Аллельные гены – гены, расположенные в одном и том же локусе (месте) гомологичных (парных) хромосом и отвечающие за развитие альтернативных признаков. Виды взаимодействия:
1 – полное доминирование
2 – неполное доминирование
3 – сверхдоминирование
4 – кодоминирование
Полное доминирование – такое взаимодействие, при котором проявление доминантного аллеля А не зависит от присутствия аллеля а, а проявление рецессивного аллеля возможно лишь в гомозиготном состоянии. Иначе говоря, ген А полностью скрывает ген а. Пример: ген карих глаз полностью скрывает ген голубых глаз.
Неполное доминирование – такое взаимодействие, при котором доминантный ген неполностью скрывает рецессивный, один признак как бы «разбавляется» другим. У гетерозигот признак проявляется частично, тогда как у доминантных гомозигот полностью. Пример: ген заболевания цистиинурии (камни в почках) в гетерозиготном состоянии возможны отдельные признаки заболевания, а в гомозиготном – тяжелая форма болезни.
Сверхдоминирование – у доминантного аллеля в гетерозиготном состоянии отмечается более сильное проявление, чем в гомозиготном состоянии. Пример: ген крупной масти у рыб проявляется в гетерозиготном состоянии больше (рыбы крупнее), чем в гомозиготном (рыбы средней величины). Такое проявление напоминает эффект гетерозиса у растений (гибридная сила). Так, в случае браков между представителями разных рас показатели здоровья их потомков превосходят таковое самих родителей: дети отличаются более высокой продолжительностью жизни, жизнеспособностью и др.
Кодоминирование – такое взаимодействие, при котором оба гена из аллельной пары проявляют себя в равной степени доминантными, в результате у гетерозигот появляются новые свойства, не похожие на свойства родителей.
Пример: наследование групп крови. Ген группы крови А и ген группы крови В совместно в генотипе дают IV группу крови АВ.
| Группа крови | Антиген в эритроците | Антитело в сыворотке | Генотип |
| О (I) А (II) В (III) АВ (IV) | 0 А В АВ | αβ β α 0 | І0І0 ІАІ0 и ІАІА ІВІ0 и ІВІВ ІАІВ |
Из данных таблицы видно, что люди с первой группой крови не имеют антигена в эритроцитах, следовательно, могут быть универсальными донорами крови (теоретически). 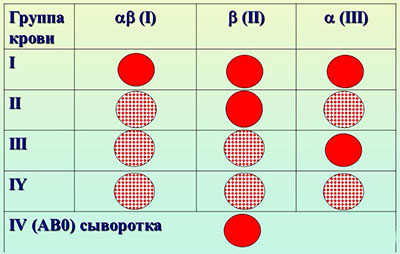 А люди с четвертой группой крови, не имеющие антител в сыворотке крови – универсальными реципиентами (теоретически).
А люди с четвертой группой крови, не имеющие антител в сыворотке крови – универсальными реципиентами (теоретически).
Кодоминирование наблюдается и при наследовании групп крови по системе М N, открытой в 1927 году. Эта система определяется двумя аллелями: IМ, IN. Оба аллеля кодоминантны, поэтому существуют люди со следующими генотипами: IМIМ, ININ, IМ IN. В сыворотке крови людей с тем или иным фенотипом по этой системе нет антител к соответствующим антигенам, поэтому при переливании крови эта система может не учитываться.
Множественное действие генов (плейотропия) – такое явление, когда один ген определяет развитие нескольких признаков (от греч. Плейрос – наибольший, тропе – направление). Пример: ген арахнодактилии (паучьи пальцы) или болезнь Марфана вызывает нарушение развития соединительной ткани и оказывает влияние на развитие нескольких признаков: нарушение в строении хрусталика глаза, аномалии в строении сердечно-сосудистой системы, суставов и т.д.
Такое понятие как множественный аллелизм в противоположность множественному действию генов, обозначает взаимодействие нескольких пар аллельных генов, отвечающих за развитие одного признака. Они получили название серии множественных аллелей. Возникают они в результате многократного мутирования одного и того же локуса в хромосоме. Так, кроме доминантного и рецессивного аллелей гена появляются промежуточные аллели, которые по отношению к доминантному ведут себя как рецессивные, а по отношению к рецессивному ведут себя как доминантные. Наличие множественных аллелей показывает, что возможно существование не двух состояний гена – доминантного и рецессивного, а целого ряда изменений определенного участка хромосомы, возникающих путем мутаций. Данный вид взаимодействия наблюдается при наследовании групп крови, структуры гемоглобина, цвета глаз дрозофилы, окраски шерсти кроликов и т.д. Хотя описываемый феномен открыт относительно недавно, уже известно более 100 моногенных болезней, обусловленных мутациями одного гена по типу аллельной серии. Например, точковые мутации гена рецептора тирозинкиназы – RET, вызывают четыре наследственные болезни: семейную медуллярную карциному щитовидной железы, болезнь Гиршпрунга, множественную эндокринную неоплазию двух типов – 2А и 2В.
Компаунд – гетерозигота – гетерозиготность по двум аномальным аллельным генам, приводящая к тяжелой форме заболевания. Примером такого наследования может быть наследование структуры гемоглобина (Hb A – гемоглобин взрослого человека, Hb Р – анемия Пенджа, Hb D – анемия Джамбул, Hb O – анемия Ораб и т.д.). Некоторые лица с этим метаболическим нарушением представляют собой компаунд-гетерозиготы, а не истинные гомозиготы. Компаунд-гетерозиготами называют лиц, получивших разные, а не идентичные мутантные аллели определенного локуса от каждого родителя. Первыми идентифицированными компаунд-гетерозиготами были лица с болезнью гемоглобина SC. Они наследуют ген гемоглобина S от одного из родителей и ген гемоглобина С от другого. Эти больные обладают двойной дозой мутации по синтезу b-глобиновой цепи, и поэтому у них не образуются нормальные b-цепи. Клинически и химически они отличаются от истинных SS- или СС-гомозигот. Компаунд-гетерозиготы выявлены также среди больных с цистинурией, иминоглицинурией, недостаточностью галактозо-1-фосфатуридилтрансферазы, L-идуронидазы, метилмалонил-КоА-мутазы и цистатионинсинтетазы. У некоторых, но не у всех компаунд-гетерозигот тяжесть состояния не уступает таковой у истинных гомозигот, что зависит от природы наследуемых мутантных аллелей.
Межаллельная комплементация – восстановление у гетерозиготы по двум мутантным формам пептида нормально функционирующей четвертичной структуры белка. Установлено, что все гены, аллели которых взаимодействуют таким образом, контролируют структуру ферментов, построенных из одинаковых белковых субъединиц. Если же мутантные аллели отличаются тем, что кодируют полипептиды различных доменов, то при их объединении в гетерозиготе в молекуле фермента соберутся все необходимые функциональные центры (так как мутантные аллели ответственны за дефектность лишь некоторых из них), то ферментативная активность, а значит, и нормальный фенотип будут восстановлены. Примером данного типа взаимодействия аллелей является проявление мутации ade у дрожжей, заключающейся в отсутствии активности фермента фосфорибозиламиноимидазолкарбоксилазы. При этом мутанты имеют красный цвет, обусловленный тем, что из-за отсутствия этого фермента они вынуждены поглощать экзогенный аденин и субстрат вместе с ним, а последний, накапливаясь в клетках, придаёт им красную окраску. У нормальных форм сохраняется белый цвет.
Аллельное исключение – у гетерозиготного организма в различных клетках, синтезирующих разнообразные белки, проявляется действие разных аллелей. Возможность проявления разных аллелей увеличивает разнообразий синтезируемых белков. Такая картина наблюдается в В-лимфоцитах, синтезирующих антитела к определённым антигенам. Поскольку эти иммуноглобулины моноспецифичны, каждый лимфоцит должен выбрать между экспрессией отцовского или материнского аллеля. Аллельное исключение происходит и при инактивации одной из Х-хромосом у особей гомогаметного пола, приводящего в соответствие дозы Х-генов у всех представителей пола (к примеру, у женщин в каждой клетке экспрессируется лишь одна Х-хромосома, другая же переходит в неактивный гетерохроматин (тельце Барра). Таким образом, женский организм у человека является мозаичным, т.к. в разных клетках могут экспрессироваться разные хромосомы). При этом проявляются только аллели, находящиеся в активной Х-хромосоме, те же аллели, которые находятся в инактивированной, как бы исключаются и не проявляются в фенотипе. При этом в каждой клетке экспрессируется или один, или другой аллель, в зависимости от того, какая Х-хромосома была инактивирована.
Эффект положения генов – это влияние расположения генов в хромосоме на проявление их активности (А. Стертевант, 1925г.), изменение экспрессии гена при изменении занимаемого им положения в хромосоме (в ряду нуклеотидных последовательностей). По современным представлениям, эффект положения не связан с нарушением структуры гена: он и его промоторная область сохраняются как единица транскрипции. Уровень экспрессии зависит от местоположения гена в геноме: либо в районах конденсированного гетерохроматина, либо в районах деконденсированного хроматина (эухроматина). При перемещении гена в область гетерохроматина (неактивную часть генома) его активность снижается. Так, у гетерозигот, вследствие эффекта положения, затронувшего доминантный ген, будет наблюдаться фенотип рецессивного. Либо, наоборот, «молчащий» ген перемещается в активно транскрибируемую область генома. Свойство обратимости при эффекте положения гена используют для доказательства того, что наблюдаемое изменение проявления данного гена – эффект положения гена, а не его мутация. В результате исчезают пуфы в эухроматиновых участках, нарушается синтез ДНК и РНК: гетерохроматин при переходе в эухроматин активируется и становится цитологически не отличим от эухроматина. Нарушение активности при эффекте положения гена может наблюдаться одновременно у нескольких эухроматиновых генов, расположенных за геном, непосредственно прилегающим к гетерохроматину, причем влияние гетерохроматина всегда направлено от места перестройки к ближайшему эухроматиновому гену и по мере увеличения расстояния между эухроматиновыми и гетерохроматиновыми генами это влияние ослабляется (эффект поляризованного распространения). Примером проявления эффекта положения является наследование резус-фактора крови человека. Резус-положительная группа крови определяется присутствием на поверхности эритроцитов особой группы антигенов. Они имеют сложную структуру, их синтез обусловлен тремя доминантными генами. Однако на самом деле проявление признака носит более сложный характер: один из трех генов является определяющим, но наличие двух других в доминантном или рецессивном варианте влияет на его экспрессию. В частности, лица с генным комплексом CDE/ Cde имеют те же гены, что и лица с генным комплексом cDe/CDE. Однако у первых много антигена Е и мало антигена С, а у вторых – наоборот, что объясняется неодинаковой локализацией генов С и Е – на одной хромосоме или на разных хромосомах соответственно. Эффект положения может проявляться при некоторых видах рака.
Модифицирующее действие генов – действие одного гена может усиливать (интенсификатор) или ослаблять (супрессор) действие другого гена.
Дата: 2018-11-18, просмотров: 1145.