальные нестероидные гормоны относятся к классу белковых, или пептидных, гормонов.
ФУНКЦИИ ГОРМОНОВ
Поскольку гормоны перемещаются с кровью, они вступают в контакт практически со всеми тканями тела. Почему же тогда их действие ограничено определенными клетками-мишенями? Оно обусловлено наличием специальных рецепторов в тканях-мишенях. Взаимодействие гормона и его определенного рецептора сравнивают с принципом взаимодействия замка (рецептора) и ключа (гормона), когда лишь подходящим ключом можно открыть соответствующий замок. Взаимодействие гормона и его рецептора называют комплексом гормона-рецептора.
Каждая клетка содержит от 2 000 до 10 000 рецепторов. Рецепторы нестероидных гормонов располагаются на оболочке клетки, тогда как рецепторы стероидных гормонов находятся в ее цитоплазме либо ядре. Каждый гормон характеризуется высокой степенью специфичности по отношению к данному типу рецепторов и связывается только с определенными (специфичными) рецепторами, воздействуя, таким образом, только на ткани, содержащие эти рецепторы.
Многочисленные механизмы позволяют гормонам контролировать действия клеток. Рассмотрим основные способы действия стероидных и нестероидных гормонов.
Стероидные гормоны
Как уже указывалось, стероидные гормоны являются липидорастворимыми и легко проходят через клеточную оболочку. Механизм их действия показан на рис. 6.2. Находясь внутри клетки, сте-роидный гормон связывается со специфичными для него рецепторами. Образовавшийся комплекс гормон — рецептор проникает в ядро и связывается с частью ДНК клетки, активируя определенные гены. Этот процесс называется непосред-
| Стероидный гормон проникает в клетку Клеточная мембрана |
| мРНК обеспечивает белковый синтез в цитоплазме |
| Гормон привязывается кК специфичному рецептору в цитоплазме или ядре |
| Ядро |
| мРНК "покидает" ядро |
|
|
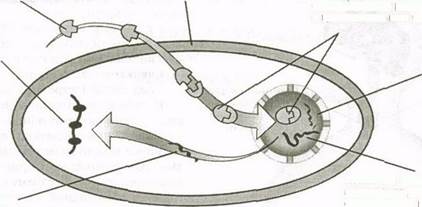
| Комплекс гормон—рецептор активирует ДНК клетки, образующую мРНК |
Рис. 6.2. Механизм действия стероидного гормона, приводящий к непосредственной активации генов
ственной активацией генов. В ответ на нее в ядрах происходит синтез мРНК. Затем мРНК поступает в цитоплазму и обеспечивает белковый синтез. Эти белки могут быть
• ферментами, оказывающими многочисленные воздействия на клеточные процессы;
• структурными белками, которые используются для роста и восстановления тканей;
• регуляторными белками, способными изменить функцию ферментов.
Нестероидные гормоны
Поскольку эти гормоны не могут легко проходить через клеточную оболочку, они вступают во взаимодействие с определенными рецепторами вне клетки, на ее оболочке. Молекула нестероидного гормона, прикрепляясь к своему рецептору, вы
зывает серию ферментных реакций, которые приводят к образованию второго внутриклеточного переносчика ("курьера"). Наиболее хорошо изученным и распространенным вторым "курьером" является циклический аденозинмонофосфат (цАМФ). Этот механизм показан на рис. 6.3. В данном случае прикрепление гормона к соответствующему рецептору на оболочке активирует фермент аденилатциклазу, находящийся на оболочке. Он катализирует образование цАМФ из клеточного АТФ. Образовавшийся цАМФ может затем вызвать определенные физиологические реакции, включая:
• активацию клеточных ферментов;
• изменение проницаемости оболочки;
• обеспечение белкового синтеза;
• изменение клеточного метаболизма;
• стимулирование клеточных выделений.
| Нестероидные гормоны не могут проходить через клеточную мембрану |
| Гормон привязывается к специфичному рецептору на клеточной мембране |
| Комплекс гормон—рецеп тор активирует аденилат циклазу в клетке |
| Клеточная мембрана |
| Адемилатциклаза образует цАМФ |
| цАМФ активирует протеинкиназы (ферменты), что приводит к клеточным изменениям и гормональным эффектам |
|
|
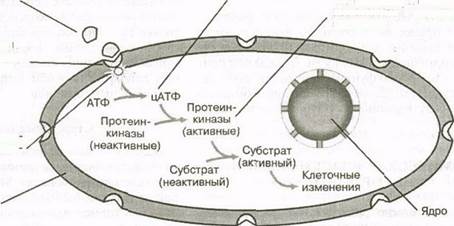
Рис. 6.3. Механизм действия нестероидного гормона с использованием
второго переносчика (цАМФ) в клетке
114
Таким образом, нестероидные гормоны, как правило, активируют систему цАМФ клетки, что приводит к изменениям внутриклеточных функций.
КОНТРОЛЬ ВЫДЕЛЕНИЯ ГОРМОНОВ
Выделение гормонов носит кратковременный и резкий характер, что обусловливает колебания уровней определенных гормонов плазмы в течение коротких периодов времени, например в течение часа и меньше. Вместе с тем колебание их уровней наблюдается и в течение более продолжительного времени, демонстрируя суточные и даже месячные циклы (подобно месячным менструальным циклам). Откуда тогда железы внутренней секреции знают, когда выделять гормоны?
Количество рецепторов
Уровни содержания гормонов в плазме не всегда отражают действительную активность гормонов, поскольку количество рецепторов на клетке может измениться, повышая или понижая ее чувствительность к определенному гормону. Чаще всего увеличение количества определенных гормонов приводит к сокращению числа соответствующих рецепторов. Когда это происходит, чувствительность клетки к данному гормону снижается, поскольку сокращается число рецепторов, с которыми может связаться гормон. Это называется десенсибилизацией, или пониженной регуляцией. У некоторых людей с избыточной массой тела, например, количество инсулиновых рецепторов на клетках понижено. Их организм реагирует на это усиленной секрецией инсулина из поджелудочной железы, вследствие чего повышаются уровни инсулина в плазме. Чтобы достичь такой же степени регуляции глюкозы плазмы как у обычных, физически здоровых людей, необходимо, чтобы их организм выделял намного больше инсулина.
Наоборот, клетка может отреагировать на продолжительное наличие большого количества гормона увеличением числа рецепторов. В этом случае ее чувствительность к данному гормону повышается, поскольку большее его количество может быть привязано в данный момент. Это — повышенная регуляция. Кроме того, иногда один гормон может восприниматься рецепторами другого гормона.
В ОБЗОРЕ...
1. Гормоны делятся на стероидные и нестероидные. Стероидные гормоны липидорастворимые и большинство из них образуется из холестерина. Нестероидные гормоны — белки, пептиды или аминокислоты.
2. Гормоны, как правило, экскретируются в кровь и циркулируют по организму, воздействуя только на клетки-мишени. Их действие по принципу "замок — ключ" заключается в связывании их специфичными рецепторами, содержащимися только в тканях-мишенях.
3. Стероидные гормоны проходят через клеточную оболочку и связываются с рецепторами, находящимися внутри клетки. С помощью механизма, получившего название непосредственной активации генов, они вызывают белковый синтез.
4. Нестероидные гормоны не могут так же легко проникать в клетки, поэтому они связываются с рецепторами, находящимися на оболочке. Это приводит к активации второго "курьера" клетки, что, в свою очередь, вызывает многочисленные клеточные процессы. :
115
5. Секреция большинства гормонов осуществляется на основании отрицательной обратной связи.
6. Количество рецепторов для данного гормона может изменяться для удовлетворения потребностей организма. Повышенная регуляция означает увеличение числа рецепторов, пониженная — уменьшение. Эти два процесса изменяют чувствительность клетки по отношению к гормонам.
Задняя доля гипофиза
Задняя доля гипофиза представляет собой отросток или вырост нервной ткани гипоталамуса. Именно поэтому ее часто называют нейрогипофизом. Она выделяет два гормона — антидиуретический гормон (АДГ, или вазопрессин) и окситоцин. В сущности, их производит гипоталамус. Они перемещаются вниз по нервной ткани и содержатся в пузырьках нервных окончаний задней доли гипофиза. В ответ на нервные импульсы, поступающие из гипоталамуса, эти гормоны попадают в кровь.
Из этих двух гормонов только АДГ играет важную роль в процессе мышечной деятельности. Он, в частности, обеспечивает сохранение воды в организме, повышая ее проникновение в канальцы почек, вследствие чего в мочу поступает меньше воды.
О влиянии физических нагрузок на секрецию задней доли гипофиза сведений мало. Известно,
Простагландины
Простагландины, хоть и не являются гормонами, очень часто рассматриваются как таковые. Они являются производными жирной кислоты — арахидоновой — и связаны с мембранами почти всех клеток тела. Обычно Простагландины действуют как местные, или локальные, гормоны, воздействуя лишь на тот участок, где они образовались. В то же время некоторые из них, перемещаясь с кровью, воздействуют на более отдаленные ткани. Выделение простагландинов может активировать другие гормоны, а также вызывать локальные повреждения. Их функ
ции многочисленны, поскольку существует несколько различных типов простагландинов. Очень часто они усиливают действия других гормонов. Кроме того, они могут непосредственно воздействовать на кровеносные сосуды, повышая их проницаемость (что способствует их набуханию) и обеспечивая расширение. В этой связи они — важные медиаторы воспалительной реакции. Кроме того, они повышают чувствительность нервных 9кончаний болевых волокон и, таким образом, участвуют в развитии воспаления и ощущении боли.
116
Таблица 6.1. Железы внутренней секреции, их гормоны, органы-мишени и основные функции
| Эндокринная железа | Гормон | Орган-мишень | Основные функции |
| Гипофиз | |||
| передняя доля | Гормон роста | Все клетки тела | Обеспечивает рост и развитие всех тканей тела в пе |
| риод полового созревания; повышает интенсивность | |||
| белкового синтеза; увеличивает мобилизацию жиров | |||
| и использует их в качестве источника энергии; | |||
| снижает интенсивность утилизации углеводов | |||
| Тиротропин, или | Щитовидая | Регулирует количество производимых и выделяемых | |
| тиреостимулирую- | железа | щитовидной железой тироксина и трийодтиронина | |
| щий гормон | |||
| Адренокортико- | Корковое | Регулирует секрецию гормонов из коркового ве | |
| тропин | вещество | щества надпочечника | |
| надпочечника | |||
| Пролакпю | Молочные | Стимулирует развитие молочных желез и секрецию | |
| железы | молока | ||
| Фолликулостиму- | Яичники,яички | Стимулирует рост фолликулов в яичниках и обес | |
| лирующий гормон | печивает секрецию эстрогена из яичников, а также | ||
| (ФСГ) | образование спермы в яичках | ||
| Лютеинизирую- | Яичники, яички | Обеспечивает секрецию эстрогена и прогестерона, | |
| щий гормон (ЛГ) | вызывает разрыв фолликулов, обеспечивающий | ||
| выделение яйцеклетки; стимулирует секрецию тес- | |||
| тостерона из яичек | |||
| задняя доля | Антидиуретичес | Почки | Способствует регулированию выделения воды поч |
| (от гипоталамуса) | кий гормон (АДГ, | ками; повышает давление крови вследствие су | |
| или вазопрессин) | жения сосудов | ||
| Окситоцин | Матка, молоч | Стимулирует сокращение мышц матки, секрецию | |
| ные железы | молока | ||
| Щитовидная железа | Тироксин и | Все клетки тела | Повышает интенсивность клеточного метаболизма, а |
| трийодтиронин | также частоту и сократительную способность сердца | ||
| Кальцитонин | Кости | Регулирует концентрацию ионов кальция в крови | |
| Паращитовидная | Паратгормон, или | Кости, почки, | Регулирует концентрацию ионов кальция во вне |
| железа | паратиреоидный | кишечник | клеточной жидкости, воздействуя на кости, почки |
| гормон | и кишечник | ||
| Надпочечники | |||
| мозговое | Адреналин | Большинство | Мобилизует гликоген; усиливает кровоток в ске |
| вещество | клеток тела | летной мышце; повышает ЧСС, сократительную | |
| способность сердца и потребление кислорода | |||
| Норадреналин | Тоже | Сужает артериолы и венулы, тем самым повышая | |
| давление крови | |||
| корковое | Минералкортико- | Почки | Увеличивает задержку натрия и выделение калия |
| вещество | иды (альдостерон) | через почки | |
| Глюкокортикоиды | Большинство | Регулирует метаболизм углеводов, жиров и белков; | |
| (кортизол) | клеток тела | противовоспалительное действие | |
| Андрогены и | Яичники, мо | Развитие половых признаков | |
| эстрогены | лочные железы, | ||
| яички | |||
| Поджелудочная | Инсулин | Все клетки тела | Регулирует уровни глюкозы крови, понижая содер |
| железа | жание глюкозы; повышает утилизацию глюкозы и | ||
| синтез белков | |||
| Глюкагон | Тоже | Повышает концентрацию глюкозы в крови; стиму | |
| лирует расщепление белков и жиров | |||
| Соматостатин | Островки Лан- | Снижает секрецию инсулина и глюкагона | |
| герганса и желу- | |||
| дочно-кишеч- | |||
| ный тракт | |||
| Половые железы | |||
| яички | Тестостерон | Половые | Обеспечивает развитие половых признаков мужчин |
| органы, мышцы | (яичек, мошонки, полового члена); изменение | ||
| голоса, появление волос на лице; развитие мышц |
117
Таблица 6.1. Окончание
| Эндокринная железа | Гормон | Орган-мишень | Основные функции |
| яичники Экстроген Половые органы, Развитие женских половых признаков и органов жировая ткань; повышенное накопление жира, способствует регуляции м менструального цикла Почки Ренин Корковое вещество Способствует регуляции давления крови надпочечника - Эритропоэтин Костный мозг Образование эритроцитов
| |||
| Примечание. Приведены основные железы внутренней секреции и гормоны, имеющие определенное значение для мышечной и спортивной деятельности.
| |||
например, что введение в кровь концентрированного электролитного раствора вызывает значительное выделение АДГ из гипофиза. Эта способность АДГ сохранять воду в организме существенно снижает риск обезвоживания в условиях значительного потоотделения при выполнении больших физических нагрузок.
На рис. 6.5 показан этот механизм в действии. Вследствие мышечной деятельности и потоотделения в плазме крови повышается концентрация электролитов — процесс, называемый гемокон-центрацией. В результате увеличивается осмотическое давление плазмы. Это — основной физиологический стимул для выделения АДГ. Повышение осмотического давления ощущают осморецепторы, расположенные в гипоталамусе. Он посылает нервные импульсы в заднюю долю гипофиза, стимулируя выделение АДГ, который поступает в кровь, перемещается в почки и обеспечивает задержку воды с тем, чтобы довести концентрацию электролитов в плазме до нормального уровня.
Передняя доля гипофиза
Передняя доля гипофиза, или аденогипофиз, выделяет шесть гормонов в ответ на выделение гипоталамусом стимулирующих или тормозящих факторов (гормонов). Связь между гипоталамусом и передней долей гипофиза осуществляет специальная система кровообращения, транспортирующая эти гормоны из гипоталамуса в переднюю долю гипофиза. Главные функции каждого из гормонов передней доли гипофиза наряду с их стимулирующими и тормозящими влияниями, приведены в табл. 6.2. Физическая нагрузка оказывается значительным стимулом для гипоталамуса, поскольку повышает интенсивность выделения всех гормонов передней долей гипофиза.
Из шести гормонов, выделяемых передней долей гипофиза, четыре являются тройными, т.е. они воздействуют на функции других желез внутренней секреции. Исключение составляют гормон роста и пролактин. Первый — сильный анаболи-ческий агент. Он обеспечивает рост и гипертро-
Таблица 6.2. Гормоны передней доли гипофиза
| Гормон | Регулирующий фактор | Основные функции |
| Адренокортикотропный | Стимулируется кортико- | Обеспечивает выделение гормонов из коркового ве |
| гормон (АКТГ) | тропным гормоном | щества надпочечников |
| Гормон роста (СТГ) | Стимулируется стимулиру | Обеспечивает рост костей и мышц, использование |
| ющим гормоном гормона | жиров в качестве источника энергии, а также эко | |
| роста | номию глюкозы | |
| Ингибируется соматоста- | ||
| тином или ингибирующим | ||
| гормоном гормона роста | ||
| Тиреотропный гормон (ТТГ) | Стимулируется тироид- | Обеспечивает выделение гормонов щитовидной |
| выделяющим гормоном | железы | |
| Фолликулостимулирующий | Стимулируется гонадотро- | У женщин обеспечивает созревание фолликулов в |
| гормон (ФСГ) | пинвыделяющим гормоном | яичниках и образование эстрогена; у мужчин стиму |
| лирует образование спермы | ||
| Лютеинизирующий | Стимулируется гонадотро- | У женщин стимулирует образование эстрогена и |
| гормон(ЛГ) | пинвыделяющим гормоном | прогестерона; у мужчин —тестостерона |
| Пролактин | Стимулируется пролактин- | Обеспечивает лактацию (выделение молока молочной |
| выделяющим гормоном | железой) | |
| Ингибируется пролактин- | ||
| ингибирующим гормоном |
118
Рис. 6.4
Расположение и структура гипофиза
|
|
Передняя доля гормон роста
тиреостимулирующий гормон фолликулостимулирующий гормон лютеинизирующий гормон пролактим адренокортикотропный гормон
| Задняя доля антидиуретический гормон оцитоцин |
Промежуточная доля
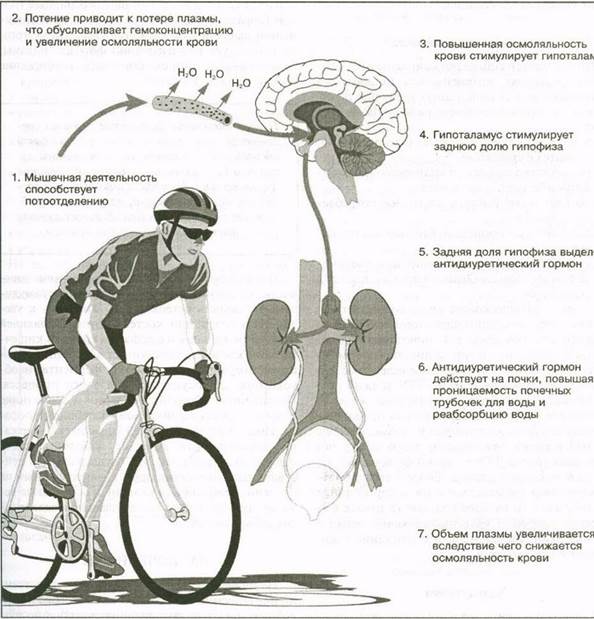
Рис. 6.5. Механизмы сохранения воды антидиуретическим гормоном
119
фию мышц, способствуя транспорту аминокислот в клетки. Кроме того, он непосредственно стимулирует метаболизм жиров (липолиз), усиливая синтез ферментов, участвующих в этом процессе. Уровни содержания гормона роста повышаются при выполнении работы аэробного характера пропорционально интенсивности и остаются повышенными еще некоторое время после завершения работы.
ЩИТОВИДНАЯ ЖЕЛЕЗА
Щитовидная железа расположена вдоль средней линии шеи, непосредственно под гортанью. Она выделяет два гормона, регулирующие обмен веществ, —трийодтиронин (Тд) и тироксин (Т^), а также кальцитонин, который способствует регуляции метаболизма кальция.
Трийодтиронин и тироксин
Эти два гормона обладают одинаковыми свойствами: повышают интенсивность метаболизма практически всех тканей и могут увеличивать интенсивность основного обмена на 60 — 100 %. Они также
• усиливают белковый синтез (следовательно, и синтез ферментов);
• увеличивают размеры и количество митохон-дрий в большинстве клеток;
• обеспечивают быстрое клеточное потребление глюкозы;
• способствуют процессам гликолиза и глюко-неогенеза;
• повышают мобилизацию липидов, увеличивая количество свободных жирных кислот для окисления.
Во время физической нагрузки возрастает выделение тиреостимулирующего гормона (ТТУ) из передней доли гипофиза, регулирующего выделение трийодтиронина и тироксина, поэтому можно предположить, что повышенное вследствие физической нагрузки выделение ТТУ должно стимулировать функцию щитовидной железы. Физические нагрузки действительно приводят к повышению уровня тироксина в плазме, вместе с тем наблюдается своеобразная пауза между повышением уровня ТТУ во время физической нагрузки и тироксина плазмы. Более того, при продолжительной субмаксимальной нагрузке после первоначального резкого подъема (в начале физической нагрузки) уровень тироксина остается относительно постоянным, а содержание трийодтиронина снижается.
Кальцитонин
Кальцитонин снижает концентрацию кальция в плазме. Он воздействует на кости и почки, ин-гибируя активность остеокластов (клеток, резор-
бирующих кость), и тем самым тормозит процесс резорбции костей. Остеокласты могут быть единственной мишенью для кальцитонина в кости. В почках этот гормон увеличивает экскрецию кальция с мочой, уменьшая его реасорбцию из почечных канальцев.
Кальцитонин имеет большое значение для роста костей в детском организме. Этот гормон не является основным регулятором гомеостаза кальция в организме взрослого человека. Вместе с тем, он в определенной степени предотвращает чрезмерную реабсорбцию костей.
ПАРАЩИТОВИДНЫЕ ЖЕЛЕЗЫ
Паращитовидные, или околощитовидные, железы расположены с тыльной стороны щитовидной железы. Они выделяют паратиреоидный гормон (паратгормон) —главный регулятор концентрации кальция в средах организма. Кроме того, он регулирует концентрацию фосфата плазмы. Выделение паратгормона стимулирует понижение уровня кальция в плазме.
Продолжительные физические нагрузки увеличивают остеогенез вследствие повышенной абсорбции Са2"^ в кишечнике, пониженной экскреции Са^ с мочой и усиления образования паратгормона. Иммобилизация или соблюдение постельного режима, напротив, способствуют резорбции костей. В таких случаях уровни паратгормона снижаются
Паратгормон воздействует на три мишени — кости, кишечник и почки. В костях он стимулирует активность остеокластов. Это ведет к увеличению резорбции костей, обусловливающей выделение кальция и фосфата в кровь. В кишечнике он косвенно увеличивает абсорбцию кальция, стимулируя деятельность фермента, необходимого для осуществления этого процесса. Повышенная абсорбция кальция в кишечнике сопровождается увеличенной абсорбцией фосфата. Поскольку паратгормон повышает содержание ионов фосфата в плазме, избыток фосфата должен быть выведен. Это осуществляется действием паратгормона в почках, направленным на усиление реабсорбции кальция и ослабление ре-абсорбции фосфата, обеспечивающим выделение фосфата с мочой.
НАДПОЧЕЧНИКИ
Надпочечники расположены непосредственно над почками. Они состоят из мозгового и коркового вещества, выделяющих совершенно разные гормоны, которые мы рассмотрим в отдельности.
120
122
ПОЧКИ
Рассмотрение в данной главе почек обусловлено тем, что они выделяют гормон эритропоэ-тин, регулирующий образование эритроцитов путем стимуляции клеток костного мозга. Эритроциты играют важную роль, транспортируя кислород в ткани и выводя диоксид углерода. Именно поэтому эритропоэтин играет исключительно важную роль в процессе адаптации организма человека к физическим нагрузкам в условиях высокогорья. Как показывают результаты исследований, одной из адаптационных реакций, обусловленной физическими нагрузками, выполняемыми в условиях высокогорья, является повышенное выделение эритропоэтина, который, в свою очередь, стимулирует образование большего числа эритроцитов, увеличивая кислородтран-спортную способность крови. Именно поэтому некоторые спортсмены вводят в организм эритропоэтин, чтобы увеличить содержание эритроцитов в крови, надеясь таким образом получить преимущество над соперниками. Более подробно это рассматривается в главе 14.
123
Таблица 6.3. Гормональные изменения, обусловленные физическими нагрузками
| Гормон | Реакция на нагрузку | Взаимосвязь | Возможное значение |
| Катехоламины | Увеличение | Более значительное увеличение содер | Повышенное содержание |
| жания при более высокой интенсивнос | глюкозы в крови | ||
| ти, норадреналин > адреналин, меньшее | |||
| увеличение после тренировок | Не известно | ||
| Гормон роста | и | Больше увеличивается у неподготовленных | |
| людей; быстрее снижается после прекра | |||
| щения нагрузки у подготовленных людей | |||
| АКТГ-кортизол | (( | Более значительное увеличение коли | Повышенный глюконеогенез |
| чества при более высокой интенсив-ности | в печени (почках) | ||
| нагрузки; меньшее увеличение после | |||
| субмаксимальных нагрузок | |||
| Тиреотропин- | (( | Повышает обмен тироксина вследствие | Не известно |
| тироксин | тренировок; токсические воздействия не | ||
| наблюдались | |||
| Лютеинизирующий | Не известна | Не известна | " |
| гормон | |||
| Тестостерон | Увеличение | и | и |
| Эстрадиол-прогесте- | Увеличение | Повышается во время лютеальной фазы | Не известно |
| рон | цикла | ||
| Инсулин | Снижение | Меньшая степень снижения после тре | Пониженный стимул к |
| нировок | использованию глюкозы крови | ||
| Глюкагон | Увеличение | Меньшая степень увеличения после тре | Повышенный уровень глюкозы |
| нировок | крови благодаря гликогенолизу | ||
| и глюконеогенезу | |||
| Ренин-ангиотензин- | «' | Такая же степень увеличения у крыс | Задержка натрия с целью |
| альдостерон | после тренировок | поддержания объема плазмы | |
| Антидиуретический | Возможно | Не известна | Задержка воды с целью |
| гормон | увеличение | поддержания объема плазмы | |
| Кальциотонин | Не известна | и | Необходим для обеспечения |
| нормального развития костей | |||
| Эритропоэтин | «с | " | Может играть важную роль в |
| повышении эритропоэза | |||
| Простагландины | Возможно | Могут увеличиваться в ответ на про | Могут обеспечивать локальное |
| увеличение | должающиеся изометрические сокра | расширение сосудов | |
| щения |
1) глюкагон;
2) адреналин;
3) норадреналин;
4) кортизол.
Концентрация глюкозы в плазме во время мышечной деятельности зависит от соотношения между потреблением ее мышцами и выделением печенью. В состоянии покоя выделению глюкозы из печени способствует глюкагон, обеспечивающий расщепление гликогена печени и образование глюкозы из аминокислот. Во время физической нагрузки секреция глюкагона усиливается. Мышечная активность также повышает интенсивность выделения катехоламинов из мозгового вещества надпочечников, и эти гормоны (адреналин и норадреналин) совместно с глюкагоном обеспечива
ют дальнейшее усиление гликогенолиза. Установлено, что во время физической нагрузки уровни кортизола также повышаются. Кортизол, в свою очередь, усиливает катаболизм белков, освобождая аминокислоты для глюконеогенеза, который реализуется в печени. Таким образом, все четыре гормона увеличивают количество глюкозы в плазме, усиливая процессы гликогенолиза и глюконеогенеза. Кроме того, гормон роста повышает мобилизацию свободных жирных кислот и снижает клеточное потребление глюкозы, вследствие чего клетки используют меньше глюкозы (больше глюкозы остается в системе кровообращения), а гормоны щитовидной железы способствуют катаболизму глюкозы и метаболизму жиров.
Количество глюкозы, выделяемой печенью,
124
зависит от интенсивности и продолжительности физической нагрузки. С увеличением интенсивности увеличивается выделение катехоламинов. Это может вынудить печень выделять больше глюкозы, которую используют активные мышцы. На рис. 6.8 показан уровень глюкозы после кратковременной нагрузки "взрывного" типа. К концу 60-секундной спринтерской нагрузки (езда на велосипеде) уровень глюкозы в плазме превышает ее содержание в состоянии покоя, что свидетельствует о том, что количество выделяемой глюкозы превышает ее потребление. Почему не используется это дополнительное количество глюкозы?
Чем выше интенсивность физической нагрузки, тем больше выделяется катехоламинов. Следовательно, значительно повышается интенсивность гликогенолиза. Этот процесс происходит не только в печени, но и в мышцах. Глюкоза, выделяемая печенью, поступает в кровь и становится доступной мышцам. Однако мышцы имеют и другой доступный источник глюкозы — свой собственный гликоген. Мышца использует свои запасы гликогена, прежде чем использовать глюкозу плазмы во время кратковременной физической нагрузки "взрывного" типа. Глюкоза, выделяемая печенью, используется не сразу, а остается в системе кровообращения, повышая уровень глюкозы в плазме. После завершения физической нагрузки уровни глюкозы в плазме снижаются по мере того, как глюкоза поступает в мышцы, восполняя истощенные запасы мышечного гликогена.
Во время физической нагрузки продолжительностью несколько часов интенсивность выделения глюкозы печенью максимально соответству-
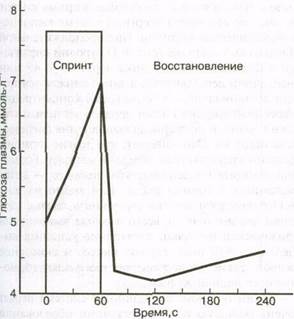
Рис. 6.8. Изменение содержания глюкозы в крови в течение 1 мин спринта на велосипеде
ет потребностям мышцы, и уровень содержания глюкозы в плазме соответствует или слегка превышает ее содержание в состоянии покоя. При увеличении потребления глюкозы мышцей интенсивность ее выделения печенью также повышается. В большинстве случаев уровни глюкозы в плазме не снижаются до тех пор, пока не истощатся запасы гликогена в печени. В этот момент значительно увеличивается количество глюкаго-на. Глюкагон вместе с кортизолом усиливают глю-конеогенез, обеспечивая организм большим количеством энергии.
Рис. 6.9 иллюстрирует изменение уровней содержания адреналина, норадреналина, глюкаго-на, кортизола и глюкозы в плазме во время езды на велосипеде в течение 3 ч. Хотя гормональная регуляция глюкозы при такой продолжительной физической нагрузке не нарушается, содержание гликогена в печени может существенно понизиться. В результате интенсивность выделения глюкозы печенью окажется меньше интенсивности ее потребления. В таких условиях уровень глюкозы может снизиться, несмотря на значительное гормональное стимулирование. В этот момент главную роль в сохранении уровней глюкозы плазмы может сыграть глюкоза, потребляемая во время мышечной деятельности.
Утилизация глюкозы мышцами
Простое выделение достаточного количества глюкозы в кровь не означает, что мышечные клетки получат достаточно глюкозы, чтобы удовлетворить энергетические потребности организма. Глюкоза не просто должна поступать в эти клетки, а потребляться ими. Этот процесс основан на действии инсулина. Как только глюкоза поступает в мышцу, инсулин обеспечивает ее транспорт в волокна.
Удивительно (рис. 6.10), но уровни инсулина плазмы снижаются при продолжительной субмаксимальной нагрузке несмотря на увеличение концентрации глюкозы в плазме и ее более интенсивное использование мышцами. Это
125
явное противоречие между концентрацией инсулина в плазме и потребностью мышцы в глюкозе, напоминает нам, что активность гормонов не всегда определяется их содержанием в крови. В данном случае чувствительность клетки к инсулину может быть такой же важной переменной, как и количество циркулирующего в крови гормона. Физические нагрузки могут усиливать процесс связывания инсулина рецепторами мышечного волокна [5, б]. Мышечные сокращения, непонятно по каким причинам, оказывают инсулиноподобное воздействие при рекруитировании рецепторов: на клетках появляется больше рецепторов и их активность может повышаться, тем самым снижая потребность в значительном количестве инсулина в плазме для транспорта глюкозы через оболочку клетки. Это очень важно, поскольку во время мышечной деятельности четыре гормона пытаются выделить глюкозу из мест ее хранения и образовать новую глюкозу. Их действию противостоит повышенное количество инсулина, направленное на предотвращение чрезмерного уменьшения количества глюкозы.
РЕГУЛЯЦИЯ МЕТАБОЛИЗМА ЖИРОВ ВО ВРЕМЯ ФИЗИЧЕСКОЙ НАГРУЗКИ
Хотя жиры, как правило, в меньшей степени, чем углеводы, удовлетворяют энергетические потребности организма во время физической нагрузки, мобилизация и окисление свободных жирных кислот играет важную роль для выполнения физической работы, требующей проявления выносливости. Во время такой работы запасы углеводов истощаются и энергетические потребности организма в большей степени удовлетворяются за счет окисления жиров. При пониженных запасах
углеводов (низкие уровни глюкозы плазмы и мышечного гликогена) эндокринная система может ускорить окисление жиров (липолиз), тем самым удовлетворяя энергетические потребности мышц. Процесс липолиза интенсифицируется также в результате повышения уровней адреналина и нор-адреналина.
Вспомним, что свободные жирные кислоты хранятся в форме триглицеридов в жировых клетках и внутри мышечных волокон. Тригли-цериды должны расщепиться, чтобы высвободить свободные жирные кислоты, которые затем транспортируются в мышечные волокна. Интенсивность потребления свободных жирных кислот активной мышцей характеризуется высокой степенью корреляции с концентрацией свободных жирных кислот в плазме. Повышение их концентрации приводит к более интенсивной клеточной утилизации свободных жирных кислот. Мы можем предположить, что повышенная концентрация свободных жирных кислот в плазме усиливает их окисление, поскольку повышенное клеточное потребление свободных жирных кислот способствует более сильному окислению [I]. Следовательно, интенсивность расщепления триглицеридов может частично определять интенсивность утилизации мышцами жиров в качестве источника энергии во время физической нагрузки.
Триглицериды расщепляются до свободных жирных кислот с помощью специального фермента — липазы, активируемой, по меньшей мере, четырьмя гормонами: кортизолом; адреналином;
норадреналином и гормоном роста.
Помимо того, что кортизол играет важную роль в глюконеогенезе, он также ускоряет мобилизацию и использование свободных жирных кислот в качестве источника энергии во время выполнения физической нагрузки. При продолжительной физической нагрузке (рис. 6.11) уровни кортизо-ла в плазме достигают пика через 30 — 45 мин мышечной деятельности, а затем снижаются почти до нормальных. В то же время концентрация свободных жирных кислот продолжает повышаться в течение всего периода выполнения физической нагрузки. Это означает, что другие гормоны должны продолжать активировать липазу. Гормоны, которые продолжают этот процесс, — кате-холамины и гормон роста. Как видно из рис. 6.11,6, содержание этих гормонов в плазме увеличивается в течение всего периода выполнения физической нагрузки, постепенно усиливая выделение свободных жирных кислот и окисление жиров. Такие же воздействия оказывают гормоны щитовидной железы.
Таким образом, эндокринная система играет очень большую роль в регуляции образования АТФ во время мышечной деятельности, а также обеспечивает контроль равновесия между метаболизмом углеводов и жиров.
126
В ОБЗОРЕ...
1. Содержание глюкозы в плазме повышается вследствие комбинированного воздействия глюкагона, адреналина, норадреналина и кор-тизола. Эти гормоны обеспечивают гликогено-лиз и глюконеогенез, тем самым увеличивая количество глюкозы, которую можно использовать в качестве источника энергии. Эти же функции выполняют гормон роста и гормоны щитовидной железы.
2. Инсулин помогает выделенной глюкозе поступить в клетки, где она может быть использована для образования энергии. Однако уровень инсулина снижается при продолжительной физической нагрузке, свидетельствуя о том, что физическая нагрузка сама по себе способствует действию инсулина. Таким образом, при выполнении физической нагрузки требуется меньше гормонов, чем в состоянии покоя.
3. При пониженных запасах углеводов организм переходит на использование жиров в качестве источника энергии. Этому процессу способствуют кортизол, адреналин, норадреналин и гормон роста.
4. Кортизол ускоряет процесс липолиза, выделяя в кровь свободные жирные кислоты, которые могут быть использованы клетками для образования энергии. Уровень кортизола достигает пика, а затем при продолжительной физической работе возвращается к исходному уровню. Когда это происходит, роль кортизола начинают выполнять ка-техоламины и гормон роста.
130
112
| Гипоталамус |
| ХИМИЧЕСКАЯ КЛАССИФИКАЦИЯ ГОРМОНОВ Гормоны можно разделить на два основных типа: стероидные и нестероидные. Химическая структура первых напоминает структуру холестерина и большинство этих гормонов являются его производными. Поэтому они растворяются в ли- 8 .-из 113 |
Рис. 6.1. Расположение основных эндокринных органов
-только на опреде-
ют на многие ткани, другие ленные клетки-мишени.
ПРИРОДА ГОРМОНОВ
Гормоны участвуют в большинстве физиологических процессов, поэтому их действие имеет большое значение для многих аспектов мышечной и спортивной деятельности. Прежде чем приступить к изучению функций гормонов, рассмотрим их сущность. Далее мы рассмотрим химическую природу гормонов и общие механизмы их действия.
пидах и довольно легко диффундируют через клеточные мембраны. К этой группе относятся гормоны, экскретируемые
• корковым веществом надпочечника (такие, как кортизол и альдостерон);
• яичниками (эстроген и прогестерон);
• яичками (тестостерон);
• плацентой (эстроген и прогестерон). Нестероидные гормоны не являются жирорастворимыми, поэтому они не могут легко пересекать клеточные оболочки. Группу нестероидных гормонов можно разделить на два класса: белковые, или пептидные, гормоны и производные аминокислот. К последнему классу относятся два гормона щитовидной железы (тироксин и три-йодтиронин) и два гормона мозгового вещества надпочечников (адреналин и норадреналин). Остальные нестероидные гормоны относятся к классу белковых, или пептидных, гормонов.
ФУНКЦИИ ГОРМОНОВ
Поскольку гормоны перемещаются с кровью, они вступают в контакт практически со всеми тканями тела. Почему же тогда их действие ограничено определенными клетками-мишенями? Оно обусловлено наличием специальных рецепторов в тканях-мишенях. Взаимодействие гормона и его определенного рецептора сравнивают с принципом взаимодействия замка (рецептора) и ключа (гормона), когда лишь подходящим ключом можно открыть соответствующий замок. Взаимодействие гормона и его рецептора называют комплексом гормона-рецептора.
Каждая клетка содержит от 2 000 до 10 000 рецепторов. Рецепторы нестероидных гормонов располагаются на оболочке клетки, тогда как рецепторы стероидных гормонов находятся в ее цитоплазме либо ядре. Каждый гормон характеризуется высокой степенью специфичности по отношению к данному типу рецепторов и связывается только с определенными (специфичными) рецепторами, воздействуя, таким образом, только на ткани, содержащие эти рецепторы.
Многочисленные механизмы позволяют гормонам контролировать действия клеток. Рассмотрим основные способы действия стероидных и нестероидных гормонов.
Стероидные гормоны
Как уже указывалось, стероидные гормоны являются липидорастворимыми и легко проходят через клеточную оболочку. Механизм их действия показан на рис. 6.2. Находясь внутри клетки, сте-роидный гормон связывается со специфичными для него рецепторами. Образовавшийся комплекс гормон — рецептор проникает в ядро и связывается с частью ДНК клетки, активируя определенные гены. Этот процесс называется непосред-
| Стероидный гормон проникает в клетку Клеточная мембрана |
| мРНК обеспечивает белковый синтез в цитоплазме |
| Гормон привязывается кК специфичному рецептору в цитоплазме или ядре |
| Ядро |
| мРНК "покидает" ядро |
|
|
| Комплекс гормон—рецептор активирует ДНК клетки, образующую мРНК |
Рис. 6.2. Механизм действия стероидного гормона, приводящий к непосредственной активации генов
ственной активацией генов. В ответ на нее в ядрах происходит синтез мРНК. Затем мРНК поступает в цитоплазму и обеспечивает белковый синтез. Эти белки могут быть
• ферментами, оказывающими многочисленные воздействия на клеточные процессы;
• структурными белками, которые используются для роста и восстановления тканей;
• регуляторными белками, способными изменить функцию ферментов.
Нестероидные гормоны
Поскольку эти гормоны не могут легко проходить через клеточную оболочку, они вступают во взаимодействие с определенными рецепторами вне клетки, на ее оболочке. Молекула нестероидного гормона, прикрепляясь к своему рецептору, вы
зывает серию ферментных реакций, которые приводят к образованию второго внутриклеточного переносчика ("курьера"). Наиболее хорошо изученным и распространенным вторым "курьером" является циклический аденозинмонофосфат (цАМФ). Этот механизм показан на рис. 6.3. В данном случае прикрепление гормона к соответствующему рецептору на оболочке активирует фермент аденилатциклазу, находящийся на оболочке. Он катализирует образование цАМФ из клеточного АТФ. Образовавшийся цАМФ может затем вызвать определенные физиологические реакции, включая:
• активацию клеточных ферментов;
• изменение проницаемости оболочки;
• обеспечение белкового синтеза;
• изменение клеточного метаболизма;
• стимулирование клеточных выделений.
| Нестероидные гормоны не могут проходить через клеточную мембрану |
| Гормон привязывается к специфичному рецептору на клеточной мембране |
| Комплекс гормон—рецеп тор активирует аденилат циклазу в клетке |
| Клеточная мембрана |
| Адемилатциклаза образует цАМФ |
| цАМФ активирует протеинкиназы (ферменты), что приводит к клеточным изменениям и гормональным эффектам |
|
|
Рис. 6.3. Механизм действия нестероидного гормона с использованием
второго переносчика (цАМФ) в клетке
114
Таким образом, нестероидные гормоны, как правило, активируют систему цАМФ клетки, что приводит к изменениям внутриклеточных функций.
КОНТРОЛЬ ВЫДЕЛЕНИЯ ГОРМОНОВ
Выделение гормонов носит кратковременный и резкий характер, что обусловливает колебания уровней определенных гормонов плазмы в течение коротких периодов времени, например в течение часа и меньше. Вместе с тем колебание их уровней наблюдается и в течение более продолжительного времени, демонстрируя суточные и даже месячные циклы (подобно месячным менструальным циклам). Откуда тогда железы внутренней секреции знают, когда выделять гормоны?
Отрицательная обратная связь
Секреция большинства гормонов регулируется на основании отрицательной обратной связи. Выделение гормонов вызывает определенные изменения в организме, которые, в свою очередь, тормозят их дальнейшую секрецию. Вспомним, как работает комнатный стабилизатор температуры (термостат). При снижении заданной температуры термостат посылает сигнал "в котел парового отопления, который начинает обогревать комнату. Если комнатная температура поднимается до заданного уровня, термостат посылает сигнал прекратить обогревание. Если температура снова снижается ниже заданного уровня, весь цикл начинается сначала. Точно так же процесс секреции определенных гормонов "включает" и "выключает" определенные физиологические изменения.
Гормоны могут воздействовать на специфичные для них ткани-мишени или клетки-мишени на основании сложных взаимодействий с рецепторами, расположенными на клеточной оболочке или в клетке, соответствующими данному гормону
Отрицательная обратная связь — основной механизм, посредством которого эндокринная система поддерживает гомеостаз. Рассмотрим такой пример. При высокой концентрации глюкозы в плазме поджелудочная железа выделяет инсулин, который увеличивает потребление глюкозы клетками, что приводит к снижению ее концентрации. Когда концентрация глюкозы в плазме возвращается к нормальному уровню, выделение инсулина тормозится до тех пор, пока ее уровень снова не повысится.
Количество рецепторов
Уровни содержания гормонов в плазме не всегда отражают действительную активность гормонов, поскольку количество рецепторов на клетке может измениться, повышая или понижая ее чувствительность к определенному гормону. Чаще всего увеличение количества определенных гормонов приводит к сокращению числа соответствующих рецепторов. Когда это происходит, чувствительность клетки к данному гормону снижается, поскольку сокращается число рецепторов, с которыми может связаться гормон. Это называется десенсибилизацией, или пониженной регуляцией. У некоторых людей с избыточной массой тела, например, количество инсулиновых рецепторов на клетках понижено. Их организм реагирует на это усиленной секрецией инсулина из поджелудочной железы, вследствие чего повышаются уровни инсулина в плазме. Чтобы достичь такой же степени регуляции глюкозы плазмы как у обычных, физически здоровых людей, необходимо, чтобы их организм выделял намного больше инсулина.
Наоборот, клетка может отреагировать на продолжительное наличие большого количества гормона увеличением числа рецепторов. В этом случае ее чувствительность к данному гормону повышается, поскольку большее его количество может быть привязано в данный момент. Это — повышенная регуляция. Кроме того, иногда один гормон может восприниматься рецепторами другого гормона.
В ОБЗОРЕ...
1. Гормоны делятся на стероидные и нестероидные. Стероидные гормоны липидорастворимые и большинство из них образуется из холестерина. Нестероидные гормоны — белки, пептиды или аминокислоты.
2. Гормоны, как правило, экскретируются в кровь и циркулируют по организму, воздействуя только на клетки-мишени. Их действие по принципу "замок — ключ" заключается в связывании их специфичными рецепторами, содержащимися только в тканях-мишенях.
3. Стероидные гормоны проходят через клеточную оболочку и связываются с рецепторами, находящимися внутри клетки. С помощью механизма, получившего название непосредственной активации генов, они вызывают белковый синтез.
4. Нестероидные гормоны не могут так же легко проникать в клетки, поэтому они связываются с рецепторами, находящимися на оболочке. Это приводит к активации второго "курьера" клетки, что, в свою очередь, вызывает многочисленные клеточные процессы. :
115
5. Секреция большинства гормонов осуществляется на основании отрицательной обратной связи.
6. Количество рецепторов для данного гормона может изменяться для удовлетворения потребностей организма. Повышенная регуляция означает увеличение числа рецепторов, пониженная — уменьшение. Эти два процесса изменяют чувствительность клетки по отношению к гормонам.
Дата: 2018-12-28, просмотров: 390.