Известно, что при воспалении и инфекции происходит изменение уровня активности цитохромов Р450. Этот эффект опосредован цитокинами, которые угнетают транскрипцию генов и накопление мРНК различных изоформ цитохрома Р450 в клетках [Renton, 2004]. Такая реакция взаимодействия иммунной системы и системы метаболизма ксенобиотиков на внедрение инфекционного агента обеспечивает защиту от последствий возможной неконтролируемой активации потенциально опасной для организма ферментативной системы и снижает риск развития оксидативного стресса. В связи с этим представляется важным изучение полиморфизма генов системы метаболизма ксенобиотиков в отношении ТБ инфекции.
Таблица 10
Распределение генотипов исследуемых генов ферментов биотрансформации у больных туберкулезом и здоровых
| Генотип | ТБ | Контрольная группа | р | ||
| n | % | n | % | ||
| GSTT1 | |||||
| GSTT1 + | 254 | 81,9 | 103 | 76,3 | 0,195 |
| GSTT1 0/0 | 56 | 18,1 | 32 | 23,7 | |
| GSTM1 | |||||
| GSTM1 + | 126 | 40,6 | 61 | 45,2 | 0,404 |
| GSTM1 0/0 | 184 | 59,4 | 74 | 54,8 | |
| GSTP1 313A>G | |||||
| AA | 144 | 47,2 | 58 | 48,7 | 0,034 |
| AG | 142 | 46,6 | 45 | 37,8 | |
| GG | 19 | 6,2 | 16 | 13,5 | |
| CYP2E1 7632T>A | |||||
| ТТ | 246 | 80,4 | 106 | 82,8 | 0,235 |
| ТА | 59 | 19,3 | 20 | 15,6 | |
| АА | 1 | 0,3 | 2 | 1,6 | |
| CYP2E1 1293G>C | |||||
| C1C1 | 287 | 92,3 | 113 | 92,6 | 0,498 |
| C1C2 | 24 | 7,7 | 9 | 7,4 | |
| C2C2 | 0 | 0,0 | 0 | 0,0 | |
| CYP2C19 681G>A | |||||
| *1/*1 | 234 | 76,5 | 90 | 77,6 | 0,828 |
| *1/*2 | 69 | 22,5 | 26 | 22,4 | |
| *2/*2 | 3 | 1,0 | 0 | 0,0 | |
Примечание. n – абсолютное значение; р – достигнутый уровень значимости по точному тесту Фишера.
При сравнении распределения частот аллелей и генотипов полиморфных вариантов генов метаболизма ксенобиотиков между группой больных ТБ и контрольной выборкой получены различия только для гена GSTP1, обусловленные увеличением доли гомозигот G/G у здоровых (р=0,034) (табл. 10). Установлена протективная роль генотипа G/G гена GSTP1 в отношении развития ТБ (OR=0,43; 95%CI: 0,20-0,91; р=0,026). В результате однонуклеотидной замены аденина на гуанин в этом гене происходит изменение ферментативной активности, обусловливающее повышение уровня гидрофобных аддуктов в тканях легких. Показано, что соответствующая замена изолейцина на валин в 105 положении, расположенная в субстрат-связывающем Н-участке фермента, приводит к неоднозначным изменениям кинетических параметров ферментам. Ген GSTP1 в большей степени экспрессируется в респираторном тракте. Возможно, что для гомозигот G/G гена GSTP1 характерна высокая каталитическая активность по отношению к ряду соединений, являющихся факторами риска развития ТБ (например, продукты табачного дыма), которая способствует их быстрому метаболизму и дальнейшему выведению, таким образом, снижая риск развития заболевания.
Сравнительная оценка распределения частот аллелей и генотипов генов GSTT1, GSTM1, CYP2E1 и CYP2C19 между группами больных ТБ и здоровых индивидов cтатистически значимых различий не показала (табл. 10).
Проведен анализ полиморфизма исследуемых генов для определения связи с отдельными клиническими проявлениями ТБ, характеризующими тяжесть и течение патологического процесса. Особенно важной является оценка связи исследуемых полиморфизмов с лабораторными показателями, свидетельствующими о степени тяжести туберкулезного процесса и выраженности воспалительной реакции.
Исходя из данных рентгенологического исследования, были сформированы три группы больных ТБ в зависимости от объема поражения легкого (I группа – поражено 1-2 сегмента легкого (n=76), II группа – поражена одна доля легкого (n=40), III группа – задействовано более доли легкого (n=117)). При анализе распределения частот аллелей и генотипов исследуемых локусов получены различия для гена CYP2C19: отмечено преобладание гомозигот *1/*1 в III-й группе по сравнению с I-ой (80,3% и 65,8%, соответственно, р=0,040) и повышенная частота аллеля CYP2C19*1 в III-й группе (р=0,045). Эти данные свидетельствуют в пользу гипотезы о накоплении в клетках активных форм кислорода и реактивных метаболитов, образующихся в реакциях I фазы. Известно, что CYP2C19 семейства цитохрома Р450 является основным ферментом, катализирующим в организме человека трансформацию (S)-мефенитоина в соответствующее 4’-гидроксипроизводное [Крынецкий, 1996]. Замена 681G>A в пятом экзоне гена является основным генетическим дефектом, приводящим к инактивации CYP2C19, проявляющимся на уровне фенотипа наличием медленных метаболизеров (S)-мефенитоина. Полученные различия в отношении развития ТБ можно трактовать следующим образом: у носителей CYP2C19*1 аллеля при метаболизме субстратов для CYP2C19 повышается уровень реактивных окислителей, которые независимо от работы ферментов II фазы, накапливаются в организме и оказывают повреждающее действие на клетки, усиливая патологический процесс. Развитие окислительного стресса при ТБ способствует усилению процессов деструкции в легочной ткани. Подобные результаты были получены при исследовании инсерционного полиморфизма гена CYP2E1 с развитием инфильтративного ТБ у жителей Башкортостана [Бикмаева и др., 2004].
Основной этиологической причиной в развитии туберкулезного процесса являются микобактерии ТБ, несмотря на важное значение факторов, способствующих развитию заболевания (курение, асоциальный образ жизни и др.). С момента открытия Р.Кохом в 1882 году, возбудитель ТБ достаточно полно изучен в отношении биологических свойств, характера и условия заражения. Сложилось четкое представление о патогенезе заболевания, его клинических проявлениях, течении и исходах. ТБ отличается клиническим полиморфизмом, что проявляется в развитии различных форм заболевания – от малых с бессимптомным течением до обширных деструктивных процессов с выраженной клинической картиной. Известно, что контакт организма человека с M. tuberculosis не обязательно приводит к развитию болезни [Хоменко, 1990]. В легких процесс начинается с формирования небольшого очага, где происходит накопление серозного экссудата с нейтрофильными гранулоцитами и макрофагами, подвергающегося первичному некрозу. В дальнейшем экссудативные изменения заканчиваются формированием туберкулезной гранулемы, затем при благоприятном течении процесс ограничивается рубцеванием или инкапсуляцией. Развитие туберкулезных изменений в ранее не инфицированном M. tuberculosis организме свидетельствует о первичном генезе заболевания. В случае обострения существующего первичного процесса, либо повторного экзогенного инфицирования, возникает вторичный ТБ [Цинзерлинг, 1996]. Несмотря на накопленные данные о патогенезе заболевания до сих пор не решен вопрос, почему в одних случаях развивается очаговый ТБ, а в других – инфильтративный или другие клинические формы [Хоменко, 1990].
Предположив, что различия в патогенезе заболевания связаны с индивидуальными особенностями генома человека, был изучен полиморфизм генов системы метаболизма в группах больных ТБ, различающихся по клиническим формам заболевания. Наиболее характерной клинической формой заболевания среди исследуемых больных жителей г. Томска является инфильтративный ТБ (50,3%) (табл. 11). По Сибирскому Федеральному округу также отмечается преобладание вторичного ТБ, среди которых инфильтративная форма занимает первое место в общей структуре клинических форм и составляет 55,1% [Краснов, 2004].
Процентное соотношении доли индивидов мужского и женского пола колеблется в зависимости от клинической формы заболевания. В выборке больных ТБ г. Томска отмечена высокая заболеваемость диссеминированной формой у мужчин по сравнению с женщинами (р=0,047), для остальных форм статистически значимых различий не показано (р=0,602-0,783). По мнению многих исследователей, более высокая заболеваемость у мужчин обусловлена не только биологическими свойствами макроорганизма, но и особенностями условий труда и влиянием различных вредных привычек [Рабухин, 1976; Хоменко, 1990].
Таблица 11
Половозрастной состав групп больных туберкулезом с различными клиническими формами
| Группы сравнения | Мужчины n, (%) | Женщины n, (%) | Cуммарно n, (%) | Средний возраст ± S.D. |
| ТБ внутригрудных лимфоузлов | 21 (9,9) | 13 (12,6) | 34 (10,8) | 6,8 ± 3,4 |
| Очаговый ТБ | 17 (8,1) | 10 (9,7) | 27 (8,6) | 35,7 ± 14,1 |
| Диссеминированный ТБ | 53 (25,1) | 15 (14,6) | 68 (21,7) | 36,9 ± 13,3 |
| Инфильтративный ТБ | 104 (49,3) | 54 (52,4) | 158 (50,3) | 33,4 ± 12,7 |
| Остальные формы | 16 (7,6) | 11 (10,7) | 27 (8,6) | 36,4 ± 11,6 |
Примечание. n – численность в группе; S.D. – стандартное отклонение.
При сравнительном изучении распределения частот «нулевого» генотипа гена GSTM1 группы больных первичным ТБ с поражением внутригрудных лимфоузлов и пациентов с инфильтративной формой отмечена тенденция к снижению доли лиц, гомозиготных по делеции, среди индивидов с ТБ вторичного генеза (70,6% и 52,3%, соответственно, р=0,058).
Анализ частот генотипов полиморфизма 313A>G гена GSTP1 в группах больных с ТБ внутригрудных лимфоузлов и инфильтративным ТБ также показал различия между этими двумя группами (р=0,026) за счет преобладания гетерозигот A/G у лиц больными вторичным ТБ по сравнению с первичной формой (50,0% и 34,4% соответственно).
Таким образом, отмечена связь полиморфных вариантов генов ферментов I и II фаз метаболизма GSTP1 313A>G и CYP2C19 681G>A в развитии ТБ легких у русских жителей г. Томска. Показана протективная роль генотипа G/G полиморфизма гена GSTP1 313A>G для развития ТБ легких. В то же время, установлено возможное модифицирующее влияние полиморфизма 681G>A гена CYP2C19 на увеличение объема зоны поражения легочной ткани при уже возникшем заболевании. Эти данные говорят о том, что изученные полиморфные варианты генов системы метаболизма оказывают вклад на развитие туберкулезного процесса у русских г. Томска.
3.2.3. Сравнительный анализ роли полиморфных вариантов генов системы метаболизма ксенобиотиков в детерминации бронхиальной астмы и туберкулеза
Согласно поставленной задаче, в ходе исследования был проведен сравнительный анализ распределения частот аллелей и генотипов полиморфных вариантов исследуемых локусов между группами больных ТБ и БА. Для делеционного полиморфизма гена GSTM1 показано, что частота «нулевого» генотипа у больных БА превышает таковую в группе больных ТБ (71,0% и 59,4% соответственно; c2=4,85, р=0,028) (рис. 4), а относительный риск развития БА по сравнению с ТБ у носителей «нулевого» генотипа гена GSTM1 составил 1,68 (95% CI: 1,06-2,67). Полученные данные еще раз подтверждают высокую важность функционирования глутатионовой S-трансферазы m1 в развитии БА, поскольку были получены различия в частотах генотипов данного полиморфизма гена GSTM1 при сравнении больных БА и контрольной группы (р=0,008). Для полиморфных вариантов генов глутатионовых S-трансфераз q1 и p1 при сравнении частот генотипов статистически значимых различий не показано (р=0,162 и р=0,387 соответственно).
|
|
|
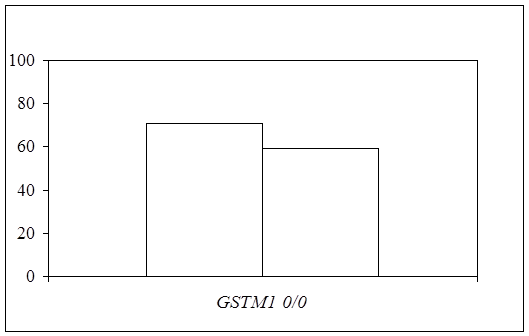
Рис. 4. Частоты «нулевых» (0/0) генотипов гена GSTM1 у больных бронхиальной астмой и туберкулезом.
При сравнении частот генотипов исследуемых в настоящей работе полиморфных вариантов генов цитохромов Р450 различий не было показано между группами больных БА и ТБ (р=0,079-0,437). Среди больных ТБ преобладал *1/*1 генотип полиморфизма 681G>A гена СYP2C19 по сравнению с больными БА (76,5% и 66,4% соответственно), однако эти различия статистически не значимы (р=0,079). В то же время были получены различия между группами больных с ТБ и БА при сравнении частот аллелей этого локуса, где частота СYP2C19*1 аллеля у больных ТБ выше по сравнению с больными БА (87,7% и 82,5% соответственно; c2=3,96, р=0,047). Интересно, что СYP2C19*1 аллель участвует в увеличении объема поражения легочной ткани у больных ТБ легких, и, по-видимому, данная ассоциация предполагает важное участие гена СYP2C19 именно в формировании клинических проявлений при ТБ. Соответственно, для индивидов носителей СYP2C19*2 аллеля среди больных БА относительный риск выше по сравнению с больными ТБ (OR=1,52, 95% CI: 1,01-2,30; p=0,047).
Таким образом, при сравнительном анализе распределения частот аллелей и генотипов полиморфных вариантов генов ферментов метаболизма ксенобиотиков между больными ТБ и БА были получены следующие различия: «нулевой» генотип гена GSTM1 и носительство СYP2C19*2 аллеля полиморфизма 681G>A гена СYP2C19 определяют развитие БА.
Дата: 2019-07-24, просмотров: 361.