Под высшей организацией ДНК понимают организацию ДНК внутри ядер клеток млекопитающих. Двойная спираль ДНК дважды обернута вокруг гистонов (октомеры). Эта суперспираль формирует центральную часть (10 нм в диаметре), называемую нуклеосомами:
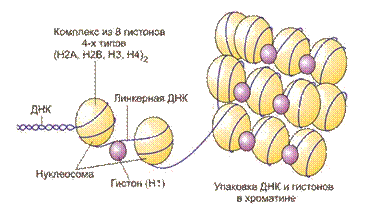
Нуклеосома состоит из участка ДНК длиной около 146 нуклеотидных пар и 8 молекул гистонов четырех видов, которые в количестве по два каждого вида образуют комплекс – нуклеосомный кор. ДНК и белки удерживаются друг с другом за счет ионных связей. Линкерный участок ДНК связан с гистоном Н1.
Основная функция нуклеосомы - собрать и стабилизировать ДНК. Комплекс гистона и ДНК называется хроматином. Длина ДНК очень большая, она содержит 7 × 109 пар нуклеотидов. Состояние хроматина меняется в зависимости от клеточного цикла. В фазе покоя он распределен по всему ядру. В фазе деления образуются компактные частицы - хромосомы.
Когда клетка делится, дочерние клетки должны получить идентичную копию генетической информации от материнской клетки. Копирование (удвоение) ДНК происходит в течение S-фазы, предшествующей клеточное деление. Процесс образования идентичной копии, то есть, удвоение ДНК, называется репликацией.
Наиболее последовательно изучен процесс репликации для прокариот, и его изучение позволяет лучше понять процесс репликации эукариот и его особенности, которые тщательно изучаются и в наши дни.
Биосинтез ДНК (репликация)
Репликация - матричный процесс. Во время репликации каждая из двух цепей ДНК служит матрицей для образования новой цепи. Субстратами и источниками энергии для синтеза ДНК являются дезоксирибонуклеозидтрифосфаты (дНТФ: дАТФ, дГТФ, дЦТФ, дТТФ).
Процесс включает следующие основные этапы.
I. Формирование репликативной вилки.
II. Синтез новых цепей ДНК.
| III. Исключение праймеров. Завершение формирования отстающей цепи ДНК. |
I. Формирование репликативной вилки (рис. 5) идет при участии:
ДНК-топоизомеразы, которая является «обратимой нуклеазой». Сначала фермент разрывает 3'-,5'-фосфодиэфирную связь в одной из цепей ДНК;
ДНК-хеликаз - ДНК-зависимых АТФаз, использующих энергию АТФ для расплетения двойной спирали ДНК;
SSB (single strand binding) -белков, связывающихся с одноцепочечными участками ДНК. Эти белки, не закрывая оснований, предотвращают повторное комплементарное скручивание матричных цепей и образование шпилек.
II. Синтез новых цепей ДНК.
На этой стадии дочерние нити ДНК образуются на обеих нитях материнской ДНК. Процесс катализирует несколько ДНК-полимераз, которые синтезируют полинуклеотидные цепи из дНТФ: дАТФ, дГТФ, дТТФ и дЦТФ в направлении от 5'- к 3'-концу антипараллельно матрице, имеющей направление от 3'- к 5'-концу.
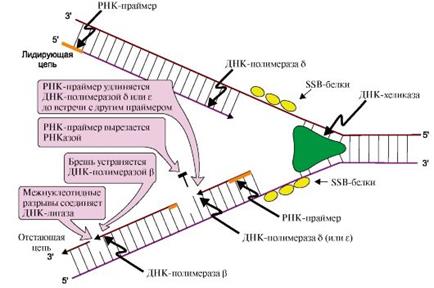
Рис.: Синтез новых цепей в области репликативной вилки
Новые цепи синтезируются по-разному. На матрице ДНК с направлением от 3'- к 5'-концу цепь растет непрерывно по ходу движения репликативной вилки и называется лидирующей. На матрице с направлением от 3'- к 5'-концу вторая цепь синтезируется против движения репликативной вилки в виде коротких отрезков - фрагментов Оказаки. Рост этой цепи начинается только тогда, когда на матрице ДНК появляется одноцепочечный участок длиной около 200 нуклеотидов, поэтому ее называют запаздывающей или отстающей.
Лидирующая нить растет непрерывно, а отстающая - в виде фрагментов Оказаки, каждый из которых включает:
- РНК-праймер (~10 нуклеотидов);
- участок ДНК, примерно равный длине цепи из 150 нуклеотидов
ДНК-полимеразы δ, β и ε не способны инициировать синтез новых цепей ДНК, они могут лишь удлинять имеющуюся нуклеотидную цепь. Синтез лидирующей и отстающей нитей начинается с образования затравки или праймера-олигорибонуклеотида (РНК), включающего около 10 мононуклеотидов. Его образование катализирует праймаза -субъединица ДНК-полимеразы α. Далее этот же фермент, используя в качестве субстратов дНТФ, переключается на образование ДНК и включает во вновь синтезируемую нить 20-50 дезоксирибонуклеотидов, после чего заменяется другими ДНК-полимеразами. Синтез лидирующей цепи продолжает ДНК-полимераза δ, а отстающей - ДНК-полимераза δ или ε. Оба фермента, помимо, полимеразной обладают еще и экзонуклеазной активностью. В ходе синтеза они могут исправлять допущенную ошибку и отщеплять неправильно включенный нуклеотид, что обеспечивает высокую точность синтеза ДНК.
III. Исключение праймеров. Завершение формирования отстающей цепи ДНК
В отстающей нити праймер удаляется эндонуклеазой или РНКазой. Затем ДНК-полимераза β заполняет образованную «брешь», присоединяя по принципу комплементарности матрице дезоксирибонуклеотиды в количестве, равном вырезанному праймеру. Связывание 3'-ОН-группы одного фрагмента с 5'-фосфатом предыдущего фрагмента и образование фосфодиэфирной связи катализирует ДНК-лигаза. Фермент, используя энергию АТФ, из множества фрагментов Оказаки образует непрерывную цепь ДНК.
2. Результатом процесса является образование дочерних цепей, комплементарных и антипараллельных нитям материнской ДНК.
Репарация ДНК
Репарация – особая функция клеток, заключающаяся в способности исправлять химические повреждения и разрывы в молекулах ДНК, поврежденной при нормальном биосинтезе ДНК в клетке или в результате воздействия физических или химических агентов. Осуществляется специальными ферментными системами клетки (рис. 8). Ряд наследственных болезней, к примеру, пигментная ксеродерма, связан с нарушениями системы репарации.

Рис. 8. Репарация эукариотической ДНК
Дата: 2018-12-21, просмотров: 445.