Для представителей класса Насекомые (Insecta) характерно разделение тела на голову, грудь и брюш- ко. Голова несет простые или фасеточные глаза, пару членистых усиков и пару щупиков. Ротовой аппарат грызущего, сосущего, лижущего, колюще-сосущего и других типов состоит из пары верхних и пары нижних челюстей, верхней и нижней губы.
Грудь состоит из трех сегментов. С брюшной сто- роны каждый сегмент несет пару конечностей раз- личного типа. Со спинной стороны предпоследний и последний сегменты несут по паре крыльев. Первая пара крыльев может быть видоизменена в жесткие
надкрылья (отряд жесткокрылые), вторая — превра- титься в жужжальца (отряд двукрылые). Крылья мо- гут отсутствовать (первичная или вторичная бескры- лость).
Брюшко членистое, конечностей не имеет. Дыхание трахейное. Выделительная система представлена маль- пигиевыми сосудами (слепо заканчивающиеся вырос- ты кишечника на границе средней и задней кишки) и жировым телом («почка накопления»)
Развитие с полным (яйцо — личинка — куколка — имаго) или с неполным (яйцо — личинка — имаго) пре- вращением.
100 МЕДИЦИНСКАЯ ПАРАЗИТОЛОГИЯ НАСЕКОМЫЕ — КОМПОНЕНТЫ ГНУСА
Гнус — совокупность кровососущих насекомых от- ряда Двукрылые.
Heleidae), мошки (сем. Simuliidae), комары, слепни (сем. Tabanidae) — летающие кровососущие насеко- мые, компоненты гнуса.
Москиты (сем. Phlebotomidae), мокрецы (сем. МОСКИТЫ
Географическое распространение
Страны с жарким и теплым климатом.
Морфология
Мелкие кровососущие летающие насекомые длиной 1,5–3 мм (рис. 43.1, а, б). Голова вытянутая, несет хо- боток, усики, щупики. Тело имеет форму равнобедрен- ного треугольника, конечности по длине в 2–3 раза превышают туловище. Крылья относительно узкие, за- остренные к вершине, задняя пара крыльев редуциро- вана в жужжальца. Полет «прыгающий».
Голова, туловище и конечности покрыты густыми светлыми или темно-коричневыми волосками.
Жизненный цикл
Кровью питаются только самки. Нападают вече- ром и ночью, питаются на животных и человеке. Яйца откладывают 2–3 раза в течение жизни во влажных, за- щищенных от солнца местах. Развитие с полным мета- морфозом, при температуре 28 °С весь цикл развития занимает 35–45 дней. Продолжительность жизни около месяца, за это время проходит 2–3 гонотрофических цикла.
Медицинское значение
Переносчики лейшманиоза (рис. 43.2), флеботом- ной лихорадки, бартонеллеза.
Географическое распространение
Повсеместно, кроме высокогорных районов и тундры.
Морфология
Мелкие летающие насекомые размером 1–2 мм, внешне похожие на комаров. Хоботок примерно равен длине головы, голова несет пару щупиков и усиков (рис. 43.1, в).
Жизненный цикл
Кровью питаются только самки, нападающие ут- ром и вечером. Развитие с полным метаморфозом. Вы-
плаживаются преимущественно в небольших стоячих водоёмах, влажной почве. Зимуют обычно личинки.
Медицинское значение
Укусы мокрецов очень болезненны. На месте укуса образуется красноватая папула.
Могут служить переносчиками туляремии, вируса лимфоцитарного хориоменингита, японского энцефа- лита. Специфический переносчик и промежуточный хо- зяин филярии акантхейлонемы (Acanthcheilonema perstans).
Географическое распространение
Повсеместно.
Морфология
Мелкие летающие насекомые размером 1–3 мм, по- хожие внешне на мух. Тело короткое, черного или чер- но-коричневого цвета, крылья широкие, без пятен. Хо- боток короткий, толстый (рис. 43.1, е, ж; 43.3).
Жизненный цикл
Кровососущие только самки. Активны днем, с ут- ренним и вечерним пиком активности.
Яйца откладывают на водные предметы, личинки червеобразные, могут развиваться только в проточной воде. Зимуют яйца и личинки.
Медицинское значение
Укус мошек болезнен. В связи с наличием в слюне отравляющих веществ при массовых укусах могут быть симптомы общей интоксикации.
Могут передавать возбудителя туляремии, являют- ся специфическими переносчиками и промежуточными хозяевами филярии онхоцерка.
Географическое распространение
Повсеместно
Морфология
Крупные кровососущие мухи размером 1–3 см. Голо- ва крупная, несет большие ярко окрашенные глаз. Ро- товой аппарат режуще-лижущий. Крылья широкие, про- зрачные или с темным рисунком (рис. 43.1, в, д; 43.4).
Жизненный цикл
Кровососущие только самки. Нападают днем, осо- бенно вблизи водоемов. Питаются преимущественно на
крупных млекопитающих. Яйца откладывают на стеб- ли и листья растений вблизи водоёмов. Личинки живут в иле на дне водоемов, во влажной почве, на водных растениях.
Медицинское значение
Укусы слепней очень болезненны, так как их слю- на содержит токсические вещества.
Могут быть переносчиками сибирской язвы, туляре- мии, специфические переносчики и промежуточные хо- зяева филярии лоа-лоа.
Экзаменационный билет №8
1. Экспрессия генов в процессе биосинтеза белка. Регуляция экспрессии генов у прокариот и эукариот. Экспрессия генов — это процесс, в ходе которого наследственная информация от гена (последовательности нуклеотидов ДНК) преобразуется в функциональный продукт — РНК или белок. Экспрессия генов может регулироваться на всех стадиях процесса: и во время транскрипции, и во время трансляции, и на стадии посттрансляционных модификаций белков.
Регуляция экспрессии генов позволяет клеткам контролировать собственную структуру и функцию и является основой дифференцировки клеток, морфогенеза и адаптации.
ДНК сама по себе представляет инертную молекулу и должна быть таковой по своей природе. Поэтому утверждение «жизнь — это ДНК» следует воспринимать очень осторожно. Для того чтобы информация, заложенная в ДНК, превратилась в жизненные функции, она должна быть превращена в действия, которые по частям представлены активностью белков-ферментов, а в полной мере — размножающимся организмом.
Переход представляет собой экспрессию генетической информации и осуществляется в два этапа. На первом действует аппарат РНК, включающий транскрипцию с ДНК на РНК с помощью РНК-полимеразы и трансляцию с РНК(второй этап), с помощью рибосомы, в белки. Именно эти последние и используются для образования как компонентов клетки, в том числе клеточных структур, так и ферментов. Действие РНК-полимеразы основано нa матричном копировании необходимого участка ДНК (гена или группы связанных генов — оперона) в однонитевую нить матричной РНК (мРНК), которая переносится затем к рибосоме, точнее, рибосомы нанизываются на нить мРНК с образованием полирибосомы. Синтез белка осуществляется путем присоединения в рибосоме молекулы транспортной РНК (тРНК) с аминокислотой к соответствующему участку на нити мРНК с образованием полипептидной цепи, соответствующей последовательности нуклеотидов мРНК.
Между аминокислотами и основаниями существует «генетический код», в котором каждой аминокислоте соответствуют кодоны, содержащие три нуклеотида.
РНК синтезируется на матрице ДНК посредством фермента РНК-полимеразы . РНК-полимераза состоит из четырех субъединиц и о-фактора, обеспечивающего распознавание участка старта. Связывание начинается с участка, называемого промотором (рис. ). Промотор может быть сильным и слабым. Сильный промотор инициирует синтез иРНК часто, слабый - гораздо реже. С другой стороны, промотор может быть регулируемым и нерегулируемым. Двойная нить (дуплекс) ДНК в этом месте расплетается, и считывается лишь одна нить. Синтез мРНК требует затраты одного эквивалента АТФ на нуклеотид. Промотор может начинать отдельный ген или же комбинацию генов для ферментов, объединенных общей функцией, — оперон. В конце гена или оперона располагается стоп-сигнал, позволяющий РНК-полимеразе отделиться.
Нить ДНК бактерий в этот момент напоминает «ламповый ерш» с висящими на ней мРНК, и это состояние можно увидеть на электронных фотографиях, где нити нуклеиновых кислот, благодаря фосфатам, видны как темные участки. На нити мРНК, еще не отделившейся от ДНК, может уже начинаться синтез белка благодаря присоединению к ней рибосом.
ДНК эукариот расположена в ядре. И требуется решения проблем транспорта мРНК внутри компартмента-лизованной клетки довольно больших размеров. У эукариот гены состоят из кодирующих участков, экзонов, и разделены некодиру-ющими — интронами. Образованная в ядре мРНК претерпевает процессинг (созревание), при котором, в частности, из нее удаляются некодирующие участки, и лишь затем она поступает для синтеза белка в рибосомы. Все это обусловливает значительное усложнение элементарного процесса.
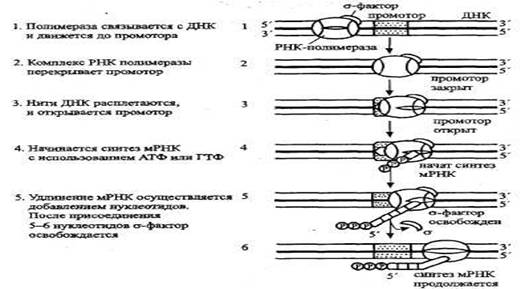
Рис. Действие РНК-полимеразы
Особенности Регуляция экспрессии генов эукариот:
Особенностью прокариот является транскрибирование мРНК со всех структурных генов оперона в виде одного полицистронного транскрипта, с которого в дальнейшем синтезируются от
дельные пептиды. Экспрессия генов у прокариот регулируется главным образом на уровне транскрипции. Роль сигнальных веществ для запуска транскрипции играют молекулы-эффекторы , представляющие собой низкомолекулярные соединения, которые являются либо субстратом для фермента, либо продуктом ферментативной деятельности соответственно. Индукция и репрессия представляют собой разные стороны одного и того же явления. Малые молекулы, индуцирующие образование ферментов, способных метаболизировать их, называются индукторами . Те же, которые предотвращают образование ферментов, способных синтезировать их, - корепрессорами .
Молекулы-эффекторы не могут вступать в прямое взаимодействие с ДНК, посредником для них служит специальный регуляторный белок . Регуляторный белок, который связывается с ДНК в отсутствии индуктора, называется репрессором .
Особенности Регуляция экспрессии генов эукариот:
У эукариотических организмов механизм регуляции транскрипции гораздо более сложен. В результате клонирования генов эукариот обнаружены специфические последовательности, принимающие участие в транскрипции и трансляции.
Для эукариотической клетки характерно:
1. Наличие интронов и экзонов в молекуле ДНК.
2. Созревание и-РНК - вырезание интронов и сшивка экзонов.
3. Наличие регуляторных элементов, регулирующих транскрипцию, таких как:
а) промоторы - 3 вида, на каждый из которых садится специфическая полимераза б) модуляторы - последовательности ДНК, усиливающие уровень транскрипции; в) (энхансеры) усилители - последовательности, усиливающие уровень транскрипции и действующие независимо от своего положения относительно кодирующей части гена и состояния начальной точки синтеза РНК; г) терминаторы - специфические последовательности, прекращающие и трансляцию, и транскрипцию.
2. Эмбриональная индукция. Дифференциация и интеграция в развитии. Эмбриональная индукция — это взаимодействие частей развивающегося зародыша, при котором один участок зародыша влияет на судьбу другого участка. Явление эмбриональной индукции с начала XX в. изучает экспериментальная эмбриология.
Классическими считают опыты немецкого ученого Г. Шпемана и его сотрудников (1924) на зародышах амфибий. Для того чтобы иметь возможность проследить за судьбой клеток определенного участка зародыша, Шпеман использовал два вида тритонов: тритона гребенчатого, яйца которого лишены пигмента и потому имеют белый цвет, и тритона полосатого, яйца которого благодаря пигменту имеют желто-серый цвет.
Один из опытов заключается в следующем: кусочек зародыша из области дорсальной губы бластопора на стадии гаструлы тритона гребенчатого пересаживают на боковую или вентральную сторону гаструлы тритона полосатого (рис. 8.8). В месте пересадки происходит развитие нервной трубки, хорды и других органов. Развитие может достичь довольно продвинутых стадий с образованием дополнительного зародыша на боковой или вентральной стороне зародыша реципиента. Дополнительный зародыш содержит в основном клетки зародыша реципиента, но светлые клетки зародыша-донора тоже обнаруживаются в составе различных органов.
Из этого и подобных опытов следует несколько выводов. Во-первых, участок, взятый из спинной губы бластопора, способен направлять или даже переключать развитие того материала, который находится вокруг него, на определенный путь развития. Он как бы организует, или индуцирует, развитие зародыша как в обычном, так и в нетипичном месте. Во-вторых, боковая и брюшная стороны гаструлы обладают более широкими потенциями к развитию, нежели их презумптивное (предполагаемое) проспективное направление, так как вместо обычной поверхности тела в условиях эксперимента там образуется целый зародыш. В-третьих, достаточно точное строение новообразованных органов в месте пересадки указывает на эмбриональную регуляцию. Это означает, что фактор целостности организма приводит к достижению хорошего конечного результата из нетипичных клеток в нетипичном месте, как бы управляя процессом, регулируя его в целях достижения этого результата.
Дата: 2018-09-13, просмотров: 2021.