• Генетический код – это способ записи информации об аминокислотном составе белка с помощью нуклеотидов
Триплетный
Однозначный
Вырожденный (избыточный)
Существуют нонсенс-кодоны
Неперекрывающийся
Непрерывный
Универсален для всех живых систем
· ТРАНСЛЯЦИЯ - процесс синтеза белка из аминокислот по матрице мРНК на рибосомах в цитоплазме. Аминокислоты соединяются в последовательность, определяемую нуклеотидными основаниями мРНК, и доставляются в рибосому транспортной РНК (тРНК). Каждая аминокислота имеет свою собственную тРНК, у которой есть область (антикодон), комплементарная кодону на мРНК.
В рибосоме имеются три различных участка, с которыми связывается РНК: один для мРНК и два – для тРНК. Участки для тРНК называются Р (пептидильный) и А (акцепторный или аминоацильный).
Ферментом, участвующим в реакции присоединения аминокислоты к тРНК в цитоплазме, является кодаза (аминоацил-тРНК-синтетаза). Процесс узнавания тРНК своей аминокислоты называется рекогниция.
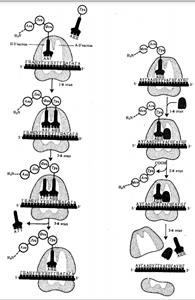
Фаза инициации, или начало синтеза пептида, заключается в объединении двух находящихся до этого порознь в цитоплазме субчастиц рибосомы на определенном участке мРНК и присоединении к ней первой аминоацил-тРНК. Этим задается также рамка считывания информации, заключенной в мРНК (рис. 3.32).
В молекуле любой мРНК вблизи ее 5'-конца имеется участок, комплементарный рРНК малой субчастицы рибосомы и специфически узнаваемый ею. Рядом с ним располагается инициирующий стартовый кодон АУТ, шифрующий аминокислоту метионин. Малая субчастица рибосомы соединяется с мРНК таким образом, что стартовый кодон АУТ располагается в области, соответствующей Пучастку. При этом только инициирующая тРНК, несущая метионин, способна занять место в недостроенном П-участке малой субчастицы и комплементарно соединиться со стартовым кодоном. После описанного события происходит объединение большой и малой субчастиц рибосомы с образованием ее пептидильного и аминоацильного участков. К концу фазы инициации П-участок занят аминоацил-тРНК, связанной с метионином, тогда как в А-участке рибосомы располагается следующий за стартовым кодон.
Описанные процессы инициации трансляции катализируются особыми белками — факторами инициации, которые подвижно связаны с малой субчастицей рибосомы. По завершении фазы инициации и образования комплекса рибосома — мРНК — инициирующая аминоацил-тРНК эти факторы отделяются от рибосомы.
Фаза элонгации, или удлинения пептида, включает в себя все реакции от момента образования первой пептидной связи до присоединения последней аминокислоты. Она представляет собой циклически повторяющиеся события, при которых происходит специфическое узнавание аминоацил-тРНК очередного кодона, находящегося в А-участке, комплементарное взаимодействие между антикодоном и кодоном. Благодаря особенностям трехмерной организации тРНК. (см. разд. 3.4.3.1) при соединении ее антикодона с кодоном мРНК, транспортируемая ею аминокислота располагается в А-участке, поблизости от ранее включенной аминокислоты, находящейся в П-участке. Между двумя аминокислотами образуется пептидная связь, катализуемая особыми белками, входящими в состав рибосомы. В результате предыдущая аминокислота теряет связь со своей тРНК и присоединяется к аминоацил-тРНК, расположенной в А-участке. Находящаяся в этот момент в Пучастке тРНК высвобождается и уходит в цитоплазму (рис. 3.33). Перемещение тРНК, нагруженной пептидной цепочкой, из А-участка в Пучасток сопровождается продвижением рибосомы по мРНК на шаг, соответствующий одному кодону. Теперь следующий кодон приходит в контакт с А участком, где он будет специфически «опознан» соответствующей аминоацил-тРНК, которая разместит здесь свою аминокислоту. Такая последовательность событий повторяется до тех пор, пока в А-участок рибосомы не поступит кодон-терминатор, для которого не существует соответствующей тРНК.
Сборка пептидной цепи осуществляется с достаточно большой скоростью, зависящей от температуры. У бактерий при 37 °С она выражается в добавлении к подипептиду от 12 до 17 аминокислот в 1 с. В эукариотических клетках эта скорость ниже и выражается в добавлении двух аминокислот в 1 с.
Фаза терминации, или завершения синтеза полипептида, связана с узнаванием специфическим рибосомным белком одного из терминирующих кодонов (УАА, УАГ или У ГА), когда тот входит в зону А-участка рибосомы. При этом к последней аминокислоте в пептидной цепи присоединяется вода, и ее карбоксильный конец отделяется от тРНК. В результате завершенная пептидная цепь теряет связь с рибосомой, которая распадается на две субчастицы.

8. Трансляционный аппарат клетки. Кинетическая коррекция трансляции.
· ТРАНСЛЯЦИЯ - процесс синтеза белка из аминокислот по матрице мРНК на рибосомах в цитоплазме. Аминокислоты соединяются в последовательность, определяемую нуклеотидными основаниями мРНК, и доставляются в рибосому транспортной РНК (тРНК). Каждая аминокислота имеет свою собственную тРНК, у которой есть область (антикодон), комплементарная кодону на мРНК.
В рибосоме имеются три различных участка, с которыми связывается РНК: один для мРНК и два – для тРНК. Участки для тРНК называются Р (пептидильный) и А(акцепторный или аминоацильный).
Ферментом, участвующим в реакции присоединения аминокислоты к тРНК в цитоплазме, является кодаза (аминоацил-тРНК-синтетаза). Процесс узнавания тРНК своей аминокислоты называется рекогниция.
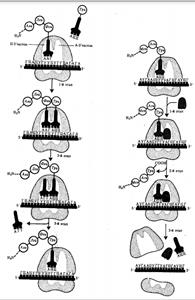
Фаза инициации, или начало синтеза пептида, заключается в объединении двух находящихся до этого порознь в цитоплазме субчастиц рибосомы на определенном участке мРНК и присоединении к ней первой аминоацил-тРНК. Этим задается также рамка считывания информации, заключенной в мРНК (рис. 3.32).
В молекуле любой мРНК вблизи ее 5'-конца имеется участок, комплементарный рРНК малой субчастицы рибосомы и специфически узнаваемый ею. Рядом с ним располагается инициирующий стартовый кодон АУТ, шифрующий аминокислоту метионин. Малая субчастица рибосомы соединяется с мРНК таким образом, что стартовый кодон АУТ располагается в области, соответствующей Пучастку. При этом только инициирующая тРНК, несущая метионин, способна занять место в недостроенном П-участке малой субчастицы и комплементарно соединиться со стартовым кодоном. После описанного события происходит объединение большой и малой субчастиц рибосомы с образованием ее пептидильного и аминоацильного участков.К концу фазы инициации П-участок занят аминоацил-тРНК, связанной с метионином, тогда как в А-участке рибосомы располагается следующий за стартовым кодон.
Описанные процессы инициации трансляции катализируются особыми белками — факторами инициации, которые подвижно связаны с малой субчастицей рибосомы. По завершении фазы инициации и образования комплекса рибосома — мРНК — инициирующая аминоацил-тРНК эти факторы отделяются от рибосомы.
Фаза элонгации, или удлинения пептида, включает в себя все реакции от момента образования первой пептидной связи до присоединения последней аминокислоты. Она представляет собой циклически повторяющиеся события, при которых происходит специфическое узнавание аминоацил-тРНК очередного кодона, находящегося в А-участке, комплементарное взаимодействие между антикодоном и кодоном. Благодаря особенностям трехмерной организации тРНК. (см. разд. 3.4.3.1) при соединении ее антикодона с кодоном мРНК. транспортируемая ею аминокислота располагается в А-участке, поблизости от ранее включенной аминокислоты, находящейся в П-участке. Между двумя аминокислотами образуется пептидная связь, катализуемая особыми белками, входящими в состав рибосомы. В результате предыдущая аминокислота теряет связь со своей тРНК и присоединяется к аминоацил-тРНК, расположенной в А-участке. Находящаяся в этот момент в Пучастке тРНК высвобождается и уходит в цитоплазму (рис. 3.33). Перемещение тРНК, нагруженной пептидной цепочкой, из А-участка в Пучасток сопровождается продвижением рибосомы по мРНК на шаг, соответствующий одному кодону. Теперь следующий кодон приходит в контакт с Аучастком, где он будет специфически «опознан» соответствующей аминоацил-тРНК, которая разместит здесь свою аминокислоту. Такая последовательность событий повторяется до тех пор, пока в А-участок рибосомы не поступит кодон-терминатор, для которого не существует соответствующей тРНК.
Сборка пептидной цепи осуществляется с достаточно большой скоростью, зависящей от температуры. У бактерий при 37 °С она выражается в добавлении к подипептиду от 12 до 17 аминокислот в 1 с. В эукариотических клетках эта скорость ниже и выражается в добавлении двух аминокислот в 1 с.
Фаза терминации, или завершения синтеза полипептида, связана с узнаванием специфическим рибосомным белком одного из терминирующих кодонов (УАА, УАГ или У ГА), когда тот входит в зону А-участка рибосомы. При этом к последней аминокислоте в пептидной цепи присоединяется вода, и ее карбоксильный конец отделяется от тРНК. В результате завершенная пептидная цепь теряет связь с рибосомой, которая распадается на две субчастицы.

· Убиквитин выполняет функцию «метки смерти» для дефектных белков: его присоединение к N-концу белка – сигнал для начала работы протеаз.
Кинетическая коррекция трансляции осуществляется с помощью ферментного комплекса, называемого фактор элонгации.

9. Генетическая инженерия как новый этап развития биотехнологии. Синтез гена. Способы внедрения в клетки искусственно синтезированных генов.
Генная (генетическая) инженерия – совокупность приемов, методов и технологий выделения генов из организмов (клеток), осуществления манипуляций с генами и введения их в другие организмы.
Получение генов.
Для химического синтеза необходимо иметь полностью расшифрованную последовательность нуклеотидов. Последовательность нуклеотидов в ДНК определяют по и-РНК. Химическим путем можно синтезировать небольшие по размеру гены прокариот. Синтез генов эукариот, состоящих из 1000 и более нуклеотидов путем химического синтеза создавать не удается. Наиболее успешным оказался ферментативный синтез. Это метод поколебал центральную догму молекулярной генетики, утверждающую, что считка информации происходит в направлении ДНК→и-РНК→белок. Оказалось, что РНК может быть предшественником ДНК. Подобное наблюдается у онкогенных РНК содержащих вирусов. С РНК вируса, попавшего в клетку, синтезируется ДНК-копия РНК с помощью фермента – обратная транскриптаза. Сам процесс называется обратная транскрипция.
Но гены, синтезированные с помощью ревертаз (обратная транскриптаза) не имеют регуляторной части, а это препятствует функционированию искусственных генов в животных клетках, что ограничивает их использование. Кроме того, и-РНК в клетках очень немного, и она не стойкая. В настоящее время рекомбинантные молекулы ДНК чаще всего получают путем гибридизации инвитро фрагментов ДНК вирусного и бактериального происхождения, и в меньшей степени эукариотического происхождения. Системы доставки экзогенных ДНК:
1. «бомбардировка» частицами на поверхности которых находятся гены
2. электропорация - создание пор в клетке под действием удара, через которые ДНК попадают в клетку.
3. липосом-опосредованный транспорт
4. рецептор-опосредованный транспорт (доставка генов к определенным клеткам)
5. использование векторных молекул
Вектор — молекула ДНК, способная к включению чужеродной ДНК и к автономной репликации, служащая инструментом для введения генетической
информации в клетку. Основные типы векторов:
· аденовирусные (для внедрения в покоящиеся клетки)
· герпесные (в клетки нервной системы)
· ретровирусные ( в быстро размножающиеся клетки)
Дата: 2019-07-31, просмотров: 1027.