Министерства здравоохранения Российской Федерации
Кафедра биохимии им. Г.Я. Городисской
Гормоны
Учебное пособие
Под редакцией Е.И. Ерлыкиной
Нижний Новгород
Г .
УДК 577.17 (075)
ББК 28.072
Г696
Составители:
профессор д.б.н. Л.М. Обухова, доценты к.м.н. П.П. Загоскин, к.б.н. Е.И. Кузьмина, старший преподаватель к.б.н. А.А. Анашкина.
Г696 Гормоны. Учебное пособие / под ред. д.б.н. проф. Е.И. Ерлыкиной. – Н. Новгород: 2018. – 39 с.
Учебное пособие составлено в соответствии с ФГОС по специальностям 31.05.01 «Лечебное дело», 31.05.02 «Педиатрия», 31.05.03 «Стоматология», 32.05.01 «Медико-профилактическое дело», 33.05.01 «Фармация» для студентов медицинских вузов.
Пособие предназначено для самостоятельной работы студентов при подготовке к практическим занятиям и коллоквиумам и должно оказать помощь в более глубоком освоении весьма сложного материала. Авторы старались избежать дублирования материала, который входит в программы ряда смежных медико-биологических и клинических дисциплин, заострив внимание на биохимических аспектах, касающихся химической классификации и молекулярного механизма действия гормонов.
Простота изложения вопросов фундаментальной биохимии, современных достижений в этой области знаний позволяет сделать это пособие востребованным не только студентами, но и аспирантами, ординаторами и врачами.
УДК 577.17 (075)
ББК 28.072
Рецензенты:
1. Зав. кафедрой биохимии и биотехнологии Института биологии и биомедицины ФГАОУ ВО «Национальный исследовательский Нижегородский государственный университет им. Н.И. Лобачевского», д.б.н., проф. А.С. Корягин.
2. Зав. кафедрой эндокринологии и внутренних болезней ФГБОУ ВО ПИМУ МЗ России, д.м.н., профессор Л.Г. Стронгин.
Печатается по решению….
© ФГБОУ ВО ПИМУ Минздрава России, 2018
Список сокращений
7-ТМС – 7-сегментные трансмембранные рецепторы
HRE – hormone response element
IRS – insulin receptor substrate
R – рецептор гормона
АКТГ – адренокортикотропный гормон
ГР-комплекс – гормон-рецепторный комплекс
ДАГ – диацилглицерол
ИФ3 – инозитол-3-фосфат
ЛО – липоксины
ЛТ – лейкотриены
ПГ – простагландины
СТГ – соматотропный гормон
Т3 – трийодтиронин
Т4 – тетрайодтиронин, тироксин
ТО – тромбоксаны
ТТГ – тиреотропный гормон
ФЛ А2 – фосфолипаза А2
Фн – фосфорная кислота (Н3РО4)
цАМФ – циклический аденозинмонофосфат
цГМФ – циклический гуанозинмонофосфат
ЦОГ – циклооксигеназа
Что такое гормоны?
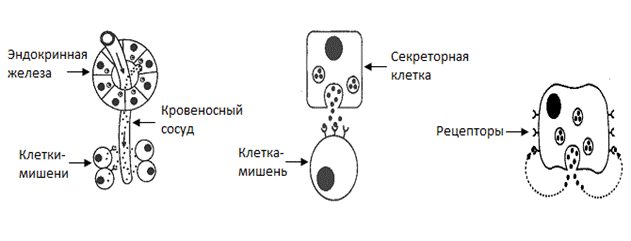
 Сравнительно недавно гормонами называли биологически активные вещества, образующиеся в специализированных (эндокринных) железах и регулирующие метаболизм и функции клеток других органов и тканей (клеток-мишеней) гуморальным путем (т.е. передаваясь через кровоток). Например, инсулин, вырабатываемый β-клетками островков Лангерганса поджелудочной железы, доставляется с помощью системы кровообращения к печени (гепатоциты рассматриваются как один из наиболее важных типов клеток-мишеней для инсулина) и активирует в ней синтез гликогена из глюкозы. Все гормоны – действительно биологически активные вещества. Этим термином обычно обозначают вещества, вызывающие выраженные физиологические эффекты в очень низких концентрациях (порядка 10-6-10-9 М и ниже). И все же, в настоящее время, приведенное выше определение понятия «гормоны» следует считать несколько устаревшим, так как многими научно-исследовательскими работами доказано, что гормоны могут синтезироваться далеко не только в специализированных эндокринных клетках, но и в самых обычных соматических клетках разных типов. Их регуляторный сигнал может передаваться вовсе не обязательно через кровь, а просто диффузионным путем от одной клетки к другой в пределах одной и той же клеточной популяции (паракринная регуляция) или даже в пределах одной и той же клетки от одной ее части к другой (аутокринная регуляция, рис. 1). Термин «гормон» (от греч. hormao – приводить в движение, побуждать) был впервые предложен Уильямом Бэйлиссом и Эрнестом Старлингом в 1905 году для секретина. Этот пептид образуется в клетках двенадцатиперстной кишки и воздействует на функции поджелудочной железы. Исторический парадокс состоит в том, что сложившееся после этого открытия определение понятия «гормон» в принципе нельзя было использовать для секретина, т.к. он не вырабатывается в какой-либо эндокринной железе и возможно даже обладает паракринным эффектом.
Сравнительно недавно гормонами называли биологически активные вещества, образующиеся в специализированных (эндокринных) железах и регулирующие метаболизм и функции клеток других органов и тканей (клеток-мишеней) гуморальным путем (т.е. передаваясь через кровоток). Например, инсулин, вырабатываемый β-клетками островков Лангерганса поджелудочной железы, доставляется с помощью системы кровообращения к печени (гепатоциты рассматриваются как один из наиболее важных типов клеток-мишеней для инсулина) и активирует в ней синтез гликогена из глюкозы. Все гормоны – действительно биологически активные вещества. Этим термином обычно обозначают вещества, вызывающие выраженные физиологические эффекты в очень низких концентрациях (порядка 10-6-10-9 М и ниже). И все же, в настоящее время, приведенное выше определение понятия «гормоны» следует считать несколько устаревшим, так как многими научно-исследовательскими работами доказано, что гормоны могут синтезироваться далеко не только в специализированных эндокринных клетках, но и в самых обычных соматических клетках разных типов. Их регуляторный сигнал может передаваться вовсе не обязательно через кровь, а просто диффузионным путем от одной клетки к другой в пределах одной и той же клеточной популяции (паракринная регуляция) или даже в пределах одной и той же клетки от одной ее части к другой (аутокринная регуляция, рис. 1). Термин «гормон» (от греч. hormao – приводить в движение, побуждать) был впервые предложен Уильямом Бэйлиссом и Эрнестом Старлингом в 1905 году для секретина. Этот пептид образуется в клетках двенадцатиперстной кишки и воздействует на функции поджелудочной железы. Исторический парадокс состоит в том, что сложившееся после этого открытия определение понятия «гормон» в принципе нельзя было использовать для секретина, т.к. он не вырабатывается в какой-либо эндокринной железе и возможно даже обладает паракринным эффектом.
 Телекринная Паракринная Аутокринная
Телекринная Паракринная Аутокринная
регуляция регуляция регуляция
(например – (например – (например –
действие действие действие
«классических» желудочно- простагландинов)
гормонов) кишечных
гормонов)
Рис. 1. Типы гормональной регуляции.
Таким образом, в современном толковании определение понятия «гормоны» может быть сведено к следующему:
Гормоны – биологически активные вещества, образующиеся в одной части организма и регулирующие метаболизм и функции в другой его части. При этом надо иметь ввиду, что под «частями организма» могут подразумеваться даже разные структурные элементы одной и той же клетки (аутокринная регуляция) или разные клетки в одной и той же клеточной популяции (паракринная регуляция).
Несомненно, что такое определение включает и всю группу «классических» гормонов, передающих регуляторный сигнал гуморальным путем (телекринная регуляция).
Классификация гормонов.
Рис. 2 Схема иерархической организации нейроэндокринной системы и
Таблица 2
Таблица 3
Примеры некоторых энхансерных участков ДНК, ответственных
Таблица 4
Рис. 7. Схема аденилатцикалазного механизма передачи гормонального сигнала в клетку.
В неактивном состоянии протеинкиназа А – олигомерный белок, состоящий из 4 субъединиц: двух регуляторных и двух каталитических. Присоединение цАМФ к регуляторным субъединицам ведет к конформационным изменениям всего белка. Каталитические субъединицы становятся некомплементарными регуляторным и отделяются (диссоциируют) от них. В диссоциированной форме каталитические субъединицы протеинкиназы А активны и катализируют реакции фосфорилирования белков по серину и треонину. Каталитические субъединицы специфичны к белкам-субстратам. Присоединение остатка фосфорной кислоты от АТФ к уже имеющемуся в клетке белку ведет к изменению его конформации, а, следовательно, и активности, если этот белок является ферментом. Напомним, что такая регуляция каталитической активности у ферментов называется регуляцией путем фосфорилирования и дефосфорилирования.
30 гормонов через 45R увеличивают концентрацию цАМФ (кортикотропин, глюкагон, паратирин, кальцитонин, катехоламины (β-рецепторы), гистамин (Н2-рецепторы) и др.).
31 гормон снижают концентрацию цАМФ (соматостатин, мелатонин, опиоиды, хемокины, катехоламины (α2-рецепторы), γ-аминомасляная кислота и др.).
цАМФ также регулирует геномные процессы, участвует в патогенезе холеры, коклюша и муковисцидоза.
После выполнения гормоном его регуляторной функции необходимо прекратить передачу гормонального сигнала. Для этого на каждом этапе существует система остановки передачи. Так, фермент фосфодиэстераза дециклизует цАМФ (рис. 8), а фосфопротеинфосфатазы дефосфорилируют фосфорилированные белки.
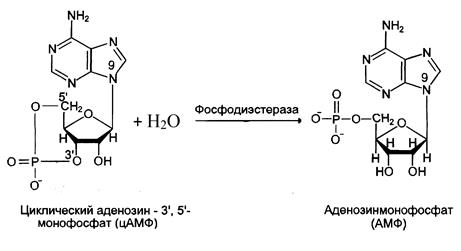
Рис. 8. Дециклизация цАМФ.
Гуанилатциклазный механизм включает в себя активацию мембранного фермента гуанилатциклазы различными гормонами или цитокинами. Появление в цитозоле клеток-мишеней циклического гуанозинмонофосфата (цГМФ) стимулирует активность протеинкиназы G (цГМФ-зависимой протеинкиназы), которая, в свою очередь, фосфорилирует специфические клеточные белки. Например, предсердный натрийуретический гормон, действуя подобным образом, способствует фосфорилированию белков ионных каналов почечных канальцев, результатом чего является повышение экскреции ионов Na+. Некоторые вазодилататоры, например оксид азота ( NO), лекарственные нитраты (нитроглицерин) и др. используют гуанилатциклазный механизм для расслабления гладкой мускулатуры сосудов, результатом чего является расширение просвета сосудов и улучшение кровотока через орган (например, миокард). Гуанилатциклазный механизм используют также гормоны задней доли гипофиза (окситоцин) и щитовидной железы (кальцитонин). Регуляторные эффекты этих гормонов указаны в табл 2.
При использовании инозитол-3-фосфатной системы передачи гормонального сигнала в клетку происходят следующие события:
Образование ГР-комплекса вызывает конформационные изменения, передающиеся на G-белок, при этом α-субъединица G-белка становится некомплементарна ГДФ и увеличивает комплементарность к ГТФ. Она диссоциирует от белка и активирует фосфолипазу С. Фосфолипаза С – поверхностный белок. Она катализирует гидролиз фосфотидилинозитол-4,5-дифосфата до диацилглицерола (ДАГ) и инозитол-1,4,5-трифосфта (ИФ3), которые являются промежуточными посредниками передачи гормонального сигнала в клетку. ИФ3 - водорастворимое вещество, стимулирующее высвобождение ионов Са2+ из эндоплазматического ретикулума. Са2+ связывается с белком кальмодулином, комплекс Са2+-кальмодулин активирует Са2+-кальмодулин-зависимую протеинкиназу. Ионы Са2+ также могут связываться с другой протеинкиназой – протеинкиназой С – и активировать ее. Кроме того, регуляторный домен этого фермента имеет участок связывания с ДАГ. ДАГ – гидрофобное вещество и всегда находится в составе липидного бислоя мембраны. При контакте с ДАГ протеинкиназа С повышает свое сродство к ионам Са. Активная каталитическая субъединица осуществляет реакцию фосфорилирования белков по серину и треонину, что ведет к изменению их конформации и активности. В результате меняется метаболизм клетки-мишени и формируется специфический клеточный ответ на регуляторное действие гормона (рис. 9).
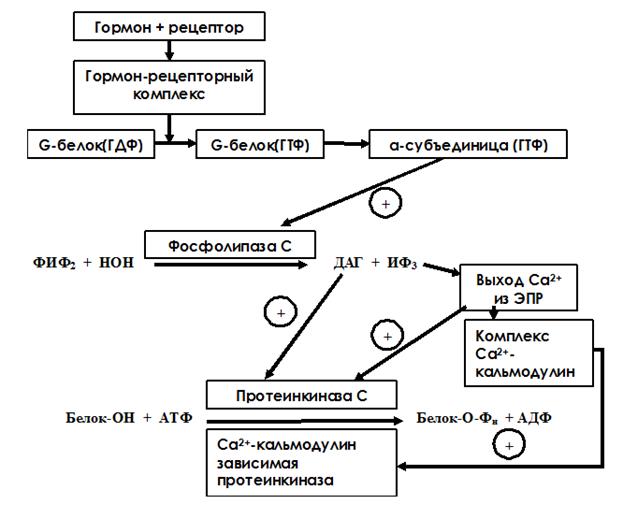
Рис. 9. Схема инозитол-3-фосфатного механизма передачи гормонального сигнала в клетку.
Такой тип передачи гормонального сигнала в клетку характерен для вазопрессина (рецептор V1) и адреналина (рецептор α1).
Подавление сигнала осуществляется за счет закачивания Ca2+ в ЭПР и образования фосфатидилинозитолдифосфата из ДАГ и ИФ3. Фосфопротеины дефосфорилируются фосфопротеинфосфатазой.
Еще одним примером мембранного механизма передачи гормонального сигнала в клетку является механизм действия инсулина. Рецептор инсулина состоит из четырех субъединиц:
- 2 α-субъединицы лежат на внешней стороне мембраны, образуют центр связывания с инсулином;
- 2 β-субъединицы – интегральные белки, пронизывают бислой, на внутриклеточных доменах имеют активный центр, способны катализировать реакцию фосфорилирования белков по тирозину, то есть обладают каталитической активностью (рис. 10).
Присоединение инсулина к α-субъединицам ведет в конечном итоге к изменению конформации всего рецептора и активных центров на β-субъединицах, происходит их активация. Сначала β-субъединица фосфорилирует по остатку тирозина соседнюю β-субъединицу (аутофосфорилирование). Это приводит к конформационным изменениям активных центров и изменению их субстратной специфичности. Субстратами становятся внутриклеточные белки – субстраты инсулинового рецептора – IRS (от англ. insulin receptor substrate). Фосфорилированные по тирозину IRS-белки активируют фосфопротеинфосфатазу.
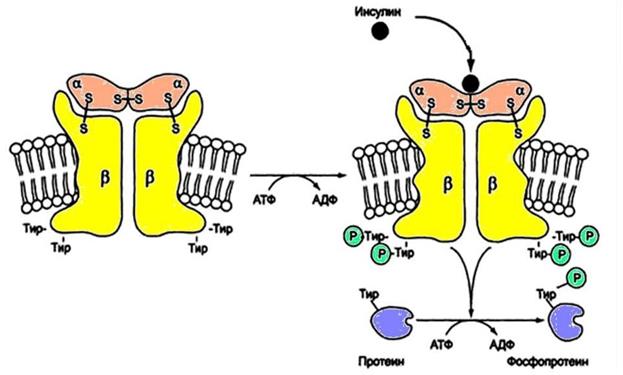
Рис. 10. Инсулиновый рецептор. Тирозинкиназный механизм передачи сигнала в клетку-мишень (https://en.ppt-online.org).
Фосфопротеинфосфатаза осуществляет гидролитическое отщепление остатка фосфорной кислоты от фосфорилированных по серину и треонину белков, участвующих в регуляции клеточного метаболизма:
Белок-О-Фн + Н2О → Белок-ОН + Фн.
Кроме того, IRS-белки активируют фосфодиэстеразу, которая дециклизует цАМФ.
Таким образом, инсулин действует как антагонист глюкагона и адреналина, выполняя функцию снижения концентрации глюкозы в крови.
Дефосфорилирование рецептора тирозиновой фосфопротеинфосфатазой возвращает его в неактивное состояние.
Для инсулинового рецептора известны и другие варианты передачи гормонального сигнала в клетку. Например, ГР-комплекс вместе с участком мембраны способен проникать (интернализация) в цитоплазму и в ядро, связываться с ДНК и изменять активность синтеза некоторых белков, что демонстрирует возможность мембранных гормонов действовать аналогично цитоплазматическим.
Таблица 5
Простагландины
Тромбоксаны
Лейкотриены
Как следует из данных табл. 5, спектр биологических эффектов эйкозаноидов весьма разнообразен, что открывает широкие перспективы их использования в практической медицине, в частности для регулирования тонуса гладкой мускулатуры сосудов, бронхов, матки, коррекции нарушенных функций клеток крови, процесса свертывания крови, управления родовой деятельностью, профилактики и лечения аллергических состояний и т. д.
Рис. 11. Закладка эндокринной системы в эмбриогенезе (Данилова ЛА, редактор. Возрастная биохимия. Учебное пособие. СПб: Сотис; 2007.)
Для периода новорожденности также характерно существенное влияние гормонов матери, поступивших как трансплацентарно, так и с грудным молоком. В первые 3 часа после родов в крови новорожденных присутствуют гормоны матери, плацентарные гормоны (эстриол, кортизол) и гормоны плода (Т3, Т4, ТТГ, соматотропный гормон).
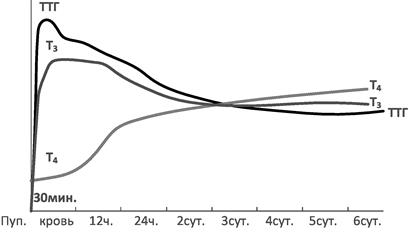
Рис. 12. Изменение уровней секреции ТТГ и гормонов щитовидной железы в первые дни жизни (Данилова ЛА, редактор. Возрастная биохимия. Учебное пособие. СПб: Сотис; 2007.)
Помимо резкого повышения уровня ТТГ, в течение первого получаса жизни отмечается также глюкагоновый всплеск, обеспечивающий, наряду с адреналиновым, эффективный гликогенолиз и липолиз в первые минуты и часы жизни. К середине-концу первой недели жизни в крови ребенка увеличивается концентрация соматотропного гормона, альдостерона (а значит, и ренина), тестостерона, паратгормона, фолликулостимулирующего гормона.
Считается, что транзиторное повышение активности надпочечников и щитовидной железы играет важнейшую роль в метаболической адаптации новорожденного к условиям внеутробной жизни. Если в первые минуты и часы жизни решающая роль в этом процессе принадлежит гормонам фетоплацентарного комплекса и матери (кортизол, кортизон, кортикостерон, эстрагены), то уже к 3-6 часу жизни особо важную роль играют собственные гормоны щитовидной и других желез. Постепенность выведения фетальных, плацентарных, материнских гормонов обеспечивает и постепенность адаптации новорожденного к внеутробным условиям жизни. Решающая роль собственных эндокринных желез (надпочечников, гипофиза и щитовидной железы) в метаболической и других видах адаптации новорожденного особенно велика в середине-конце первой недели жизни.
У новорожденных в первые дни жизни отмечается высокий уровень тропных гормонов - АКТГ, ТТГ, соматотропного гормона.
Соматотропин (гормон роста, соматотропный гормон, СТГ) у детей стимулирует рост трубчатых костей. Линейный рост ребенка до периода полового созревания определяется соматотропным гормоном. Является гормоном с выраженным анаболическим действием: активирует синтез белка, рост мышечной массы, внутренних органов. Также принимает участие в регуляции углеводного обмена - вызывает повышение уровня глюкозы в крови. По химической природе соматотропин - простой белок. СТГ синтезируется в передней доле гипофиза. Секреция соматотропина максимальна ночью. Наибольшая концентрация соматотропина в крови наблюдается на 4-6 месяц внутриутробного развития (примерно в 100 раз выше, чем у взрослого). С возрастом секреция СТГ постепенно снижается.
Адренокортикотропный гормон (кортикотропин) контролирует синтез и секрецию гормонов коры надпочечников (глюкокортикоидов, минералокортикоидов, прогестерона, андрогенов, эстрогенов). АКТГ является пептидом, синтезируется в передней доле гипофиза.
Тиреотропный гормон (тиреотропин, тиротропин, ТТГ) стимулирует синтез и секрецию йодтиронинов (тироксин, трийодтиронин) в щитовидной железе. ТТГ является гликопротеином, синтезируется в передней доле гипофиза. Концентрация ТТГ у новорожденных и детей первых 2 лет жизни в несколько раз выше, чем у взрослых. Секреция с возрастом уменьшается, приближаясь к показателям взрослого.
Тиреоидные гормоны синтезируются тиреоидными фолликулами щитовидной железы, являются йодированными производными аминокислоты тирозина и отличаются числом атомов йода. Тиреоидные гормоны регулируют скорость основного обмена, рост и дифференцировку тканей, обмен белков, углеводов, липидов, водно-солевой обмен, повышают утилизацию АТФ и поглощение кислорода тканями. Необходимы для нормального созревания скелета плода и после рождения совместно с гормоном роста влияют на рост костной ткани.
Физиологический гипертиреоз новорожденных наблюдается на фоне сниженной чувствительности рецепторов к тиреоидным гормонам в период новорожденности и раннего детства и высокой концентрации тироксинсвязывающего глобулина. Во время вскармливания отмечается нарастание интенсивности функционирования щитовидной железы. Для этого возраста характерно активное поглощение йода щитовидной железой и снижение уровня йода в сыворотке крови. В период молочных зубов (от 1 года до 6-7 лет) наибольшее влияние на организм оказывают гормоны щитовидной железы. С возрастом секреция тиреоидных гормонов уменьшается, приближаясь к показателям взрослого.
Поскольку глюкоза поступает к плоду с постоянной скоростью, инсулиносекреция не играет большой роли в обмене глюкозы во внутриутробном периоде. У новорожденных, особенно недоношенных детей концентрация инсулина в крови очень низка, не установлена зависимость уровня сахара от активности инсулина. Инсулиносекреция становится устойчивой только к 6 годам жизни.
Секреция глюкагона (повышает уровень глюкозы в крови) развивается с рождения и его активность в плазме крови сопоставима с активностью взрослых. В первые часы после рождения активность глюкагона в плазме несколько нарастает, но быстро снижается и вновь повышается на 2-3 сутки, когда происходит восстановление уровня глюкозы после периода гипергликемии.
Содержание
Стр.
Список сокращений 3
1 Что такое гормоны? 4
2 Классификация гормонов. 5
2.1 Классификация гормонов по химическому строению. 5
2.2 Классификация гормонов по месту выработки. 5
2.3 Классификация гормонов по биологической активности. 7
3 Клетки-мишени и клеточные рецепторы гормонов. 8
4 Синтез гормонов. Транспорт гормонов. 10
5 Интактивация и выведение продуктов биотрансформации гормонов из организма 11
6 Характеристика регуляторных эффектов гормонов 12
7 Механизм передачи сигнала в клетку-мишень. 16
7.1Внутриклеточный механизм действия гормонов. 16
7.2 Мембранный механизм действия гормонов. 18
8 Эйкозаноиды и их роль в регуляции метаболизма и физиологических функций 23
9 Патологические состояния, связанные с гипер- и гипопродукцией гормонов. Заместительная терапия при гипопродукции гормонов. 25
10 Гормоны как лекарственные препараты. 26
11 Особенности гормональной регуляции в эмбриональном периоде и у новорожденных. 28
12 Гормоны, регулирующие минерализацию костной ткани. 30
Тестовые задания 32
Ответы на тестовые задания 37
Список основной рекомендуемой литературы 38
Список дополнительной рекомендуемой литературы 39
Министерства здравоохранения Российской Федерации
Кафедра биохимии им. Г.Я. Городисской
Гормоны
Учебное пособие
Дата: 2018-12-21, просмотров: 486.