Клетки, воспринимающие гормональный сигнал и реагирующие на него изменением своего метаболизма и функции, называются клетками-мишенями. Эти клетки имеют определенное количество белков-рецепторов, узнающих гормон и связывающих его. Молекулярные рецепторы (R) – это белки, специфически связывающие гормон и осуществляющие сопряжение связывания гормона с передачей сигнала (300-1200 а.к.).
Белок-рецептор должен обладать специфичностью по отношению к тому или иному гормону. Взаимодействие гормона с рецептором характеризуется эквимолярной стехиометрией (один гормон – один рецептор). Продукт взаимодействия гормона с рецептором носит название гормон-рецепторный комплекс (ГР-комплекс). Количество разных типов и подтипов рецепторов у одного гормона различно:
· 1 R – либерины, тропины, окситоцин, кальцитонин, паратирин, глюкагон, инсулин, кортизол, тестостерон;
· 2 R – ангиотензин II, эстрадиол, иодтиронины, холецистокинин, эндотелины;
· 3 R – антидиуретический гормон, галанин, γ-аминомасляная кислота;
· 4 R – гистамин, простагландин Е2;
· 9 R катехоламины;
· 11 R – глутамат;
· 13 R –cеротонин.
Наличие нескольких R одного гормона и их связь с разными сигнал-трансдукторными системами способствуют плейотропности, а часто обратимости и дуализму действия гормонов. Одна и та же клетка может быть чувствительна к 4-23 разным гормонам (особенно хемокинам и цитокинам), эффекты которых взаимно модулируются и интерферируют. Следует также отметить, что уровень гормона в крови может влиять на количество рецепторов в клетке (например, прогестерон уменьшает количество своих рецепторов и рецепторов для эстрогенов в матке).
Рецепторы гормонов в клетках-мишенях могут находиться либо на плазматической мембране вместе с другими мембранными белками, либо внутри клеток в различных ее компартментах (гиалоплазма, ядро, митохондрии и др.). Локализация рецепторов определяется степенью гидрофильности или гидрофобности гормонов. Гидрофильные гормоны имеют рецепторы на поверхности плазматической мембраны клетки-мишени, т.к. гидрофильные молекулы не могут проникать сквозь гидрофобный липидный бислой мембран. Напротив, гидрофобные гормоны, свободно преодолевающие мембраны, имеют внутриклеточные рецепторы. Мембранные рецепторы являются гликопротеинами, т.е. сложными белками, имеющими разветвленные олигосахаридные цепи в качестве небелкового компонента. Эти цепи наряду с олигосахаридными фрагментами мембранных гликолипидов участвуют в узнавании и специфическом связывании гормонов. Белковая часть молекулы мембранных рецепторов имеет достаточно мощный гидрофобный домен, позволяющий им распределяться в липидном бислое, как все интегральные или трансмембранные белки. При этом их гормонсвязывающий домен гидрофилен и располагается снаружи мембраны.
Рецепторы плазматической мембраны клеток подразделяются на суперсемейства:
1. 7-сегментные трансмембранные рецепторы (7-ТМС, G-protein-coupled receptors-GPCR). Это интегральные мембранные белки с 7 трансмембранными α-спиралями, соединенные гидрофильными внеклеточными и внутриклеточными петлями. Цитоплазматический домен этих рецепторов содержит центры связывания с G-белком (рис. 3).
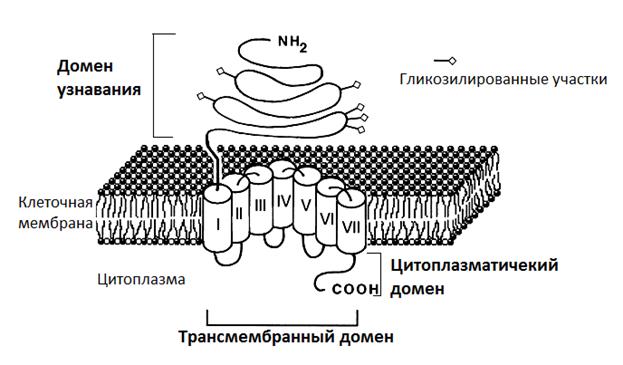
Рис. З. Структура 7-ТМС рецептора (https://studfiles.net/preview/6659447/page:2/).
2. 1-сегментные трансмембранные каталитические рецепторы (1-ТМС) – интегральные мембранные белки с 1 трансмембранным сегментом и глобулярными доменами на вне- и внутриклеточной поверхностях мембраны. Внеклеточный домен узнает и связывает лиганд-гормон, а внутриклеточный обладает каталитической активностью (гуанилатциклазной, тирозинкиназной и др.) и запускает реакцию образования внутриклеточных вторичных посредников.
3. Каналообразующие рецепторы – состоят из ассоциированных белковых субъединиц, каждая из которых содержит несколько трансмембранных сегментов. Эти рецепторы являются лигандозависимыми ионными каналами. В качестве лигандов, как правило, выступают нейромедиаторы.
Внутриклеточные рецепторы подразделяются на 2 класса:
Класс I – ядерные или цитоплазматические, без лиганда, связаны с белками теплового шока.
Класс II – ядерные, связаны с белками теплового шока.
Внутриклеточные рецепторы в целом гидрофильны и способны к диффузии в водной фазе, однако их гормонсвязывающий участок образован гидрофобными радикалами аминокислот, что важно для образования гидрофобных связей между гормоном и молекулой рецептора. Домен узнавания (гормонсвязыающий участок) служит для присоединения гормона и у внутриклеточных рецепторов он находится на С-конце полипептидной цепи рецептора. Домен связывания ДНК расположен в центральной части рецептора и содержит надвторичные структуры «цинковые пальцы» (рис. 4). В каждой такой структуре атом цинка связан с 4 остатками цистеина, при этом от остальной полипептидной цепи отделяется последовательность аминокислот, способная специфически взаимодействовать с определенными участками молекулы ДНК. Вариабельный домен расположен на N-конце полипептидной цепи внутриклеточного и служит для связывания с различными регуляторными белками, в том числе шаперонами.
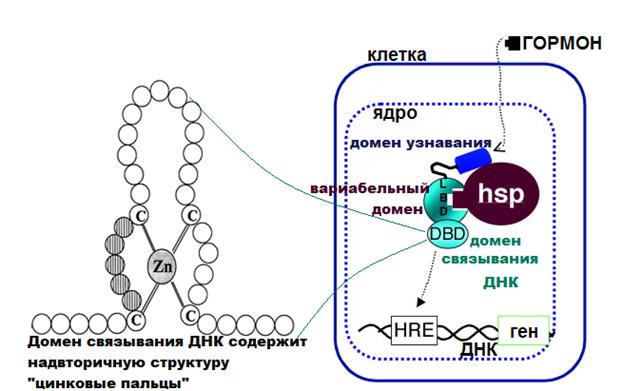
Рис.4. Структура внутриклеточного рецептора (https://megalektsii.ru/).
Дата: 2018-12-21, просмотров: 443.