Глава
Классификация нейронов.
По числу отростков.
1. Униполярные нейроны имеют 1 отросток. По мнению большинства исследователей, такие нейроны не встречаются в нервной системе млекопитающих и человека.
2. Биполярные нейроны – имеют 2 отростка: аксон и дендрит. Разновидностью биполярных нейронов являются псевдоуниполярные нейроны спинномозговых ганглиев, где оба отростка (аксон и дендрит) отходят от единого выроста клеточного тела.
3. Мультиполярные нейроны – имеют один аксон и несколько дендритов. Их можно выделить в любом отделе нервной системы.
По форме.
Веретеновидные, грушевидные, пирамидные, полигональные. Такой подход лежит в основе изучения цитоархитектоники мозга.
По выполняемой функции.
1. Чувствительный (афферентный) – помогающий воспринимать внешние раздражители (стимулы).
2. Ассоциативный (вставочный интернейрон).
3. Двигательные (эфферентные) – вызывающие сокращения и движения. Именно эти нейроны получили наименование «мотонейроны», т.е. двигательные нейроны, сконцентрированные в двигательных ядрах передних рогов спинного мозга и стволовой части головного мозга.
Биохимическая классификация.
1. Холинергические (медиатор – АХ – ацетилхолин).
2. Катехоламинергические (А, НА, дофамин).
3. Аминокислотные (глицин, таурин).
По принципу положения их в сети нейронов.
Первичные, вторичные, третичные и т.д.
Исходя из такой классификации, выделяют и типы нервных сетей:
- иерархические (восходящие и нисходящие);
- локальные – передающие возбуждение на каком-либо одном уровне;
- дивергентные с одним входом (находящиеся в основном только в среднем мозге и в стволе мозга) – осуществляющие связь сразу со всеми уровнями иерархической сети. Нейроны таких сетей называют «неспецифическими».
Именно к неспецифическим сетям относятся ретикулярные нейроны – многоугольные нейроны, образующие промежуточную зону серого вещества спинного мозга (включая боковые рога), ядра ретикулярной формации продолговатого и среднего мозга (включая вегетативные ядра соответствующих черепно-мозговых нервов), образования субталамической и гипоталамической областей промежуточного мозга.
По длине аксона.
В рамках этой классификации короткими считаются те аксоны, ветви которых остаются в непосредственной близости от тела клетки.
1. клетки 1-го типа Гольджи (эфферентные) – нейроны с длинным аксоном, продолжающимся в белом веществе мозга;
2. клетки 2-го типа Гольджи (вставочные) – нейроны с коротким аксоном, разветвления которого выходят за пределы серого вещества мозга.
По скорости проведения импульсов по аксонам.
Гассер разделил волокна на три основные группы: А, В и С.
Волокна групп А и В миелинизированы. Различия между группами А и В несущественны.
Диаметр волокон типа А варьирует от 4 до 20 мкм, а скорость, с которой импульсы проходят по ним, определяемая в м/сек, приблизительно равна величине их диаметра в микронах, умноженной на 6.
Диаметр волокон типа С значительно меньше - от 0,3 до 1,3 мкм, а скорость проведения импульсов в них несколько меньше величины диаметра, умноженной на 2.
Гассер подразделял А-волокна по скорости проведения. Волокна с наибольшей скоростью проведения были названы «А-альфа», средней – «А-бета» и наименьшей – «А-гамма». Поскольку скорость проведения прямо пропорциональна диаметру, эти обозначения иногда используются для классификации типов миелинизированных волокон.
По форме нервных клеток.
1. Пирамидные.
2. Звездчатые.
3. Веретеновидные.
4. Переходные формы.
Пирамидные нейроны в коре имеют различную величину. Они встречаются во всех слоях коры.
Наиболее крупные пирамидные нейроны находятся в слое IV зрительной области коры и в слоях III и V других корковых зон.
Особо крупные пирамидные нейроны – нейроны Беца (по имени В.А. Беца, впервые их описавшего) обнаружены в области коркового конца двигательного анализатора.
В отдельных областях коры пирамидные нейроны особенно богато представлены в слое III; в местах деления этого слоя на три подслоя наиболее крупные пирамидные нейроны находят в третьем подслое. Они, как правило, имеют верхушечный (аликальный) дендрит со значительным разветвлением, направленный к поверхности коры. В большинстве случаев верхушечные дендриты доходят до слоя I коры, где и разветвляются в горизонтальном направлении. От основания пирамидного нейрона в горизонтальном направлении отходят базальные и боковые дендриты, также постепенно дающие разветвления различной длины. Единственный длинный аксон, отходящий от пирамидного нейрона, направляется вниз в белое вещество и дает разветвленные по разным направлениям коллатерали. Иногда его ответвления образуют дугу и направляются к поверхности коры, давая по пути отростки, образующие межнейронные связи.
Звездчатые клетки коры больших полушарий, особенно у человека, многообразны. Нейроны этого типа составляют значительную часть всех клеточных элементов коры больших полушарий мозга человека. Весьма разнообразны и богаты разветвлениями их дендритные и аксонные окончания, особенно в верхних слоях коры. Аксоны звездчатых нейронов, в отличие от аксонов пирамидных и веретеновидных клеток, как правило, не выходят за пределы коры больших полушарий, а зачастую и за пределы одного слоя. В коре больших полушарий наблюдаются значительные различия в сложности форм и многообразии дендритных и аксонных разветвлений звездчатых нейронов: особенно многообразны межнейронные связи.
Веретеновидные нейроны характерны в основном для VI-VII слоев коры. Однако веретеновидные нейроны нередко встречаются и в V слое. Наиболее характерной особенностью веретеновидных нейронов является наличие у них двух дендритов, направленных в противоположные стороны. Часто наряду с этими основными дендритами и их разветвлениями от тела веретеновидных клеток отходит еще боковой дендрит, идущий в горизонтальном направлении. Дендриты веретеновидных клеток обычно образуют немного разветвлений. Разветвления аксонов веретеновидных клеток также весьма незначительны по сравнению с разветвлениями звездчатых и пирамидных нейронов. Верхушечный дендрит веретеновидной клетки, поднимаясь вверх, может дойти до I слоя, однако в большинстве своем эти дендриты оканчиваются в слоях V, IV и III.
Греческое слово «глия» означает «клей». Глиальные клетки впервые описал в 1846г. Р. Вирхов, который считал, что они «склеивают», скрепляют нервные клетки, «придавая целому его особую форму».
Глиальные клетки выполняют в нервной системе множество еще не совсем ясных функций. В отличие от нейронов глиальные клетки сохраняют способность к делению в течение всей жизни.
Хотя они имеют мембранный потенциал, но способны генерировать потенциал действия — возбуждаться. Нейроглия составляет почти половину объема мозга, а число клеток глии значительно превышает число нейронов (по меньшей мере в 10 раз).
Различают три типа глиальных клеток: астроглия, олигодендроглия и микроглия.
Астроглия. Происходит из спонгиобластов, развивающихся в клетки, имеющие множество отростков. Длинные извитые отростки астроцитов переплетаются с отростками нейронов. Значительное число отростков астроцитов представляют собой «ножки», плотно прилегающие к капиллярам и покрывающие собой почти всю поверхность сосуда. Астроциты, расположенные в местах концентрации тел нейронов (серое вещество), образуют больше отростков, чем астроциты в белом веществе. Таким образом, астроциты — это клетки, располагающиеся между капиллярами и телами нейронов и осуществляющие транспорт веществ из крови в нейроны и обратно. Кроме того, астроглия связывает с кровеносным руслом спинномозговую жидкость.
Олигодендроглия . Олигодендроциты имеют то же происхождение, что и астроциты. По размерам они меньше, чем астроциты и имеют меньше отростков. Основная масса олигодендроцитов располагается в белом веществе мозга и ответственна за образование миелина. Эти олигодендроциты обладают длинными отростками. Олигодендроциты, расположенные в периферической нервной системе, называются Шванновскими клетками (см. рис. 2). Те олигодендроциты, которые находятся в сером веществе, располагаются, как правило, вокруг тел нейронов, плотно прилегая к ним. Поэтому их называют клетками-сателлитами. Они характеризуются наличием коротких отростков (см. рис. 3).
Микроглия. Клетки микроглии происходят из мезодермы. Как видно из названия, они отличаются небольшими размерами. Эти клетки могут активно передвигаться и выполнять фагоцитарные функции. Благодаря способности к активной миграции микроглия распределена по всей центральной нервной системе.
Часть аксонов центральной нервной системы покрывается специальным электроизолирующим веществом — миелином. Миелинизацию аксонов осуществляют клетки глии. В центральной нервной системе эту роль выполняют олигодендроциты, в периферической — Шванновские клетки (рис. 2, 3), являющиеся разновидностью олигодендроцитов. Олигодендроцит оборачивается вокруг аксона, образуя многослойную оболочку. Миелинизации не подвергается область аксонного холмика (рис. 2, 3) и терминали аксона. Цитоплазма глиальной клетки выдавливается из межмембранного пространства в процессе «обертывания». Таким образом, миелиновая оболочка аксона состоит из плотно упакованных, перемежающихся липидных и белковых мембранных слоев (рис. 3). Аксон не сплошь покрыт миелином. В миелиновой оболочке существуют регулярные перерывы — перехваты Ранвье. Ширина такого перехвата от 0,5 до 2, 5 мкм. Функция перехватов Ранвье — быстрое скачкообразное (сальтаторное) распространение потенциалов действия, осуществляющееся без затухания. В центральной нервной системе аксоны различных нейронов, направляющиеся к одной структуре, образуют упорядоченные пучки — проводящие пути. В подобном проводящем пучке аксоны направляются «параллельным курсом» и часто одна глиальная клетка образует оболочку нескольких аксонов. Поскольку миелин является веществом белого цвета, то проводящие пути нервной системы, состоящие из плотно лежащих миелинизированных аксонов, образуют белое вещество мозга. В сером же веществе мозга локализуются тела клеток, дендриты и немиелинизированные части аксонов.
Рис. 2. Типы нейронов:
а — сенсорные нейроны: 1 — биполярный; 2 — псевдобиполярный; 3 — псевдоуниполярный; б — двигательные нейроны: 4 — пирамидная клетка; 5 — мотонейроны спинного мозга; 6 — нейрон двойного ядра; 7 — нейрон ядра подъязычного нерва; в — симпатические нейроны: 8 — нейрон звездчатого ганглия; 9 — нейрон верхнего шейного ганглия; 10 — нейрон бокового рога спинного мозга; г — парасимпатические нейроны: 11 — нейрон узла мышечного сплетения кишечной стенки; 12 — нейрон дорсального ядра блуждающего нерва; 13 — нейрон ресничного
Сведения о форме нейронов, их местоположении и направлении отростков очень важны, поскольку позволяют понять качество и количество связей, приходящих к ним (структура дендритного дерева), и пункты, в которые они посылают свои отростки.
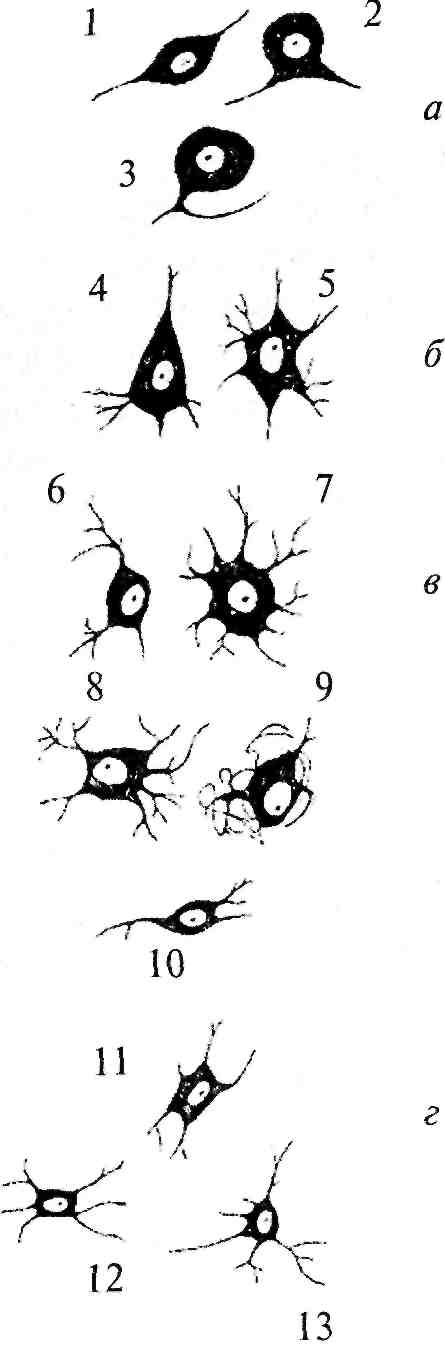
Рис. 3. Схема миелинизации аксонов:
1 — связь между телом клетки глии и миелиновой оболочкой; 2 — олигодендроцит; 3 — гребешок; 4 — плазматическая мембрана; 5 — цитоплазма олигодендроцита; 6 — аксон нейрона; 7 — перехват Ранвье; 8 — мезаксон; 9 — петля плазматической мембраны
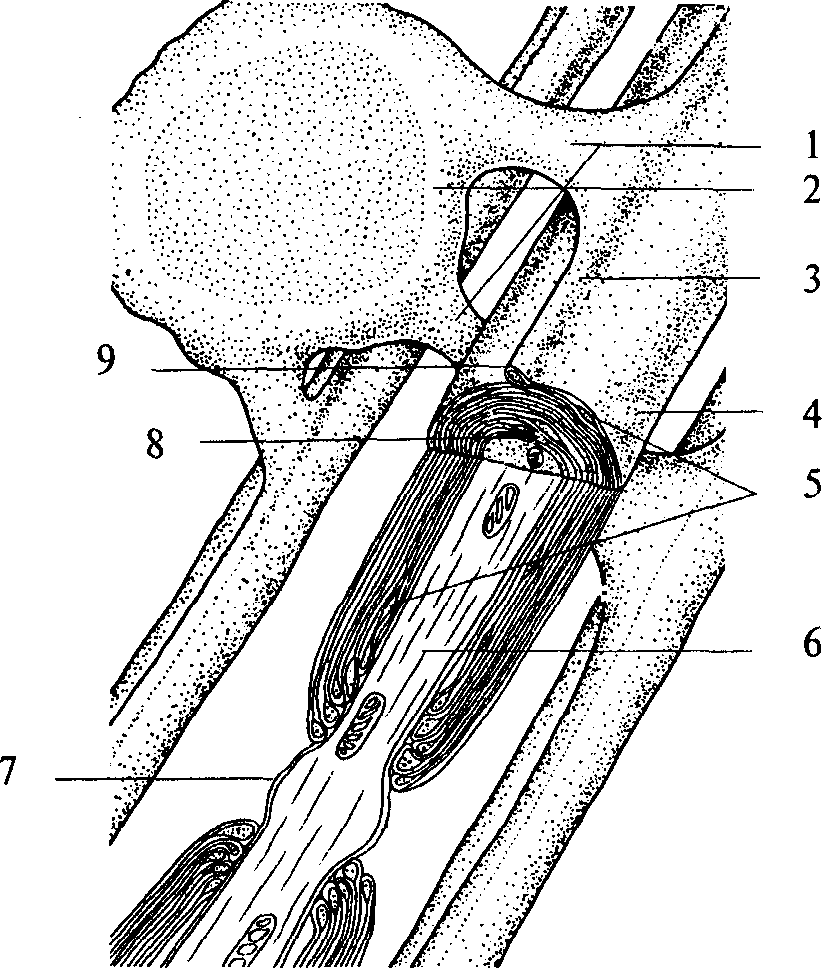
Рис. 4. Строение синапса:
1 — аксон пресинаптического нейрона; 2 — микротрубочки; 3 — синаптический пузырек (везикула); 4 — синаптическая щель; 5 — дендрит постсинаптического нейрона; 6 — рецептор для медиатора; 7 — постсинаптическая мембрана; 8 — пресинаптическая мембрана; 9 — митохондрия
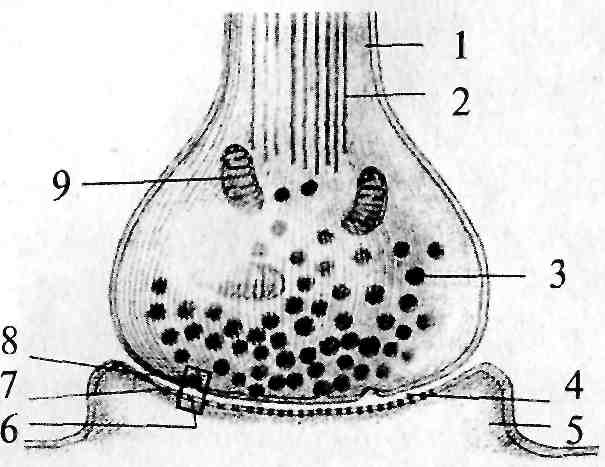
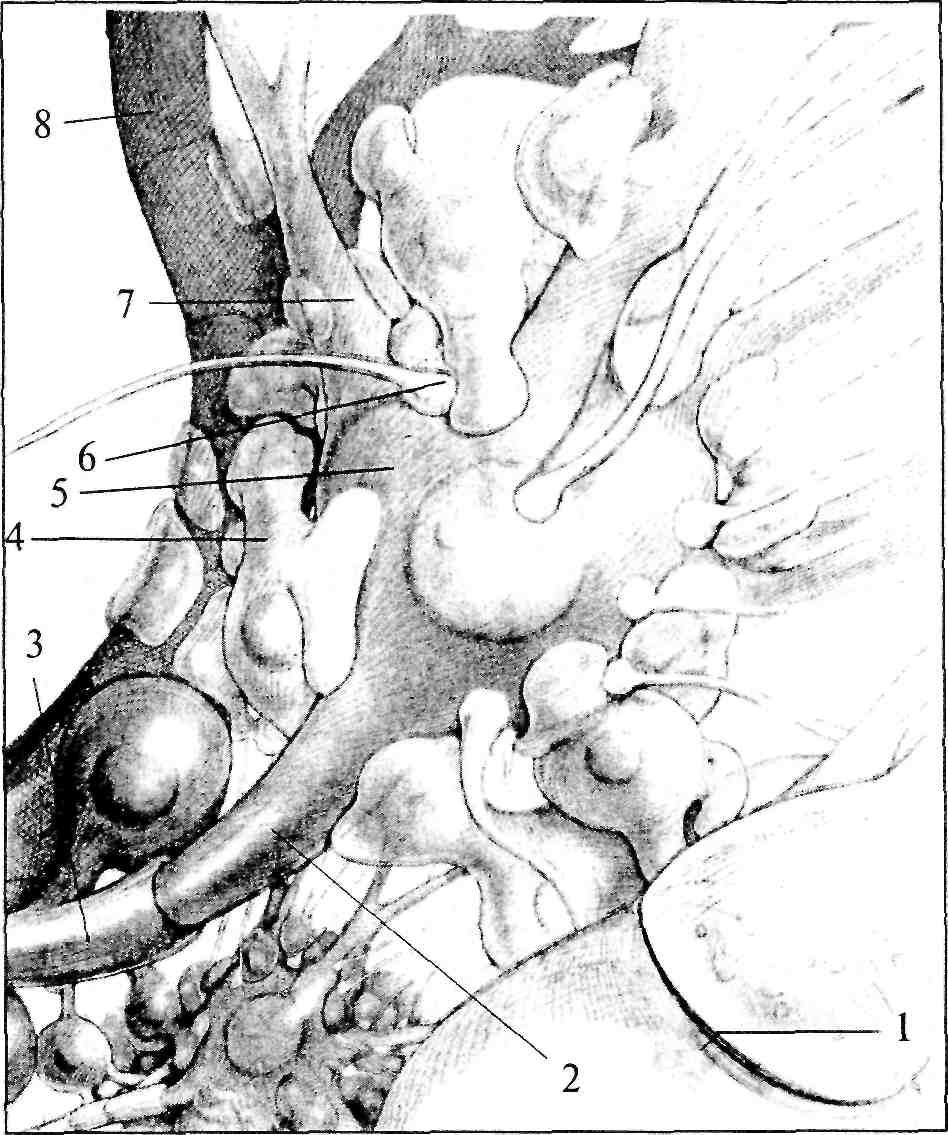
Рис. 4. Схема взаиморасположения элементов нервной ткани:
1 — синапс; 2 — аксонный холмик; 3 — миелиновая оболочка аксона; 4 — астроцит; 5 — тело нейрона; 6 — синаптическая бляшка; 7 — дендрит; 8 — капилляр
Оболочки спинного мозга
Снаружи спинной мозг окружен тремя оболочками.
Наружная - твердая оболочка спинного мозга фиксирована связочным аппаратом в позвоночном канале и отделена от надкостницы этого канала эпидуральным пространством, заполненным жировой клетчаткой и венозным сплетением.
Средняя - тонкая оболочка спинного мозга, прозрачная, называется паутинной (арахноидальной) оболочкой спинного мозга.
Внутренняя – мягкая оболочка спинного мозга, тесно прилегает к спинному мозгу и содержит кровеносные сосуды, питающие спинной мозг.
Твердая оболочка спинного мозга представляет собой продолговатой формы мешок с прочными и толстыми стенками, расположенный в позвоночном канале и содержащий спинной мозг с корешками и оболочками.
Внутренняя поверхность твердой оболочки спинного мозга отделена от паутинной оболочки узким щелевидным субдуральным пространством, которое пронизано большим количеством тонких соединительнотканных перекладин. Вверху субдуральное пространство спинного мозга свободно сообщается с аналогичным пространством в полости черепа. Внизу это пространство заканчивается слепо на уровне II крестцового позвонка.
Паутинная оболочка спинного мозга плотно прилегает к спинному мозгу. В ней различают два слоя - внутренний и наружный, между которыми располагаются кровеносные сосуды. Внутренний слой оболочки очень прочно сращен с тканью спинного мозга и в виде отростков внедряются в него вместе с кровеносными сосудами.
Кровоснабжение спинного мозга осуществляется позвоночной артерией - ветвью подключичной артерии, а также от задних межреберных, поясничных и латеральных крестцовых артерий спинного мозга.
Возрастные особенности
У новорожденного спинной мозг составляет в длину 14 см, к двум годам – 20 см, к 10 годам – 29 см. Масса спинного мозга у новорожденного составляет 5,5 гр., к двум годам – 13 гр., к 7 годам – 19 гр. У новорожденного хорошо выражены два утолщения, а центральный канал шире, чем у взрослого. В первые два года происходит изменение просвета центрального канала. Объем белого вещества возрастает быстрее, чем объем серого вещества.
Тема: Мозговые оболочки
Вопросы:
ü Оболочки спинного мозга. Строение и топография твердой, паутинной и мягкой оболочек спинного мозга.
ü Головной мозг. Начальные этапы развития головного мозга.
ü Стадия трех и пяти мозговых пузырей, образование изгибов головного мозга, происхождение желудочков головного мозга.
ü Оболочки головного мозга: строение, топография и функции твердой, паутинной и сосудистой оболочек.
Кора головного мозга.
Строение.
Кора представляет собой филогенетически наиболее молодой и вместе с тем сложный отдел мозга, предназначенный для обработки сенсорной информации, формирования поведенческих реакций организма.
Кора больших полушарий делится на древнюю (обонятельная луковица, обонятельный тракт, обонятельный бугорок), старую (часть лимбической системы) и новую кору. Новая кора занимает 95-96% общей площади и 4-5% приходится на долю древней и старой коры. Толщина коры колеблется от 1,3 до 4,5 мм. Площадь коры увеличивается за счет борозд и извилин. У взрослого человека она составляет 2200 см²
Кора состоит из серого и белого вещества, а также нейроглии. Количество нейронов 16-18 млрд. Глиальные клетки выполняют трофическую функцию.
По функциональному признаку нейроны коры делятся на 3 вида: афферентные (сенсорные) – к ним подходят нервные волокна афферентных путей, ассоциативные (вставочные) – в пределах головного и спинного мозга, эфферентные (двигательные) – образует нисходящие (эфферентные) проводящие пути, идущие от коры к разным ядрам головного и спинного мозга. К сенсорным клеткам относятся звездчатые клетки, входящие в 3 и 4 слоя сенсорных областей коры. К эфферентным нейронам относятся нейроны 5 слоя моторной зоны, которые представлены гигантскими пирамидными клетками Беца. К ассоциативным клеткам относятся веретенообразные и пирамидные клетки 3 слоя.
В связи с тем, что тела и отростки описанных выше нейронов имеют упорядоченное расположение, кора построена по экранному принципу, т.е. сигнал фокусируется не точка в точку, а на множество нейронов, что обеспечивает полный анализ раздражителя, а также возможность передачи сигнала в другие зоны коры, которые заинтересованы в нем.
Кора состоит из 7 слоев.
1. Молекулярный слой – мелкие нейроны и волокна. Сюда приходят афферентные таламокортикальные волокна от неспецифических ядер таламуса, регулирующие уровень возбудимости корковых нейронов.
2. Наружный зернистый слой образован мелкими нейронами в форме зерен и мелкими пирамидными клетками.
3. Наружный пирамидный слой состоит из пирамидных клеток разной величины. Функционально II и III слои коры объединяют нейроны, отростки которых обеспечивают кортико-кортикальные ассоциативные связи.
4. Внутренний зернистый слой образован звездчатыми клетками. Здесь оканчиваются афферентные таламокортикальные волокна, идущие от проекционных ядер таламуса.
5. Внутренний пирамидный слой включает крупные пирамидальные клетки – клетки Беца, аксоны которых идут в головной и спинной мозг.
6. Полиморфный слой (мультиформный) – многоформенные нейроны, имеющие треугольную и веретенообразную форму.
7. Веретенообразные нейроны связывают все слои коры, их волокна поднимаются до 1 слоя. Имеются только в некоторых областях коры.
Функциональной единицей коры является вертикальная колонка, состоящая из 7 клеток, они вместе реагируют на один и тот же раздражитель.
В коре выделяют сенсорные, ассоциативные и двигательные зоны, исходя из расположения нейронов:
Сенсорные зоны – это входные участки коры, которые через восходящие нервные пути получают сенсорную информацию от большинства рецепторов тела.
Ассоциативные зоны – 1) связывают вновь поступающую сенсорную информацию с полученной ранее и хранящейся в блоках памяти, благодаря чему новые стимулы «узнаются», 2) информация от одних рецепторов сопоставляются с сенсорной информацией от других рецепторов, 3) участвуют в процессах запоминания, научения и мышления.
Двигательные зоны – выходные области коры. В них возникают двигательные импульсы, идущие к произвольным мышцам по нисходящим путям, которые находятся в белом веществе больших полушарий.
Цитоархитектоника – это расположение нейронов в коре.
Миелоархитектоника – это распределение волокон в коре головного мозга.
Начало разнокачественного строения коры больших полушарий было положено в 1674 г. киевским анатомом А.А. Бецом. Позже К. Бродман в 1903-09 гг. выделил 52 цитоархитектонических полей. О. Фогт и Ц. Фогт выделили в коре 150 миелоархитектонических полей.
Локализация функций в коре больших полушарий.
И.П. Павлов рассматривал кору больших полушарий как сплошную воспринимательную поверхность, как совокупность корковых концов анализаторов. Анализатор – сложная система, которая состоит из рецептора - воспринимающего аппарата, проводников нервных импульсов и мозгового конца, где происходит высший анализ раздражений. И.П. Павлов показал, что в коре различают ядра и рассеянные элементы. Ядро – это место концентрации нейронов, где проецируются все структуры периферического рецептора и происходит важный анализ и синтез, и интеграция функций.
Рассеянные элементы могут располагаться по периферии ядра и на различном расстоянии от него. В них происходит более простой анализ и синтез.
Корковые концы анализатора осуществляют анализ и синтез сигналов.
Рассмотрим некоторую локализацию ядер моторных анализаторов:
1. Ядро коркового анализатора общей чувствительности (температурной, болевой, осязательной и проприоцептивной) - постцентральная извилины (поля 1, 2, 3) и верхняя теменная долька (поля 5 и 7). Проводящие чувствительные пути, следующие в кору большого мозга, перекрещиваются либо на уровне различных сегментов спинного мозга (пути болевой, температурной чувствительности, осязания и давления), либо на уровне продолговатого мозга (пути проприоцептивной чувствительности коркового направления). Вследствие этого постцентральные извилины каждого из полушарий связаны с противоположной половиной тела. В постцентральной извилине рецепторные поля различных участков тела человека спроецированы таким образом, что наиболее высоко расположены корковые концы анализатора чувствительности нижних отделов туловища и нижних конечностей, а наиболее низко (ближе к латеральной борозде) проецируются рецепторные поля верхних участков тела, головы и верхних конечностей.
2. Ядро двигательного - двигательная область коры (процентральная извилина (поля 4 и 6) и парацентральная долька на медиальной поверхности полушария). В 5 слое коры предцентральной извилины залегают пирамидные нейроны (клетки Беца), которые И.П. Павлов относил к вставочным, и отмечал, что эти клетки своими отростками связаны с подкорковыми ядрами, двигательными клетками ядер черепных и спинномозговых нервов. Причем в верхних участках предцентральной извилины и в парацентральной дольке расположены клетки, импульсы от которых направляются к мышцам самых нижних отделов туловища и нижних конечностей. В нижней части предцентральной извилины находятся также двигательные центры, регулирующие деятельность мышц лица.
Таким образом, все участки тела человека спроецированы в предцентральной извилине «вверх ногами». В связи с тем, что пирамидные пути, берущие начало от гигантопирамидных клеток, перекрещиваются либо на уровне мозгового ствола (корково-ядерные волокна) на границе со спинным мозгом, либо в сегментах спинного мозга (корково-спинномозговой путь), двигательные области каждого из полушарий связаны со скелетными мышцами противоположной стороны тела. Если мышцы конечностей изолированно связаны с одним из полушарий, мышцы туловища, гортани и глотки имеют связь с двигательными областями обоих полушарий.
3. Ядро зрительного анализатора располагается в затылочной доле полушария большого мозга (поля 17, 18, 19). Ядро зрительного анализатора правого полушария связано проводящими путями с латеральной половиной сетчатки правого глаза и медиальной половиной сетчатки левого глаза. В коре затылочной доли левого полушария проецируюся соответственно рецепторы латеральной половины сетчатки левого глаза и медиальной половины сетчатки правого глаза.
4. В глубине латеральной борозды на обращенной к островку поверхности средней части верхней височной извилины находится ядро слухового анализатора (поля 41, 42, 52). К нервным клеткам, составляющим ядро слухового анализатора каждого из полушарий, проходят проводящие пути от рецепторов как левой, так и правой стороны. В связи с этим одностороннее поражение этого ядра не вызывает полной утраты способности воспринимать звуки. Двустороннее поражение сопровождается корковой глухотой, как и в случае полной корковой слепоты.
5. Ядро двигательного анализатора артикуляции речи (речедвигательный анализатор) располагается в задних отделах нижней лобной извилины (поле 44). Повреждение участка коры этой области (поле 44) приводит к двигательной афазии, т.е. утрате способности к сокращению мышц, участвующих в речеобразовании. Более того, при повреждении поля 44 не утрачивается способность к произношению звуков или пению.
В центральных отделах нижней лобной извилины (поле 45) находится ядро речевого анализатора, связанного с пением. Поражение поля 45 сопровождается вокальной амузией – неспособностью к составлению и воспроизведению музыкальных фраз, и аграмматизмом, когда утрачивается способность к составлению осмысленных предложений из отдельных слов. Речь таких людей состоит из не связанного по смысловому значению набора слов.
6. Ядро слухового анализатора устной речи тесно взаимосвязано с корковым центром слухового анализатора и располагается, как и последний, в области верхней височной извилины (поле 42).
Поражение ядра не нарушает слухового восприятия звуков, однако утрачивается способность понимать слова, речь. Функция этого ядра состоит в том, что человек не слышит и не понимает речь другого человека, но контролирует свою собственную.
В средней трети верхней височной извилины (поле 22) находится ядро коркового анализатора, поражение которого сопровождается наступлением музыкальной глухоты, когда музыкальные фразы воспринимаются как бессмысленный набор различных шумов. Этот корковый конец слухового анализатора относится к центрам второй сигнальной системы, воспринимающим словесное обозначение предметов, действий, явлений, т.е. воспринимающих сигналы сигналов.
7. В непосредственной связи с ядром зрительного анализатора находится ядро зрительного анализатора письменной речи (поле 39), расположенное в угловой извилине нижней теменной дольки. Поражение этого ядра приводит к утрате способности воспринимать написанный текст, читать.
Различают в коре 3 группы полей: первичные, вторичные и третичные.
Первичное поле связано с органами чувств и органами движения, оно раньше формируется в онтогенезе и имеет наиболее крупные клетки. Это так называемые ядерные зоны анализаторов. Они осуществляют анализ раздражений, поступающих в кору от соответствующих рецепторов. Если разрушить ядерную зону, наступит корковая слепота, глухота, двигательный паралич.
Вторичные поля (периферические зоны анализаторов) связаны с отдельными органами только через первичные поля. Они служат для обобщения и дальнейшей обработки поступающей информации. Если разрушить это поле, человек видит, слышит, но не понимает смысла.
Третичные поля (зоны перекрытия анализаторов) занимают почти половину территории коры и имеют обширные связи с другими отделами коры и неспецифическими системами мозга. Здесь в основном располагаются мелкие и разнообразные (звездчатые) клетки и происходит высший анализ и синтез информации, в результате чего вырабатываются цели и задачи поведения. Согласно им происходит программирование двигательной деятельности. При врожденном недоразвитии третичных полей человек не в состоянии овладеть речью и даже простыми двигательными навыками.
Первичные и вторичные поля есть у человека и животных, а третичное поле только у человека. Третичные поля созревают у человека позже других корковых полей. Для развития полей необходимо, чтобы больше информации поступало от зрительных, слуховых, мышечных рецепторов.
Онто- и филогенез коры.
К 30-тидням внутриутробного развития формируется кора. К 7‑12‑му месяцам постнатального развития происходит созревание мозговых систем.
У новорожденного развиты филогенетически старые отделы мозга: мозжечок, мост, а также промежуточный мозг. У новорожденных основные борозды и извилины (центральная, латеральная) выражены хорошо, а ветви борозд и извилин слабо. Миелинизация афферентных волокон начинается в 2 месяца и заканчивается к 4-5 годам, а эфферентные волокна несколько позже – от 4-5 месяцев до 7-8 лет. Соотношения борозд, извилин и швов, характерные для взрослого человека, устанавливаются у детей в 6-8 лет.
Промежуточный мозг.
Вопросы:
ü Таламический мозг и его части:
ü таламус, гипоталамус, метаталамус, эпиталамус.
ü Ядра зрительного бугра, их классификация, топография и структура, связи с другими отделами мозга.
ü Гипоталамус, классификация и топография основных ядер гипоталамуса. Гипофиз и его функции.
ü Третий мозговой желудочек.
Гипоталамус и поведение.
Гипоталамус выполняет следующие функции:
- участвует в регуляции пищеварения, поведения, которое тесно связано с уменьшением содержания глюкозы в крови;
- обеспечивает терморегуляцию организма;
- участвует в регуляции осмотического давления;
- участвует в регуляции деятельности половых желез;
- участвует в формировании оборонительных реакций – оборонительного поведения и бегства.
Пищевое поведение сопровождается поиском пищи. При этом вегетативная реакция несколько иная – увеличивается слюноотделение, повышается моторика и кровоснабжение кишечника, уменьшается мышечный кровоток, так как повышается активность парасимпатической нервной системы.
В гипоталамусе имеются области, отвечающие за те или иные поведенческие реакции, которые перекрываются между собой. При нарушении боковых (латеральных) областей гипоталамуса, где расположены ядра голода и насыщения, возникает афагия (отказ от приема пищи) и гиперфагия (чрезмерное потребление пищи).
В гипоталамусе вырабатывается большое количество медиаторов: адреналин, нордадреналин – возбуждающие медиаторы, глицин, g-аминомасляная кислота – тормозящие медиаторы.
Таким образом, гипоталамус занимает ведущее место в регуляции многих функций организма и прежде всего гомеостаза. Под его контролем находятся функции автономной нервной системы и эндокринных желез.
Эпиталамус.
Эпиталамус - задняя дорсальная структура промежуточного мозга, именуемая иногда “Третьим глазом”.
В состав эпиталамуса входит уздечка, соединительные волокна, задняя расширенная часть мозговой полоски (треугольник поводка) и шишковидное тело (шишковидная железа или эпифиз). Треугольник поводка имеет отношение к подкорковым центрам обоняния. Шишковидная железа принимает участие в регулировании сна, циркадных ритмов, других процессов, в ней вырабатывается гормон мелатонин, является железой внутренней секреции.
Рядом с шишковидной железой располагается уздечка (хабенулярные ядра). Функции данной структуры заключаются в регулировании потребления питательных веществ и воды. Данная структура также содержит в себе афферентные и сенсорные входящие пути, волокна, которые соединяют мозговую полоску с другими частями мозга, участвующими в выбросах серотонина, норадреналина и дофамина.
Шишковидная железа получает входящие нервные волокна из супрахиазменного ядра гипоталамуса, в которое в свою очередь напрямую входят волокна зрительного нерва. Это дает шишковидной железе получать информацию об уровне света для регуляции уровней мелатонина.
Проекция в обратном направлении в сторону хабенулярных ядер, является мозговой полоской таламуса.
Функции эпиталамуса заключаются в подключении лимбической системы к другим частям мозга, обусловлены функциями шишковидной железы, которая вырабатывает мелатонин, осуществляет регуляцию циркадных ритмов, сна, контроль эмоций, контроль потребления питательных веществ и воды организмом. Шишковидная железа принимает участие и в других функциях организма, например, оказывает влияние на половое развитие, ингибируя активность половых желез в период полового созревания.
Эпифиз является железой внутренней секреции (пинеальная железа) и состоит из глиальных элементов и особых клеток пинеалоцитов.
В шишковидном теле у взрослых людей, особенно в старческом возрасте, нередко встречаются причудливой формы отложения, которые придают эпифизу определенное сходство с еловой шишкой, чем и объясняется его название.
Метаталамус представлен латеральным и медиальным коленчатыми телами – парными образованиями.
Метаталамус состоит из серого вещества.
Латеральное коленчатое тело, правое и левое, является подкорковым, первичным центром зрения. К нейронам его ядра подходят нервные волокна зрительного тракта (от сетчатки глаза). Аксоны этих нейронов идут в зрительную зону коры. Медиальные коленчатые тела являются подкорковыми первичными центрами слуха.
III желудочек
3 желудочек головного мозга представляет собой щелевидную полость, ограниченную зрительными буграми таламуса, находящимися в промежуточном мозге. Внутри он имеет мягкую оболочку, которая разветвлена сосудистыми сплетениями и заполнена спинномозговой жидкостью.
Физиологическое значение обширно. Благодаря нему возможен ток ликвора для осуществления омовения. А также происходит циркулирование спинномозговой жидкости.
Третий желудочек внешне напоминает некое кольцо, которое находится между двумя буграми, а внутренняя поверхность содержит вещество серого цвета с подкорковыми центрами. Внизу он контактирует с 4 желудочком.
Кроме того, выделяют определенные функции:
· защита ЦНС;
· выработка ликвора;
· нормализация микроклимата органов центральной нервной системы;
· обмен веществ, предотвращение попадания ненужного к мозгу;
· циркуляция ликвора.
Вкусовая сенсорная система
Вкусовая и обонятельная сенсорные системы позволяют человеку оценивать химический состав пищи и окружающего воздуха. По этой причине их объединяют под названием хемосенсорные системы. Сюда же относятся внутренностные хеморецепторы (каротидного синуса, пищеварительного тракта и другие). Химическая рецепция — одна из наиболее филогенетически древних форм связи организма со средой.
Рецепторный отдел вкусовой сенсорной системы расположен в ротовой полости и представлен вкусовыми рецепторными клетками. Они собраны во вкусовые почки, которые находятся главным образом в сосочках на дорсальной поверхности языка — грибовидных, листовидных и желобовидных. Одиночные вкусовые почки рассеяны в слизистой оболочке мягкого неба, миндалин, задней стенки глотки и надгортанника. У детей область их распространения шире, чем у взрослых; к старости их количество уменьшается. Среди клеток вкусовых почек различают рецепторные, опорные и базальные.
Человек различает четыре основных вкуса (сладкий, соленый, горький, кислый) и несколько дополнительных (металлический, щелочной и т. д.). Рецепция вкусовых веществ становится возможной тогда, когда эти вещества попадают на поверхность языка, растворяются в слюне, проходят через вкусовую пору и достигают апикальной мембраны рецепторных клеток.
Срок жизни рецепторных и опорных клеток невелик — около 10 дней. Их обновление происходит за счет митотического деления клеток в базальной части почки.
В слизистой оболочке передней части языка оканчиваются также волокна тройничного нерва (V пара). Они попадают сюда в составе язычного нерва. По этим волокнам передается тактильная, температурная, болевая и другая чувствительность с поверхности языка, которая дополняет информацию о свойствах стимула в ротовой полости.
Соматосенсорная система
Тело человека покрывает кожный покров. Кожа состоит из поверхностного эпителиального слоя и глубоких слоев (дермы), образованных плотной неоформленной соединительной тканью и подкожной жировой клетчаткой. Кроме того, имеются производные кожи — волосы, ногти, сальные и потовые железы.
Кроме покровной (защитной) кожа выполняет еще целый ряд функций. Она участвует в терморегуляции и выделении, а также несет большое количество рецепторных образований. Эти рецепторы воспринимают информацию о тактильных, болевых, температурных и других раздражениях, приложенных к различным участкам кожи. Другими словами, поверхность нашего тела (сомы) обладает чувствительностью, которая получила название соматической. Для проведения этой импульсации существует несколько проводящих путей, по которым информация передается в различные отделы ЦНС, в том числе в кору больших полушарий. Для каждого вида чувствительности существуют свои проекции, соматотопическая организация которых позволяет определить, к какому участку нашего тела приложено раздражение, какова его сила и модальность (прикосновение, давление, вибрация, температурное или болевое воздействие и т. д.).
Для восприятия этих раздражителей существует несколько видов рецепторных образований. Все они относятся к первичночувствующим, т. е. представляют собой концевые разветвления чувствительных нервных волокон.
Проприорецепция объединяет сенсорные сигналы от скелета и мышц и, следовательно, включает в себя мышечное чувство. Кинестезия — это чувство положения тела и движения конечностей, а также ощущения усилий, силы и тяжести. В ее обеспечении участвуют все рецепторы опорно-двигательного аппарата и кожи.
Мышечные рецепторы — мышечные веретена — служат для определения степени растяжения мышцы. Их особенно много в мышцах, управляющих точными движениями. Эти рецепторы представляют собой образования веретеновидной формы, заключенные в тонкую растяжимую соединительнотканную капсулу.
При растяжении мышцы информация от нервных окончаний поступает в спинной мозг, где часть ее переключается на мотонейроны передних рогов. Их ответная рефлекторная импульсация приводит к сокращению мышцы. Другая часть импульсов переключается на вставочные нейроны и поступает в другие отделы нервной системы (см. ниже).
Мышечные веретена имеют также и эфферентную иннервацию, которая контролирует степень их растяжения. Эфферентные волокна подходят к мышечным веретенам от мотонейронов спинного мозга, но не от тех, что иннервируют саму мышцу, волокна которой называют экстрафузалъными. Однако в некоторых случаях мышечные веретена получают моторную иннервацию по коллатералям от аксонов, идущим к мышцам. Это наблюдается, например, в мышцах глазного яблока.
Кроме рецепторных окончаний, лежащих в самих мышцах и реагирующих на степень их растяжения, существуют рецепторы в местах соединения мышц с сухожилиями. Они носят название сухожильных органов (рецепторов) Голъджи.
Большое количество рецепторных окончаний расположено в суставах. Они чувствительны к растяжению и сжатию, возникающим при движении, и таким образом сигнализируют о положении тела в пространстве и движении отдельных его частей (кинестезия). Свободные нервные окончания могут, кроме того, воспринимать боль.
Глава
Тема: Нервная система и её роль в жизнедеятельности организма
Вопросы:
ü Подразделение нервной системы соответственно развитию, строению и функции на центральную и периферическую, на соматическую и вегетативную.
ü Основные морфологические элементы нервной системы.
ü Нейрон: общая морфология и ультраструктура нейронов, обусловленные его функцией (сома, дендриты, аксон).
ü Оболочки аксона, процесс миелинизации.
ü Немиелинизированные и миелинизированные волокна.
ü Классификация нейронов по форме и функции.
ü Макроглия. Особенности и функции разных видов макроглии (эпендимоциты, астроциты, олигодендроциты).
ü Строение и функции микроглии.
Нервная система состоит из специфической возбудимой ткани — нервной ткани — и представлена центральным и периферическим отделами.
Центральная нервная система находится под защитой костных образований скелета: черепной коробки, в которой расположен головной мозг, а также позвоночника, в спинномозговом канале которого расположен спинной мозг.
К периферической нервной системе относятся нервы и нервные узлы. Выделяют соматическую и вегетативную части периферической нервной системы.
Часть нервной системы, которая регулирует работу скелетных мышц, называют соматической (греч. soma — тело). Посредством соматической нервной системы человек может управлять движениями, произвольно вызывать или прекращать их.
Часть нервной системы, регулирующую деятельность внутренних органов (сердца, желудка, желез и т.д.), называют вегетативной. Работа вегетативной нервной системы не подчиняется воле человека.
Для обозначения взаимного расположения структур нервной системы анатомы пользуются специфическими терминами. Названия направлений основаны на их латинских названиях.
Плоскость, проходящая вдоль по середине тела и делящая его на правую и левую половины, называется сагиттальной (sagittalis).
Плоскость, проходящая вертикально, под прямым углом к сагиттальной, параллельно лбу называется фронтальной (frontis – лоб).
Плоскость, делящая тело на два отдела - верхний и нижний, проходящая перпендикулярно к сагиттальной и фронтальной плоскостям, называется горизонтальной.
Структуры называются:
дорсальными – расположены на спинной стороне тела (dorsalis – спинной),
вентральными – расположены на брюшной стороне тела (ventralis –брюшной),
медиальные – расположены по центру тела (medialis – срединный),
латеральные – расположены вбок от сагитальной плоскости (lateralis –боковой),
апикальные – самые верхние точки (apicalis – верхушечный),
базальные - лежащие в основании (basalis).
Направление к нижней части тела называется каудальное (caudalis –хвостовой), а к головной части – ростральное (rostrum – клюв).
Нейроны (рис. 1) являются возбудимыми клетками нервной системы. В отличие от глиальных клеток они способны возбуждаться (генерировать потенциалы действия) и проводить возбуждение. Нейроны высокоспециализированные клетки и в течение жизни не делятся.
Рис. 1. Нейрон:
1 — сома (тело) нейрона; 2 — дендрит; 3 — тело Швановской клетки; 4 — миелинизированный аксон; 5 — коллатераль аксона; 6 — терминаль аксона; 7 — аксонный холмик; 8 — синапсы на теле нейрона
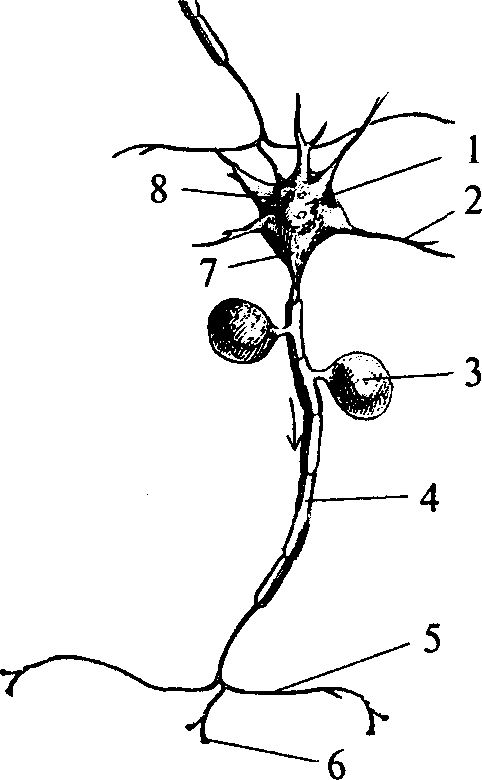
НЕЙРОН – это отдельная нервная клетка, строительный блок мозга. Она передает нервные импульсы по единственному длинному волокну (аксону) и получает их по многочисленным коротким волокнам (дендритам).
Хотя нейроны, или нервные клетки, имеют те же самые гены, то же общее строение и тот же биохимический аппарат, что и другие клетки, они обладают и уникальными особенностями, которые делают функцию мозга совершенно отличной от функции, скажем, печени. Важными особенностями нейронов являются характерная форма, способность наружной мембраны генерировать нервные импульсы и наличие уникальной структуры – синапса, служащего для передачи информации от одного нейрона другому.
Большинству нейронов присущи определенные структурные особенности, позволяющие выделить три области клетки: клеточное тело, дендриты и аксон. Тело содержит ядро и биохимический аппарат синтеза ферментов и других молекул, необходимых для жизнедеятельности клетки. Обычно тело нейрона имеет приблизительно сферическую или пирамидальную форму. Дендриты представляют собой тонкие трубчатые выросты, которые многократно делятся и образуют ветвистое дерево вокруг тела клетки. Они создают ту основную физическую поверхность, на которую поступают идущие к данному нейрону сигналы. Аксон тянется далеко от тела клетки и служит той линией связи, по которой сигналы, генерируемые в теле данной клетки, могут передаваться на большие расстояния в другие части мозга и остальную нервную систему.
Функционирование мозга связано с движением потоков информации по сложным цепям, состоящим из нейронных сетей. Информация передается от одной клетки к другой в специализированных местах контакта – синапсах. Типичный нейрон может иметь от 1000 до 10000 синапсов и получать информацию от 1000 других нейронов. Хотя в своем большинстве синапсы образуются между аксонами одной клетки и дендритами другой, существуют и иные типы синаптических контактов: между аксоном и аксоном, между дендритом и дендритом и между аксоном и телом клетки. В области синапса аксон обычно расширяется, образуя на конце пресинаптическую бляшку, которая является передающей информацию частью контакта.
Если расстояние между мембранами нейронов не превышает 2—4 нм или они контактируют между собой, то такой синапс является электрическим, поскольку подобное соединение обеспечивает низкоомную электрическую связь между этими клетками, позволяющую электрическому потенциалу непосредственно или электротонически передаваться от клетки к клетке. Доля электрических синапсов в ЦНС позвоночных очень мала.
Чаще всего мембраны нейронов расположены в непосредственной близости друг к другу и разделены обычным межклеточным пространством (щелью шириной примерно 20 нм) — смежное соединение. Такая смежность мембран облегчает перемещение из одной клетки в межклеточную щель химических веществ (ионов, метаболитов нейронов), которые оказывают влияние как на ту же самую клетку, так и на отростки соседних нейронов. Эти соединения нейронов относят к химическим синапсам.
Концевая бляшка содержит мелкие сферические образования, называемые синаптическими пузырьками, каждый из которых содержит несколько тысяч молекул химического медиатора. По прибытии в пресинаптическое окончание нервного импульса некоторые из пузырьков выбрасывают свое содержимое в узкую щель, отделяющую бляшку от мембраны дендрита другой клетки, предназначенного для приема таких химических сигналов. Таким образом, информация передается от одного нейрона к другому с помощью некоторого посредника или медиатора.
До 50-х годов XX столетия к медиаторам относили две группы низкомолекулярных соединений: амины (ацетилхолин, адреналин, норадреналин, серотонин, дофамин) и аминокислоты (гамма-аминомасляная кислота, глутамат, аспартат, глицин). Позже было показано, что специфическую группу медиаторов составляют нейропептиды, которые могут выступать также и в качестве нейромодуляторов (веществ, изменяющих величину ответа нейрона на стимул).
В настоящее время известно, что нейрон может синтезировать и выделять несколько нейромедиаторов (сосуществующие медиаторы). Такое представление о химическом кодировании вошло в основу принципа множественности химических синапсов. Нейроны обладают нейромедиаторной пластичностью, т.е. способны менять основной медиатор в процессе развития. Сочетание медиаторов может быть неодинаковым для разных синапсов.
Импульсация нейрона отражает активацию воздействующими нейронами сотен синапсов. Некоторые синапсы являются возбуждающими, т.е. они способствуют генерированию импульсов, тогда как другие – тормозные – способны аннулировать действие сигналов, которые в их отсутствие могли бы возбудить постсинаптический нейрон.
В аксонах, имеющих миелиновую оболочку, распространение нервного импульса происходит путем его перескакивания от перехвата к перехвату, где внеклеточная жидкость оказывается в непосредственном контакте с клеточной мембраной.
Нейроны способны выполнять свою функцию только благодаря тому, что их наружная мембрана обладает особыми свойствами. Мембрана аксона по всей его длине специализирована для проведения электрического импульса. Мембрана аксонных окончаний способна выделять медиатор, а мембрана дендритов реагирует на медиатор.
Классификация нейронов.
По числу отростков.
1. Униполярные нейроны имеют 1 отросток. По мнению большинства исследователей, такие нейроны не встречаются в нервной системе млекопитающих и человека.
2. Биполярные нейроны – имеют 2 отростка: аксон и дендрит. Разновидностью биполярных нейронов являются псевдоуниполярные нейроны спинномозговых ганглиев, где оба отростка (аксон и дендрит) отходят от единого выроста клеточного тела.
3. Мультиполярные нейроны – имеют один аксон и несколько дендритов. Их можно выделить в любом отделе нервной системы.
По форме.
Веретеновидные, грушевидные, пирамидные, полигональные. Такой подход лежит в основе изучения цитоархитектоники мозга.
По выполняемой функции.
1. Чувствительный (афферентный) – помогающий воспринимать внешние раздражители (стимулы).
2. Ассоциативный (вставочный интернейрон).
3. Двигательные (эфферентные) – вызывающие сокращения и движения. Именно эти нейроны получили наименование «мотонейроны», т.е. двигательные нейроны, сконцентрированные в двигательных ядрах передних рогов спинного мозга и стволовой части головного мозга.
Дата: 2018-11-18, просмотров: 495.