Министерство здравоохранения Свердловской области
Фармацевтический филиал ГБОУ СПО Свердловский областной медицинский колледж
КРАТКИЙ КУРС ЛЕКЦИЙ ПО БОТАНИКЕ
Для студентов очной и очно-заочной формы обучения
Екатеринбург
Г.
Министерство здравоохранения Свердловской области
Фармацевтический филиал ГБОУ СПО Свердловский областной медицинский колледж
КРАТКИЙ КУРС ЛЕКЦИЙ ПО БОТАНИКЕ
Для студентов очной и очно-заочной формы обучения
Екатеринбург, 2012 г.
Автор: Голубева Н.Д. – преподаватель биологии высшей категории.
Рецензент: Федосеева Г.П. – к.б.н., старший научный сотрудник, директор Ботанического сада.
«Краткий курс лекций по ботанике» составлен в соответствии с программой, содержит несколько разделов и предназначен для студентов фармацевтического колледжа очно-заочной формы обучения.
В нем в сжатой форме изложен основной материал по анатомии, морфологии и систематике растений, приведено 100 рисунков по этим разделам. Материал тесно связан со специальной дисциплиной – фармакогнозией. Екатеринбург: Фармацевтический филиал ГБОУ СПО Свердловский областной медицинский колледж, 2012 – 97 с.
«Краткий курс лекций по ботанике» утвержден на заседании кафедры ОПД.
Протокол №12 от 02 июля 2012 года
Фармацевтический филиал ГБОУ СПО Свердловский областной медицинский колледж, 2012.
Введение
Курс ботаники в среднем фармацевтическом образовании является общетеоретической дисциплиной, которая готовит студентов к изучению специального курса – фармакогнозии.
Данный курс лекций по ботанике составлен в соответствии с программой (Москва, 2002). В нем приводятся сведения об особенностях анатомии, морфологии и систематики основных ботанических семейств, изучаемых в курсе ботаники и фармакогнозии. Многие разделы иллюстрированы рисунками.
Анатомия растений изучает строение тканей и органов на клеточном уровне. Знания по анатомии растений необходимы фармацевту для проведения микроскопического анализа измельченного лекарственного растительного сырья с целью установления его подлинности.
Морфология растений рассматривает внешние формы органов, знания чего необходимы фармацевту для проведения макроскопического анализа лекарственного растительного сырья.
Систематика растений знакомит студентов с лекарственными растениями из разных семейств. В практической работе фармацевта важно умение отличать одни виды растений от других по морфологическим признакам, а так же знание диагностических признаков разных семейств.
СОДЕРЖАНИЕ
Анатомия и морфология растений. 7
Глава 1. Клетка. Строение растительной клетки. 7
1.1. Мембранные органоиды растительной клетки. 7
1.2. Немембранные органоиды растительной клетки. 9
1.3. Ядро. 9
1.4. Производные протопласта – включения. 10
1.5. Вакуоли. 12
1.6. Клеточная оболочка. 13
Глава 2. Растительные ткани. 14
2.1. Образовательные ткани - меристемы.. 14
2.2. Покровные ткани. 15
2.3. Механические ткани. 18
2.4. Основные ткани - паренхима. 20
2.5. Проводящие ткани. 23
2.6. Выделительные (секреторные) ткани. 27
Глава 3. Вегетативные органы высших растений. 31
3.1. Морфология корня. 31
3.2. Видоизменения корня. 32
3.3. Морфология стебля. 34
3.4. Видоизменения побега. 37
3.5. Морфология листа. 39
3.6. Сложные листья. 44
3.7. Расположение листьев на стебле. 46
Глава 4. Генеративные органы растений. 47
4.1. Морфология цветка. 47
4.2. Морфология соцветий. 50
4.3. Морфология плодов. 54
4.4. Морфология семян. 58
Систематика растений. 60
Глава 5. Понятие о систематике. Основные систематические единицы.. 60
5.1. Систематика низших и высших растений. 62
5.2. Грибы – Mycota. 63
5.3. Растения – Plantae. 64
5.4. Подцарство Настоящие водоросли – Phycobionta. 65
5.5. Царство растения – Plantae. Подцарство Настоящие водоросли – Phycobionta. Отдел Бурые водоросли – Phaeophyta. 66
5.6. Подцарство Высшие растения – Embryophyta. 67
5.7. Отдел плауновидные – Lycopodiophyta. 68
5.8. Отдел хвощевидные – Eguisetophyta. 69
5.9. Отдел папоротниковидные – Polypodiophyta. 71
Глава 6. Семенные растения. 73
6.1. Отдел голосеменные – Gymnospermae. 73
6.2. Отдел цветковые или покрытосеменные растения – Magnoliophyta или Angiospermae. 76
6.3. Класс двудольные – Dicotyledones. Порядок лютикоцветные – Ranunculales. Семейство лютиковые – Ranunculaceae. 77
6.4. Порядок Маковые – Papaverales. Семейство Маковые – Papaveraceae. 78
6.5. Порядок Гречишные – Polygonales. Семейство Гречишные – Polygonaceae. 79
6.6. Порядок каперсовые – Capparales. Семейство Капустные – Brassicaceae. 80
6.7. Порядок Розоцветные – Rosales. Семейство Розоцветные – Rosaceae. 81
6.8. Порядок Бобовые – Fabales. Семейство Бобовые – Fabaceae. 84
6.9. Порядок Аралиевые – Arales. Семейство Сельдерейные – Apiaceae. 85
6.10. Порядок Губоцветные – Lamiales. Семейство Яснотковые – Lamiaceae. 87
6.11. Порядок Пасленовые – Solanales. Семейство Пасленовые – Solanaceae. 89
6.12. Порядок Астровые – Asterales. Семейство Астровые – Asteraceae. 90
6.13. Класс Однодолные – Monocotyledones. Подкласс Лилииды – Liliidae. Порядок Лилейные – Liliales. Cемейство Лилейные – Liliaceae. 94
6.14. Порядок Злаки – Poales. Семейство Мятликовые – Poaceae. 95
Использованная литература. 97
Анатомия и морфология растений
Ядро
Ядро – это обязательный органоид клеток, имеет шаровидную форму. В молодых клетках ядро расположено в центре, в зрелых – оттеснено вакуолью к стенке клетки. В его состав входят ядрышко, хроматин (хромосомы ) и ядерный сок. Диаметр ядра 5-20 мкм. Ядро – это носитель наследственной информации, управляет процессами метаболизма через белки – ферменты, снабжает клетку РНК, участвует в делении клетки и оплодотворении, образует некоторые органоиды клетки. Покрыто двойной ЦПМ с порами, через которые из ядра в цитоплазму выходят молекулы и–РНК, т–РНК, рибосомы (субъединицы), а в ядро поступают нуклеотиды, белки, ферменты, АТФ, вода, ионы и т.д. Через ЭПС ядро связано со своей клеткой и другими клетками. Внутри ядра находится ядерный сок (кариолимфа), сходный по структуре и составу с гиалоплазмой. Ядерный сок заполняет пространство между ядерными структурами, участвует в транспорте веществ, нуклеиновых кислот, субъединиц рибосом.
Ядрышко - имеет шаровидную форму, состоит из белка и р–РНК. Участвует в формировании половинок рибосом из р-РНК и белка.
Хромосомы (хроматин)- состоят из молекул ДНК и белковой обкладки, являются носителями наследственной информации.
Рис. 3. Липидные капли (ЛК) и митохондрии (М) в дифференцирующейся трахеиде ели под электронным микроскопом (схемат.), увел. 12000.
Липиды – в виде жировых капель (Рис. 3) обычно располагаются в гиалоплазме и встречаются практически во всех растительных клетках. Особенно много их в семенах подсолнечника, льна, горчицы и др.
Из группы экскреторных веществ в растительных клетках чаще всего встречается щавелево-кислый кальций, или оксалат кальция (СаС2О4). Выделяемая в процессе дыхания клетки щавелевая кислота нейтрализуется ионами кальция, поступающими в растения из почвенного раствора. Форма отложений оксалата кальция специфична у разных растений: одиночные кристаллы (кубические, призматические, стилоиды), рафиды (игловидные кристаллы, лежащие в клетке параллельно друг другу виде пучка), друзы (сростки пирамидальных кристаллов), кристаллический песок (скопление множества мелких кристаллов) (Рис. 4). Кристаллы образуются в тканях и органах, которые периодически сбрасываются растениями (листья, кора). Форма кристаллов используется для микродиагностики лекарственного растительного сырья (ЛРС).
Для диагностики ЛРС используется также цистолиты, которые состоят из карбоната кальция или кремнезема и представляют собой выросты клеточной оболочки внутрь клетки (Рис. 4).
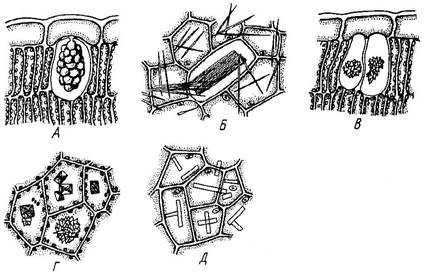
Рис. 4. Кристаллы и скопления минеральных солей в клетках. А – цистолит в клетке эпидермы листа инжира; Б – рафиды в клетках листа традесканции; В – друзы в клетках палисадной ткани листа инжира; Г – друзы и одиночные кристаллы в клетках черешка бегонии; Д – одиночные кристаллы в клетках эпидермы чешуи луковицы лука.
Вакуоли
Вакуоли - производные ЭПС, ограниченные мембраной и заполненные клеточным соком. Клеточный сок – это водный раствор органических и неорганических веществ. Большинство органических веществ клеточного сока - продукты метаболизма протопласта. Наиболее обычны различные углеводы, играющие роль запасных питательных веществ (глюкоза, фруктоза, сахароза, инулин и др.), а также органические кислоты (щавелевая, яблочная, лимонная и др.). В вакуолях часто концентрируются вторичные метаболиты: флавоноиды, антоцианы, танниды, азотосодержащие вещества – алкалоиды. Многие из них используются для изготовления лекарственных препаратов.
Функции вакуолей: регуляция водно-солевого обмена; поддержание тургорного давления внутриклеточной жидкости; накопление запасных веществ и «захоронение» отбросов, т.е. конечных продуктов метаболизма клетки.
Клеточная оболочка
Клеточная оболочка - располагается по периферии клетки, придает ей прочность, форму, защищает протопласт, является продуктом жизнедеятельности протопласта. Состоит из клетчатки (целлюлозы), гемицеллюлозы, пектиновых веществ. Поры – это неутолщенные места клеточной оболочки. Они связывают соседние клетки друг с другом. В процессе жизнедеятельности клеток наступают вторичные изменения клеточной стенки:
1. Одревеснение – пропитывание клеточной стенки лигнином, что повышает ее твердость и прочность, понижает пластичность и способность расти. Процесс отложения лигнина – одревеснение – наблюдается в разных органах и тканях.
2. Опробковение – пропитывание оболочки суберином. Оболочка клетки становится непроницаема для воды и газов, а клетка быстро отмирает. Выполняет защитную функцию.
3. Кутинизация – это наслоение на клеточную стенку кутина (жироподобное вещество). Предохраняет органы растения от избыточного испарения, защищает от проникновения паразитов, механических повреждений.
4. Минерализация – инкрустация стенок поверхностных клеток углекислым кальцием или кремнеземом (хвощи, осоки, злаки). Осевые органы становятся более прочными; минерализация защищает растения от поедания животными и паразитами.
5. Ослизнение – образование слизей и камедей в стенках клеток. Они создают влажную среду вокруг зародыша семени.
Оболочки делящихся и растущих клеток называют первичными. Они богаты водой, целлюлозы в них не более 30%. В процессе жизнедеятельности толщина клеточной стенки может увеличиваться, а объем полости клетки сократиться. Такая оболочка называется вторичной. В ней содержится меньше воды, а количество целлюлозы достигает 40-50% от массы сухого вещества. Вторичная оболочка иногда откладывается неравномерно: в виде колец, спиралей, что позволяет клеткам сохранять способность к растяжению в длину (кольчатые и спиральные сосуды) (Рис. 15).
Глава 2. Растительные ткани
Все органы растений имеют клеточное строение. Группа клеток, сходных по строению, функции и имеющих общее происхождение, называется тканями. В зависимости от основной функции их делят на:
1. образовательные (меристематические);
2. покровные;
3. механические;
4. основные;
5. проводящие;
6. выделительные (секреторные).
Образовательные ткани - меристемы
Расположены в зонах роста стеблей, листьев и корней. Меристема состоит из живых, плотно расположенных, мелких, паренхимных клеток, с тонкими целлюлозными оболочками, густой цитоплазмой, крупным ядром, без пластид и вакуолей. Основная функция меристемы - образование клеток постоянных тканей путём митоза и их дифференциация (Рис. 5).
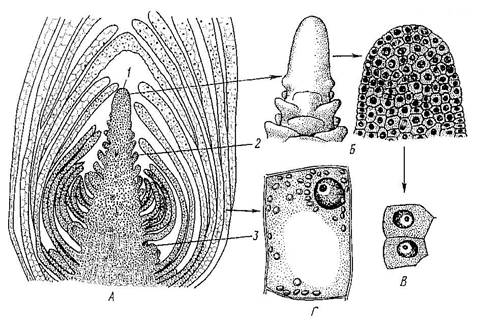
Рис. 5. Верхушечная меристема побега элодеи. А – продольный срез; Б – конус нарастания (внешний вид и продольный срез); В – клетки первичной меристемы; Г – паренхимная клетка сформировавшегося листа:
Покровные ткани
Покровные ткани растений представлены первичными и вторичными тканями.
Первичные покровные ткани состоят у растений из эпидермы. (Рис. 6).
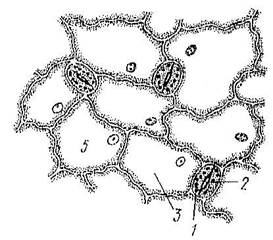
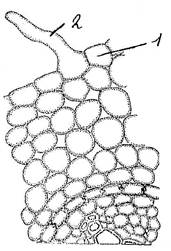

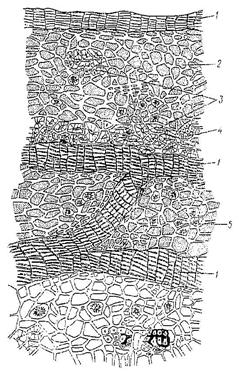
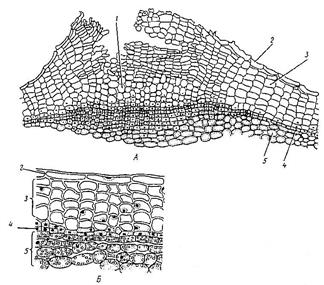

Рис. 6. Покровные ткани.
Эпидерма стебля и листа и эпидерма (ризодерма) корня является первичными покровными тканями. У большинства растений эпидерма состоит из одного слоя живых, плотно примыкающих друг к другу клеток, лишённых хлоропластов. Оболочки клеток эпидермы подвергаются кутинизации, заключающейся в том, что на наружную стенку клетки протопластом откладывается слой жироподобного вещества – кутина. Этот слой называется кутикулой. У растений засушливых мест кутикула толще, чем у растений, обитающих при среднем увлажнении. У водных и тенелюбивых растений кутикулы может и не быть. Характерной особенностью эпидермы является наличие устьиц, состоящих из двух замыкающих клеток и устьичной щели между ними. Через них осуществляется транспирация (выделение паров воды) и дыхание.
Характер клеток эпидермы различен. У двудольных растений боковые стенки клеток эпидермы извилистые, у однодольных растений (на листьях) - сильно вытянутые в длину (Рис. 7).
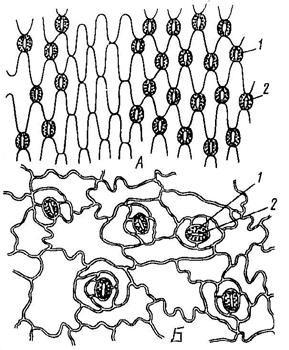
Рис. 7. Эпидерма хлорофитума – однодольное растение (А) и шелковицы белой – двудольное растение (Б): 1 – замыкающая клетка; 2 – устьичная щель.
На некоторых клетках эпидермы образуются выросты. Они могут быть представлены простыми волосками и головчатыми (железистыми) волосками. Волоски бывают одноклеточные или многоклеточные различной формы (Рис. 8).
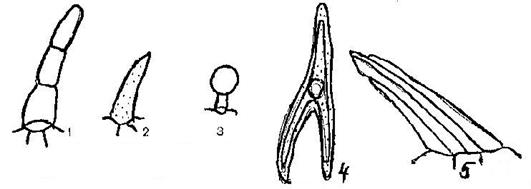
Рис. 8. Различные виды волосков:
1 – простой многоклеточный; 2 – простой одноклеточный; 3 – головчатый;
Трехконечный; 5 – пучковый.
В фармакогнозии особенности волосков, строение эпидермы используют для диагностики лекарственного растительного сырья (ЛРС).
К первичным покровным тканям относится и эпидерма (ризодерма), покрывающая корни в зоне всасывания (Рис. 6). Клетки ризодермы тонкостенные, лишены кутикулы, устьиц нет. В результате выпячивания наружной стенки клеток ризодермы образуются корневые волоски длиной 1-2 мм. Он поглощают воду и минеральные соли из почвенного раствора.
Вторичной покровной тканью являются перидерма и корка, образующиеся из вторичной меристемы - феллогена (пробкового камбия). Клетки феллогена при делении параллельно поверхности органа откладывают наружу слои клеток пробки, внутрь - клетки феллодермы. Пробка, феллоген, феллодерма образуют перидерму (Рис. 6). Стенки клеток пробки пропитаны суберином, который не пропускает газы и воду. Протопласт этих клеток отмирает, а связь тканей с внешней средой происходит через чечевички - отверстия, прикрытые рыхлой тканью из паренхимных клеток с большими межклетниками. Чечевички используют для диагностики коры (дуб, калина), заготавливаемой в качестве ЛРС.
Корка - со временем клетки наружных слоев перидермы и располагающихся под ними тканей отмирают, образуя корку (Рис. 6). Она покрывает стволы, многолетние ветви и корни деревьев. Надёжно предохраняет стволы деревьев от механических повреждений, низовых пожаров, смены температур, проникновения микробов и паразитов.
Механические ткани
Механические ткани встречаются во всех органах растений. Это опорные ткани, обеспечивающие прочность органов растения, их способность противостоять нагрузкам на растяжение, сжатие и изгиб. Прочность и твёрдость клеток, составляющих механическую ткань, достигается утолщением и часто одревеснением их оболочек с пропитыванием лигнином. В зависимости от формы клеток, химического состава клеточных стенок и способа их утолщения механические ткани делят на три типа: колленхиму, склеренхиму, склереиды (Рис. 9).
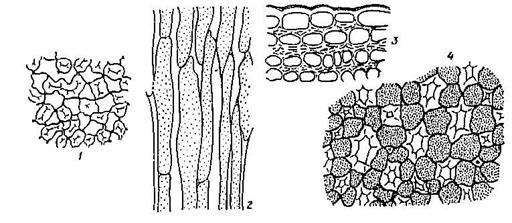
Рис. 9.1. Колленхима:
1, 2 – уголковая колленхима в стебле лебеды (на поперечном и продольном разрезах); 3 – пластинчатая колленхима в побеге бузины; 4 – рыхлая колленхима с межклетниками в черешке листа белокопытника.


Колленхима - живая ткань, располагается под эпидермой и характерна для двудольных растений. Неравномерные целлюлозные утолщения клеточных стенок могут располагаться по уголкам клеток (уголковая колленхима), или клеточная оболочка утолщена равномерно (пластинчатая колленхима), или утолщены стенки, примыкающие к крупным межклетниками (рыхлая колленхима) (Рис. 9.1).
Склеренхима - располагается в растениях глубже, чем колленхима; она есть как у двудольных, так и у однодольных растений. Клетки склеренхимы прозенхимные, мёртвые, с равномерно утолщёнными одревесневшими оболочками, полость клетки мала и узка. Одревеснение (пропитывание целлюлозной оболочки лигнином) повышает прочность склеренхимы у древесинных волокон - либриформа. Лишь иногда клетки склеренхимы не одревесневают - лубяные волокна льна. Поры в оболочках склеренхимы простые, немногочисленные. Лубяные волокна расположены во флоэме (лубе) и нередко достигают значительной длины (у прядильных растений). Древесинные волокна, или волокна либриформа входят в состав ксилемы (древесины), они короче лубяных волокон и их стенки всегда одревесневают.
Склереиды - это мёртвые паренхимные клетки с равномерно и очень сильно утолщёнными одревесневшими оболочками. Возникают из клеток основной паренхимы, разнообразные по форме. Наиболее часто встречаются каменистые клетки (Рис. 9.3). Они располагается группами в мякоти плодов груши, айвы, образуют эпикарпий плодов костянок у персика, черёмухи, абрикоса и др. Склереиды могут располагаться поодиночке - клетки идиобласты (Рис. 10).
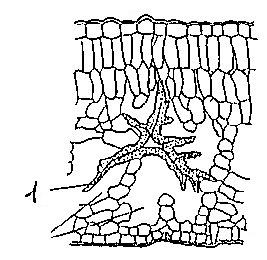
Рис. 10. Идиобласт (1) в листе троходендрона.
В мезофилле листьев некоторых растений (сем. Чайные) встречаются склереиды звёздчатой формы - астросклереиды. Считается, что склереиды противостоят сдавливанию сочных тканей.
Основные ткани - паренхима
Основные ткани создают массу тела травянистых растений, занимают пространство между другими постоянными тканями. Это относительно мало специализированные ткани. Клетки живые, паренхимные, разнообразные по форме (округлые, эллиптические, цилиндрические и др.). Клеточные оболочки тонкие, целлюлозные, цитоплазма этих клеток чаще расположена постенно. Клетки паренхимы располагаются рыхло, т.е. имеют межклетники. В связи с выполнением главной функции выделяют несколько разновидностей основных тканей: ассимиляционную, запасающую, водоносную и воздухоносную паренхиму.
Ассимиляционная паренхима (хлоренхима) - состоит из живых паренхимных клеток, содержащих хлоропласты, в которых протекает фотосинтез. Основная масса хлоренхимы сосредоточена в листьях. Меньшая часть - в молодых зелёных стеблях (Рис. 11).
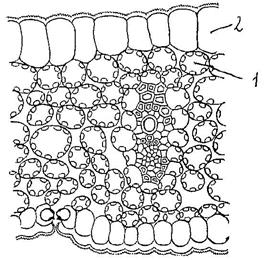
Рис. 11. Поперечный срез листа хлорофитума: 1 – хлоренхима; 2 – эпидермис.
Запасающая паренхима состоит из живых тонкостенных паренхимных, наполненных отложившимися в них запасными питательными веществами -крахмалом, сахаром, инулином, белком, жирами и др. Кроме того, в них иногда запасаются алкалоиды, гликозиды, дубильные вещества, каучук и др. Запасающие ткани имеются в самых различных органах: это эндосперм или зародыш семян, клубни, луковицы, утолщения корней, сердцевина стеблей и др. (Рис. 12).
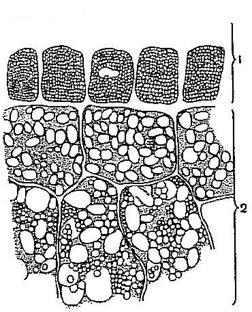
Рис. 12. Разрез через запасающую ткань зерновки пшеницы.
Рис. 13-А. Схема поперечного разреза через лист алоэ: ВТ – водоносная ткань, ПП – проводящие пучки, Хл – хлоренхима, Э – эпидерма.
Воздухоносная паренхима (аэренхима) хорошо развита в разных органах водных и болотных растений и способствует газообмену (Рис. 13-Б), в некоторых случаях снабжает листья диоксидом углерода для обеспечения плавучести растений. Аэренхима имеет большие воздухоносные полости, возникающие как крупные межклетники или путём отмирания и разрушения ткани.
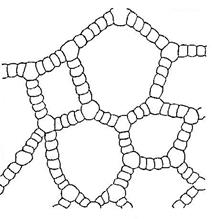
Рис. 13-Б. Аэренхима в черешке листа кувшинки.
Проводящие ткани
Проводящие ткани проводят по растению воду и растворённые в ней органические и минеральные вещества. Вместе с механическими и основными тканями они образуют комплексы: флоэму (луб) и ксилему (древесину).
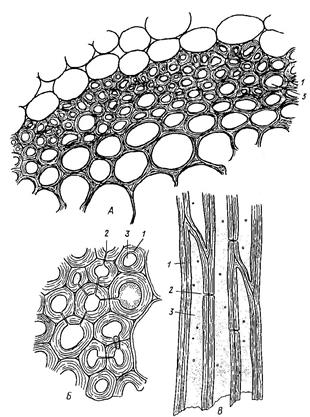
Флоэма осуществляет нисходящий, ассимиляционный ток органических веществ от листьев к корням по проводящим элементам: ситовидным клеткам. Они имеют прозенхимную форму, ситовидные поля и ядро. Это основной проводящий элемент флоэмы у всех высших растений, исключая покрытосеменные. Ситовидные трубки (Рис. 14) состоят из живых отдельных клеток-члеников, располагающихся друг над другом, не имеют ядер.
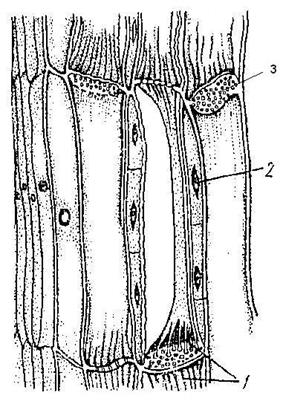
Рис. 14. Ситовидные трубки (1), клетки-спутницы (2), ситовидная пластинка (3).
Ситовидные поля двух смежных члеников образуют ситовидную пластинку (Рис. 14.3). Сбоку от ситовидных трубок располагается клетки-спутницы, осуществляющие ближний радиальный транспорт веществ. Они имеют тонкую целлюлозную оболочку, цитоплазму, ядро. Во флоэме располагается механическая ткань склеренхима - лубяные волокна и запасающая ткань - лубяная паренхима.
Ксилема осуществляет восходящий или транспирационный ток воды и растворённых в ней минеральных солей от корней к листьям по проводящим элементам: трахеидам и трахеям (сосудам). В состав ксилемы входят проводящие элементы, запасающие и опорные. Трахеиды - мёртвые прозенхимные клетки, суженные на концах, их длина 1-4 мм. Стенки трахеид толстые, одревесневшие, несут простые или окаймленные поры. Сосуды - это полые трубки, состоящие из отдельных члеников, располагающихся друг над другом. Между члениками одного и того же сосуда имеются сквозные отверстия - перфорации, благодаря которым вдоль всего сосуда свободно осуществляется ток жидкости.
Помимо первичной оболочки, сосуды и трахеиды имеют вторичные утолщения в форме, колец, спиралей и др. В связи с этим различают кольчатые, спиральные, лестничные, пористые сосуды и трахеиды (Рис. 15).
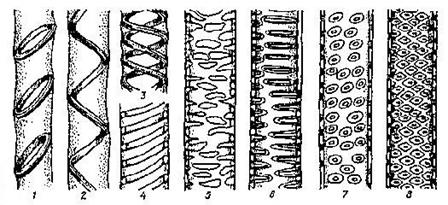
Рис. 15. Типы утолщения и поровости боковых стенок трахеальных элементов: 1 – кольчатое, 2-4 – спиральные, 5 – сетчатое утолщение, 6 – лестничная, 7 – супротивная, 8 – очередная поровость.
Помимо трахеальных элементов ксилема включает живые паренхимные клетки - древесинную паренхиму. По ней отчасти осуществляется ближний транспорт веществ и запасаются питательные вещества. Механическими элементами ксилемы являются древесинные волокна – либриформ (см. «Механические такни»).
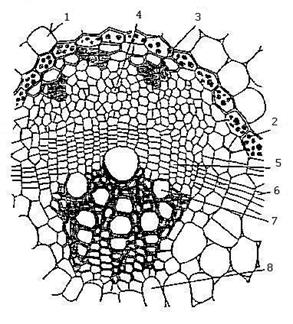
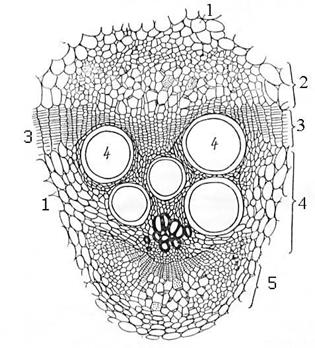
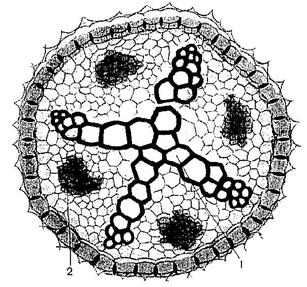
Элементы флоэмы и ксилемы в стеблях и листьях растений формируют проводящие пучки. Если флоэма и ксилема расположены бок о бок, то эти пучки называются коллатеральные (бокобочные) проводящие пучки (Рис. 16.1 и 16.2). У двудольных растений между флоэмой и ксилемой пучка имеется вторичная образовательная ткань - камбий. Такой проводящий пучок называется открытым (Рис. 16.2), у однодольных растений в пучках камбия нет - это закрытые пучки (Рис. 16.1). У некоторых растений (сем. Тыквенные, Пасленовые и др.) к участку ксилемы в открытом коллатеральном пучке примыкает ещё одна группа клеток флоэмы. Такие пучки называются биколлатеральные-двубокобочные пучки (Рис. 16.3).
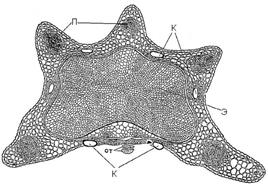
В корневищах и стеблях растений встречаются концентрические проводящие пучки. При этом либо флоэма окружает ксилему (центроксилемный пучок), либо, наоборот ксилема окружает флоэму (центрофлоэмные пучки) (Рис. 16.4). Центрофлоэмные пучки найдены в стеблях и корневищах двудольных растений (ревень, щавель, бегония) и однодольных (лилейные). Центроксилемные пучки встречаются в корневищах папоротников.
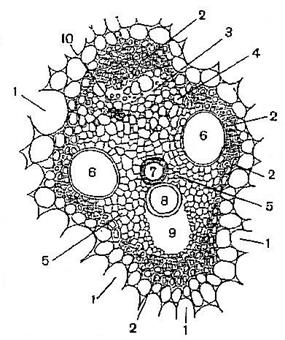
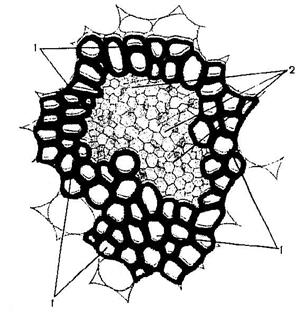
Для корней покрытосеменных растений в зоне всасывания и корней голосеменных растений характерны радиальные проводящие пучки, в которых ксилема расходится лучами от центра, а флоэма располагается между лучами ксилемы. Радиальные и концентрические пучки всегда закрытые (Рис. 16.5).
|
|

|

Рис. 16. Радиальные и концентрические пучки.
Рис. 17. Железистые волоски (1), эфирномасличная железка: вид сверху (2), вид сбоку (3).
Желёзки - это структуры с многоклеточной секретирующей головкой, располагающейся на короткой ножке (Рис. 17). Желёзки, в основном, выделяют эфирные масла. Они характерны для растений семейств яснотковые и астровые, служат диагностическим признаком при микроскопии ЛРС.
Нектарники устроены очень сложно, это железистые образования, выделяющие нектар. Они располагаются в цветках.
Гидатоды - водяные устьица - это сложная система клеток, выделяющая водно-солевые растворы в виде капель. Гидатоды располагаются по краям или на верхушках листьев.
К выделительным тканям внутренней секреции относят вместилища выделений, смоляные ходы, эфирномасличные каналы, млечники.
Вместилища выделений представляют собой полости различной формы, располагающиеся в толще других тканей. По происхождению бывают лизигенные и схизогенные.
Лизигенные вместилища образуются в результате распада - лизиса клеток после накопления секрета в межклетнике (Рис. 18).
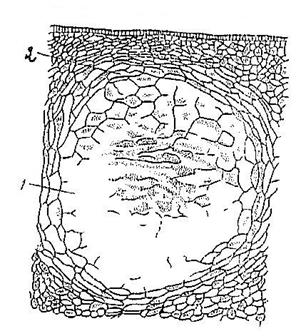
Рис. 18. Лизигенное вместилище в наружной части плода мандарина: 1 – полость вместилища, 2 - выстилающие клетки.
Встречаются в перикарпии плодов цитрусовых.
Схизогенные вместилища образуются из межклетников, путём расхождения клеток, тесно примыкавших друг к другу. Секрет выделяется в полость межклетников живыми эпителиальными (выделительными) клетками, выстилающими полость изнутри. Чаще всего в них накапливаются слизь, реже эфирные масла и смолы. К ним относятся: смоляные ходы и эфирномасличные каналы, млечники.
Смоляные ходы и эфирномасличные каналы (Рис. 19) всегда образуются схизогенно и изнутри выстланы секретирующими эпителиальными клетками.
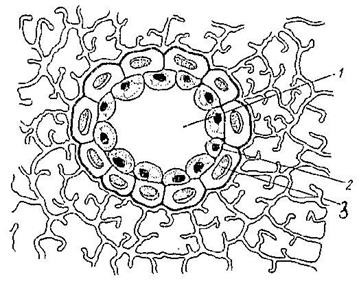
Рис. 19. Схизогенное вместилище в хвое сосны: 1 – полость вместилища, 2 – клетки эпителия (выстилающие клетки), 3 – механические клетки, выстилающее вместилище.
От схизогенных вместилищ они отличаются формой. Ходы и каналы более или менее вытянуты и могут ветвиться. Смоляные ходы содержат смолу, а эфирномасличные каналы - эфирные масла (в плодах сельдерейных), (Рис. 20).
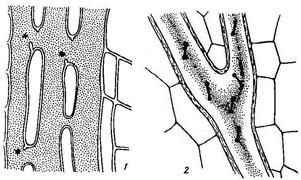
А. Б.
Рис. 20. Эфирномасличные каналы (А) и млечники (Б).
А - Эфирномасличные каналы (к) в семенах фенхеля; п – проводящие пучки; э – эндосперм.
Б – млечники: 1 – членистый млечник (стебель латука); 2 – нечленистый млечник (стебель молочая).
Млечники пронизывают всё растение. В вакуолях млечников находится млечный сок-латекс, представляющий собой эмульсию молочно-белого (одуванчик) или оранжевого цвета (чистотел), содержит воду, углеводы, белки, алкалоиды, танниды и др. Различают членистые и нечленистые млечники. Членистые млечники образуются из нескольких клеток путём разрушения перегородок между ними и слияния многих клеток в сплошную разветвлённую систему (Рис. 20.Б.1). Такие млечники встречаются у маковых, астровых и др. Нечленистые млечники образуются из одной разветвлённой клетки (Рис. 20.Б.2). Млечники участвуют в проведении и запасе питательных веществ.
Морфология корня
Корень – это вегетативный осевой орган растения, имеющий неограниченный рост за счет верхушечной меристемы, закрепляющий растение в субстрате и выполняющий функцию минерального и водного питания. Корень выполняет и другие функции: синтезирует органические вещества; в корнях откладываются запасные питательные вещества (крахмал, инулин и др.); за счет корней происходит связь растений с бактериями и грибами почвы (симбиоз); с помощью корней осуществляется вегетативное размножение растений. Морфологически корень отличается от побега тем, что на нем никогда не возникают листья, а верхушечная меристема прикрыта корневым чехликом.
Совокупность крупных и мелких корней образует корневую систему. Первый корень семенного растения развивается из зародышевого корешка. Он называется главным. У двудольных и голосеменных растений от него отходят боковые корни первого порядка, от них корни второго порядка и т.д. В результате формируется стержневая корневая система (Рис. 21.А) или ее разновидность – ветвистая корневая система.
У однодольных растений зародышевый корешок быстро погибает и главный корень не развивается. Вместо него при основании побега образуются придаточные корни, сходные между собой по размерам. Корневая система, сформированная массой придаточных корней, называется мочковатой (Рис. 21.Б).
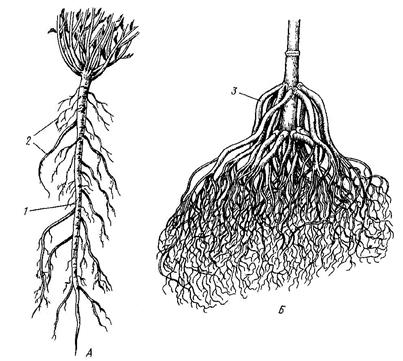
Рис. 21. Корневые системы. А – стержневая, Б – мочковатая:
1 – главный корень, 2 – боковые корни, 3 – придаточные корни (отходят от стебля).
Видоизменения корня
Некоторые корни выполняют особые функции, в связи, с чем меняется их строение. Различают:
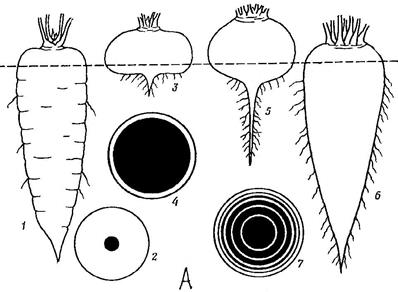
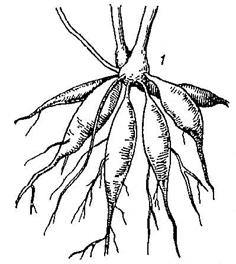
1.Запасающие корни – запасают питательные вещества. При разрастании главного корня образуется корнеплоды(Рис. 22-А) (морковь, репа, свекла), а при разрастаний боковых или придаточных корней образуется корневые клубни (георгины) или корневые шишки (таволга) (Рис. 22-Б).
 Б
Б 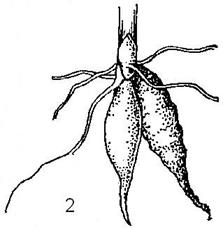
Рис. 22. А – корнеплоды моркови (1, 2), репы (3, 4), свеклы (5, 6, 7).
На поперечных разрезах ксилема показана черной; пунктирной линией обозначена граница стебля и корня.
Б – корневые клубни георгина (1) и любки (2).
2.Втягивающие корни – отходят от луковиц, корневищ и, укорачиваясь, втягивают их в почву, на оптимальную для их сохранения глубину зимой или летом, в засуху (луковичные растения).
3.Шнуровидные корни – это равномерно утолщенные придаточные корни, отходящие от корневища (чемерица).
4.Дыхательные корни – характерны для растений, живущих на бедных кислородом, затопляемых почвах (болотные растения).
5.Корни-подпорки – придаточные корни, отходящие от стволов деревьев на разной высоте и поддерживающие растение.
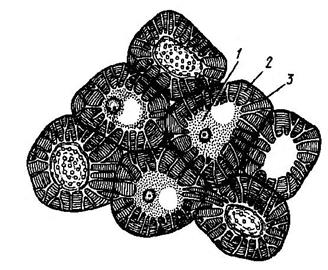
6.Клубеньки – образуются на корнях бобовых растений азотфиксирующими бактериями, которые усваивают азот из воздуха. Благодаря этому почвы обогащаются азотом (Рис. 23).
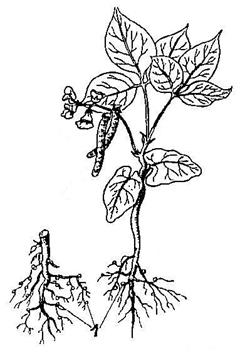
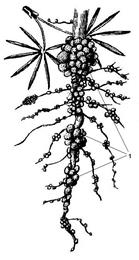
Рис. 23. Клубеньки (1) на корнях бобовых.
7.Микориза – это симбиоз корней высших растений и почвенных грибов. Гифы грибов заменяют корневые волоски и поглощают из почвы воду и минеральные вещества, а высшие растения снабжает грибы органическими веществами.
Морфология стебля
Побег – это вегетативный орган растения, состоит из стебля, листьев и почек, имеет неограниченный рост за счет верхушечной меристемы.
Стебель – осевая часть побега. Участок стебля, от которого отходит лист (или листья), называется узлом. Участки стебля между соседними узлами – междоузлия. На верхушке побега располагается верхушечная почка.
Почка – это зачаточный укороченный побег. За счет нее осуществляется рост побега. Внутренний угол между листом и стеблем называется пазухой листа, а почка, расположенная в ней – пазушной или боковой почкой. Придаточные почки возникают на взрослых частях того или иного органа из внутренних тканей и обеспечивают вегетативное возобновление. Помимо чисто вегетативных почек бывают смешанные (вегетативно-генеративные). Они несут зачаточные листья, цветки и соцветия. Цветочные почки несут только зачаточные цветки или соцветия. Почки окружены чешуями – видоизмененными листьями. Они защищают конус нарастания почки от внешних воздействий. Конус нарастания состоит из верхушечной меристемы, в результате меристематической активности апекса образуются зачатки листьев, зачаточные узлы и междоузлия (Рис. 24).


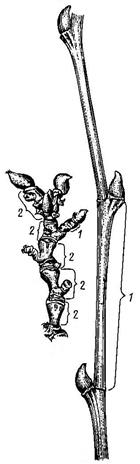
В зависимости от длины междоузлий побеги делят на укороченные (Рис. 25) и удлиненные (Рис. 24). Первые имеют сближенные узлы, вторые – длинные междоузлия и расставленные узлы.
Главный побег в большинстве случаев бывает прямостоячий и занимает вертикальное положение. Иногда главный побег сначала располагается параллельно поверхности почвы, а затем занимает вертикальное положение – приподнимающийся побег (спорыш). Побеги, стелющиеся по земле и не укореняющиеся в узлах, называются лежачие или стелющиеся. Если же стелющиеся побеги укореняются в узлах, то это ползучие побеги (земляника, клевер ползучий). Вьющиеся побеги обвиваются вокруг опоры по часовой стрелке или против (фасоль, вьюны, хмель), а цепляющиеся побеги прикрепляются к опоре при помощи усиков (горох, огурцы) (Рис. 26).
Рис. 26. Стебель: а – прямостоячий, б – приподнимающийся, в – лежачий, г – ползучий, д – цепляющийся, е – въющийся.
В зависимости от формы поперечного сечения стебли бывают:
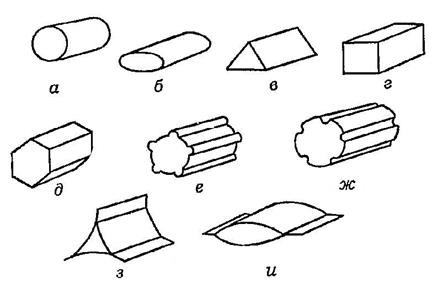
Рис. 27. Стебель в поперечном сечении:
а – округлый, б – сплюснутый, в – 3-гранный, г – 4-гранный, д – многогранный, е – ребристый, ж – бороздчатый, з-и – крылатый.
В процессе эволюции побеги многих растений видоизменились, приспособились к перенесению неблагоприятных условий, запасу питательных веществ и др. Различают надземные и <
Дата: 2016-10-02, просмотров: 647.