Institute of Psychology, USSR Academy of Sciences and Institute of Neurology, USSR Academy of Medical Sciences, Moscow
Summary
The change of conscious mental processes to unconscious ones is usually observed under conditions of monotonousness and occurs periodically. The periodicity is manifested both in the mental activity dynamics and the infraslow oscillations of the brain potentials. In both phenomena three classes of periods areobserved: 2-5 min., 7-12 min., and about 12 min. The periodicity was recorded in five series of experiments: 1) in transitions from the conscious perception of a text to an unconscious switchoff; 2) under reduced informational interaction with the environment; 3) after prolonged mental tension under conditions of relaxed attention; 4) in the course of the evolvement of mental automatism; 5) during activity, with the intuitive aspect predominating. The fluctuations may reflect the operation of mechanisms directed at sustaining the attention in the absence of factors of novelty.
Литература
1. Маркс К., Энгельс Ф., Из ранних произведений, М., 195G.
2. Аладжалова Н. А., Сверхмедленные ритмические изменения электрического потенциала головного мозга. Биофизика, 1956, 1, 2, 127.
3. Аладжалова И. А., Медленные электрические процессы в головном мозге, М., 1962.
4. Аладжалова Н. А., Многоминутная периодичность в биоэлектрических явлениях мозга человека, проходящая сквозь все циклы сна. Тезисы Всесоюзного симпозиума "Саморегуляция процессов сна", Ленинград, 1974, 3.
5. Аладжалова Н. А., Каменецкий С. Л., Сверхмедленные колебания потенциала головного мозга человека при переходе от бодрствования в гипнотическое состояние. Вопросы психологии, 1974, I, 94.
6. Аладжалова Н. А., Кольцова А. В., Каштоянц О. X., Частотный спектр сверхмедленных ритмических колебаний потенциала в мозге человека, Докл. Акад. наук СССР, 1973, 209, 3, 252.
7. Аладжалова Н. А., Кольцова А. В., Каштоянц О. Х., Микаэлян М. X., Сверхмедленные колебания ритмических потенциалов мозга человека во сне. В кн.: Функциональные состояния мозга, М., 1975, 62.
8. Бехтерева Н. П., Бондарчук А. Н., Смирнов В. Н., Трохачев А. И., Физиология и патофизиология глубоких структур мозга человека. Л., 1967.
9. Добрынин Н. Ф., Колебания внимания, М., 1928.
10. Зейгарник Б. В., Патология мышления, М., 1962.
11. Ломов Б. Ф., О системном подходе в психологии. Вопросы психологии, 1975, 2, 31.
12. Лурия А. Р., Мозг человека и психические процессы, М., 1970.
13. Рождественская В. И., Левочкина И. А., Функциональные состояния при монотонной работе и сила нервной системы. В кн.: Проблемы дифференциальной психофизиологии, М., 1972, 5, 11.
14. Смирнов В. М., Сперанский М. М., Медленные биоэлектрические процессы коры и глубоких структур мозга человека и эмоциональное поведение, Вопросы психологии, 1972, 3.
15. Хомская Е. Д., Мозг и активация, М., 1972.
16. Шорохова Е. В., В кн.: Теоретические проблемы психологии личности, М., 1974, 3.
17. Aladjalova, N. A., Slow electrical processes in the brain. Progress in Brain Research, 7, Elsevier, Amsterdam, London, N. Y., 1964.
18. Antrobus, J. S., Information theory and stimulus-independent thought. Brit. J. Psychol., 59, 423, 1968.
19. Filler, M. S., Giambra, L. N., Daydreaming as a function of cueing and task difficulty. Percept, and Mot. Skills, 37, № 2, 503, 1973.
20. Farley, F. H., Measuring the stimulation-seeking motive by global self-rating. Percept. and Mot. Skills, 39, № 1, Part 1, 101, 1974.
21. Kripke, An ultradian biologic rhythm associated with perceptual deprivation and REM sleep. Psychosomatic Med., 34, 3, 221, 1972.
22. Lindsley, D. В., Attention, consciousness, sleep and wakefulness. In: Handbook of Physiology, 1960, v. 3, p. 1553.
23. McKim, R. H., Related attention, J. Creat. Behav., 8, № 4, 265, 1974.
54. О физиологических механизмах "психологической защиты" и безотчетных эмоции. Э. А. Костандов
Центральный НИИ судебной психиатрии, Москва
При тахиотоскопических экспозициях отдельных слов Мак-Джини [38] впервые обнаружил, что у нормальных исследуемых - студентов - порог "осознаваемого" опознания ругательных слов ("неприличные" или "табу" - слова) явно повышен по сравнению с порогом "нейтральных" слов. На "табу"-слова регистрировалась кожно-гальваническая реакция (КГР) во время таких коротких экспозиций, при которых исследуемые еще не могли прочесть слово. Догадки об этих "табу"-сло-вах, если они при этом делались, обычно не были связаны с содержанием слова. Было удивительно наблюдать, отмечает Мак-Джини, как нормальная молодая девушка с ненарушенным зрением в отдельных случаях была не в состоянии прочесть "неприятные" слова, в то время как они были ясно видны другому наблюдателю. Факт повышения порога опознания слова Мак-Джини объяснял как "психологическую защиту", как результат существования как бы фильтра в зрительном восприятии, который, насколько это возможно, предохраняет субъекта от осознания эмоционально неприятных раздражителей внешней среды. Интересно, что уже в этой первой работе Мак-Джини поставил вопрос о нейрофизиологическом объяснении обнаруженного им феномена. Он допускал два возможных нервных .механизма развития КГР еще до того, как "табу"-слово осознается исследуемым: КГР - результат действия обратной связи из корковых ассоциативных центров или же прямого действия зрительных импульсов на таламус.
Описанный Мак-Джини факт пороговых изменений восприятия эмоционально значимых стимулов и их влияния на различные функции человека на подпороговом "бессознательном" уровне был подтвержден в многочисленных работах [14; 26; 48; 49; 50; 51]. Вместе с тем, все эти работы подверглись серьезной критике с методической стороны [20; 21; 25; 29; 30]. Было высказано мнение о том, что пороги опознания слов, измеряемые восходящим методо.м границ, сильно зависят от частоты их употребления в прошлом данным субъектом. Кроме того, человек может сознательно тормозить свою словесную реакцию, т. е. воздерживаться от произнесения "табу"-слов. Однако в специально поставленных опытах было показано, что частота применения слов в прошлом не оказывает существенного влияния на порог их опознания [32; 41].
Лазарус и Мак-Клири [33] получили экспериментальным путем феномен повышения порога опознания эмоциональных стимулов. У здоровых взрослых людей они предварительно вырабатывали на отдельные слоти оборонительную условную реакцию на электрокожном подкреплении. После этого измерялись пороги зрительного опознания различных слогов. Оказалось, что пороги опознания слогов, на которые была выработана оборонительная реакция, явно выше, чем на другие "нейтральные" слоги, хотя какой-либо разницы в степени употребления исследуемыми этих слогов в прошлом не было и нельзя было думать, что они подавляют свою реакцию. Фарес [44] повторил эти опыты и обнаружил, что пороги опознания слогов, наоборот, понижаются, если до этого на них вырабатывается оборонительная реакция избегания. Диксон и Лир [19] отмечали как повышение, так и понижение; Гольдштейн и Хим-мельфарб [27] - только повышение порога такистоскопически предъявляемых эмоциональных слов.
Диксон и Лир [19] повышение порога опознания эмоциональных слов пытаются объяснить предеознательной (prior to awareness) оценкой предъявляемых стимулов, в результате чего изменяется уровень корковой активности. В эксперименте, проверяющем эту гипотезу, ее авторы регистрировали фоновую ЭЭГ во время процедуры определения порога зрительного опознания слов. У одной части исследуемых наблюдался высокий порог опознания эмоциональных слов по сравнению с нейтральными словами, у другой - более низкий. Оказалось, что амплитуда альфа-активности, регистрируемой до опознания эмоционального вербального стимула, положительно коррелирует с величиной порога опознания: при высоком пороге отмечается синхронизация ЭЭГ в затылочной области, при низком - уменьшение синхронности. Эти данные, по мнению Диксона и Лира, подтверждают их гипотезу о нервном механизме пороговых изменений восприятия эмоциональных слов, как результат предеознательного изменения уровня корковой активности, осуществляемой ретикулярной формацией ствола мозга.
К сожалению, кроме упомянутой работы Диксона и Лира [19], нам не удалось обнаружить исследований, в которык с физиологических позиций изучались бы нервные механизмы "психологической защиты" и реакции человека на неосознаваемые стимулы. Эти два вопроса тесно связаны между собой, так как подпороговый эффект неосознаваемых эмоциональных слов проявляется только в случаях повышения порогов их осознания. Как справедливо пишет Спенс [50], восприятие "без осознания" и "психологическая защита", т. е. повышение порога опознания внешнего стимула, - это две стороны одной медали.
Принципиальная связь подпорогового эффекта эмоциональных слов и явления "психологической защиты" обнаружилась также и в наших исследованиях. Они проводились на возбудимых психопатических личностях, которые находились в тяжелой конфликтной жизненной ситуации. Эти лица с эмоционально-лабильным характером, пребывающие длительное время в состоянии эмоционального стресса, являются весьма подходящим объектом для исследования особенностей восприятия эмоционально-значимых раздражителей. В этих случаях значительно облегчается задача подбора словесных раздражителей, вызывающих отрицательные эмоции.
После десятиминутной адаптации к темноте проводилось измерение порога опознания "нейтральных" (общеупотребительные имена существительные) и "конфликтных" или "эмоциональных" слов, имеющих отношение к трудной жизненной ситуации исследуемого. Отдельное слово предъявлялось на электролюминесцентном экране с малым послесвечением (1 мсек). Время экспозиции слова в начале его предъявления было заведомо небольшим (50 мсек), так что исследуемый не мог его прочесть. Затем периоды экспозиции слова постепенно (с паузой между каждым предъявлением в 20-60 секунд) увеличивались (по 20-50 мсек) до тех пор, пока исслздуемый, согласно предварительной инструкции, не произносил вслух это слово без ошибок. Таким образом, порог восприятия слова определялся по минимальному времени экспозиции, которое было необходимо исследуемому для его "сознательного" опознания. После двух-четырех нейтральных слов предъявлялось эмоциональное слово, затем снова одно-два нейтральных. Сопоставлялись пороги опознания слов с одинаковым количеством букв.
С целью выявления подпорогового действия эмоционального слова в течение нсей процедуры измерения порога как нейтральных, так и эмоциональнььх слов на полиграфе регистрировались фоновая ЭЭГ, КГР (по Тарханову), дыхательные движения грудной клетки, частота пульса и плетизмограмма пальца. Подпороговый эффект определялся по разнице в порогах между словесным отчетом об опознании слова и биоэлектрической или вегетативной реакцией.
Порог опознания нейтральных слов в наших исследованиях колеблется в довольно широких пределах - от 100 до 600 мсек (таблица). Он в определенной степени зависит от числа букв в слове и, по-видимому, от типологических особенностей исследуемого, его прошлого жизненного опыта, установки на исследование и т. д.

Разница порогового осознания нейтральных и эмоциональных слов
У всех исследуемых пороги опознания эмоциональных слов, имеющих отношение к их тяжелой жизненной ситуации, значительно отличаются от порогов опознания нейтральных слов. Наблюдается как понижение, так и повышение порогов опознания эмоциональных слов. Последнее происходит чаще, приблизительно в двух третях случаев. Как это видно из таблицы, разница между нейтральными и эмоциональными словами может быть весьма существенной, более чем в два раза. Повышение порога опознания эмоциональных слов, по-видимому, находится в прямой зависимости от степени аффектогенности слова. У одного и того же человека порог опознания разных эмоциональных слов может значительно колебаться. Так у исследуемого Ш. В., убившего по мотивам ревности свою жену, порог опознания слова "жена" значительно выше другого эмоционального слова - "арест" (таблица). У исследуемой С. Т. (психопатическая личность, совершила кражу и поджог, очень боится своего отца) порог опознания слов, имеющих отношение к краже, значительно выше порога слова "поджог".
Интересный и сложный вопрос о том, почему в одних случаях происходит "сенсибилизация" восприятия, а в других, наоборот, повышение порога опознания эмоционально значимых стимулов, т. е. "психологическая защита", требует специального анализа и обсуждения. В данной работе будут рассмотрены физиологические механизмы явления "психологической защиты", так как эффект неосознаваемых эмоциональных слов проявляется только в случаях повышения порогов их опознания.
Уже в первых работах, обнаруживших повышение порога опознания эмоциональных слов, регистрировались кожно-гальванические реакции пои столь коротких экспозициях, время которых не давало возможности исследуемому их прочесть [38]. Вскоре же появился ряд работ, в которых хотя и не отрицался по существу факт развития КГР при экспозиции неосознаваемого слова, но этому феномену давалась другая трактовка. Так Брюнер [29] предположил, что КГР,развивающаяся при предпороговых экспозициях "табу"-слов, отражает большие усилия субъекта, которые он прилагает, чтобы увидеть более "трудное" слово, т. е. слово с более высоким порогом опознания.
Было также высказано предположение, что "подпороговый эффект" слова в виде КГР может быть следствием того, что процедура определения порога опознания требует от субъекта определенного словесного ответа типа "да-нет". В подобной ситуации исследуемый, особенно если он принадлежит к типу "консерваторов", может не сообщить о словесном раздражителе, в вербальной оценке которого он сомневается, хотя слово, по крайней мере частично, им осознается. При этом нередко возникают биоэлектрические и вегетативные реакции, которые могут ошибочно относиться на счет "надпорогового восприятия" [25; 29]. Для проверки этого "методического" объяснения феномена "психологической защиты" были проведены опыты, в которых исследуемые по предварительной договоренности во время процедуры измерения порогов сообщали о всех своих догадках, о словах, которые они еще не могли ясно опознать [38]. Оказалось, что догадки или гипотезы исследуемых о "нейтральных" словах были значительно больше структурно похожи на экспонируемый стимул, чем в случаях предъявлений "табу"-слов. Против представления о КГР на неосознаваемые слова, как о методическом артефакте, говорят также данные [33], о которых мы уже упоминали. В наших экспериментах предварительно угашались но возможности все компоненты ориентировочного рефлекса на нейтральные словесные раздражители. Только после этого начиналась процедура определения порога опознания нейтральных (не тех, которые применялись в предварительных опытах) и эмоциогенных слов. В случаях, когда порог опознания последних выше, чем нейтральных, наблюдается подпороговый эффект: биоэлектрические и вегетативные (реакции возникают раньше, чем исследуемый опознает эмоциональное слово.
Подпороговая зона, т. е. разница между порогом опознания эмоционального слова и порогом биоэлектрических и вегетативных реакций на него, у разных исследуемых и даже у одного и того же лица, но на разные эмоциональные слова колеблется в довольно широких пределах - 90-820 мсек. На рис. 1 приводятся данные исследования С. Т., у которой наблюдалась большая подлороговая зона. При экспозиции в течение 180 мсек нейтрального слова "восток" реакций не отмечается (А). При той же экспозиции слова "воровка" регистрируется четкая КГР (Б). Еще больший подпороговый эффект с резко выраженной КГР вызывает это слово при экспозиции 200 мсек (В). Однако при дальнейшем увеличении экспозиции слова "воровка" реакция не усиливается, а, напротив, резко уменьшается (Г). Даже при экспозиции 800 мсек, близкой к порогу опознания слова, не регистрируется четкой реакции (Д). При опознании слова "воровка" (порог был равен 900 мсек) наступила сильная эмоциональная реакция с плачем, с хорошо выраженными КГР и дыхательной реакцией (Е).
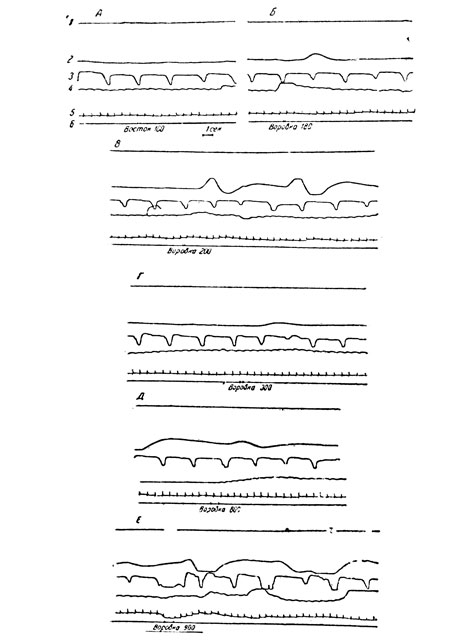
Рис. 1. Исследуемая С. Т. Подпороговый эффект на 'эмоциональное' слово. 1 - электромиограмма руки, 2 - КГР, 3 - дыхание, 4 - плетизмограмма, 5 - ЭКГ. 6 - отметка раздражения и времени его экспозиции
И у других исследуемых подпороговый эффект чаще наблюдается при относительно коротких экспозициях эмоциогенного слова, соответствующих нижней половине подпороговой зоны. С увеличением времени предъявления этот эффект исчезает, и только при экспозиции, близкой к порогу осознания, возникают биоэлектрические и вегетативные реакции, которые, по-видимому, являются уже результатом частично осознанного различения раздражителя. Смит с соавт. [491, Гольдштейн и Бартол [26] также наблюдали более выраженный эффект действия неосознанного словесного раздражителя при его более коротких экспозициях.
Наличие неэффективной зоны, когда влияние слов не усиливается с увеличением их экспозиции в пределах подпороговой зоны, а даже, наоборот, нередко пропадает, является очевидным доказательством того, что повышение порога опознания эмоциональных слов не есть следствие произвольной задержки исследуемым своей речевой реакции на неприятные слова, а является результатом изменений восприятия эмоциональных стимулов.
Факты развития биоэлектрических и вегетативных реакций на эмоциональные слова при экспозициях значительно меньших, чем пороги их опознания, наличие при этом "неэффективной зоны" убеждают нас в том, что центральная нервная система человека в состоянии различать некоторые слова без их осознания. Это различение слов на основе лишь эмоциональной памяти, без оживления в сознании словесных символов происходит, в основном, в случаях, когда порог восприятия эмоционального слова повышен.
Выяснение нервного механизма, который на основании информации, не достигающей уровня сознания, способен оценить эмоциональное значение слова, непосредственно связано с проблемой нейрофизиологического изучения эмоциональной памяти у человека, когда эмоциональное состояние в какой-то степени воспроизводится без отображения эмоциогенных раздражителей в образах или словесных символах, без участия специфически человеческой формы памяти - словесно-логической. И. С. Бериташвили [4; 6] относит к особой форме памяти воспроизведение определенного эмоционального состояния при воздействии раздражителей, связанных с данной эмоцией. В нормальных условиях у высших позвоночных животных это воспроизведение эмоционального состояния осуществляется и регулируется импульсами возбуждения из сенсорных элементов неокортекса, воспринимающих и перерабатывающих информацию из внешнего мира. Но сама эмоциональная память обеспечивается соответствующими структурными изменениями в клетках архипалеокортекса, составляющих интегративный нервный механизм эмоционального поведения [4].
У людей при определенных состояниях или обстоятельствах могут возникать так называемые безотчетные эмоции, без осознания их причин или повода, когда человек не в состоянии понять, почему у него изменилось настроение. Эмоции, психическая сущность которых исчерпывается неясными переживаниями приятного или неприятного, М, И. Аствацатуров 11] называл таламическими или агностическими. Эти эмоции из-за их безотчетности, видимой беспричинности, нередко называли эндогенными. Можно думать, что в этих случаях эмоциональные реакции возникают на основе условнорефлекторной эмоциональной памяти без участия специфически человеческой словесно-логической памяти.
Каким образом это происходит? Как высшие отделы головного мозга "узнают" слово еще до того, как оно опознается, и каковы нервные механизмы пороговых изменений эмоциональных слов? Что это за сверхчувствительный механизм, который на основании информации, не достигающей уровня сознания, способен оценить эмоциональное (смысловое) значение слова и затем повысить или понизить порог его восприятия? Что это за "цензор" сидит позади человеческого глаза, который решает, пропустить ли быстро, с облегчением информацию о данном эмоционально неприятном для человека раздражителе до уровня сознания или же, наоборот, задержать, притормозить и тем самым "защитить" его. На эти вопросы необходимо найти ответы, так как они непосредственно связаны с проблемой изучения физиологических механизмов явлений бессознательного.
Очевидно следует допустить существование в мозгу человека чувствительного механизма, реагирующего на физически очень слабые, но психологически для данной личности весьма значимые раздражители. Функционально-структурная организация этого механизма не обеспечивает осознание эмоционального раздражителя, но его активация может приводить к возникновению биоэлектрических и вегетативных реакций, а также к изменению некоторых психологических функций и состояний. Успехи последних лет в нейрофизиологическом анализе эмоций, выяснение роли лимбическои системы в образовании условных реакций с эмоциональной окраской дают достаточно оснований для того, чтобы предложить гипотезу об упомянутом нервном механизме бессознательных психических явлений.
Клинические наблюдения у людей и эксперименты на животных дали основание считать, что структуры лимбическои системы головного мозга и, в частности, архипалеокортекс являются нервным субстратом эмоциональных переживаний и интеграции эмоциональных выражений [43; 5; 24; 17]. При этом было установлено, что структуры лимбическои системы могут возбуждаться как нисходящими импульсами из неокортекса, так и при периферических раздражениях различных рецепторов через ретикулярную формацию непосредственно через ответвления от специфических сенсорных путей.
Для понимания физиологических механизмов эмоционального поведения человека весьма важен факт тесного взаимодействия новой коры с лимбическои системой. При электрическом раздражении отдельных участков неокортекса можно вызвать различные эмоциональные реакции. Так, при раздражении задних и нижних концов сильвиевых извилин у кошек развивалась реакция "страха" [22]. Такая же эмоциональная реакция наблюдалась у больных эпилепсией при электрической стимуляции височной области [45]. По мнению И. С. Бериташвили [4], нервные импульсы из новой коры непосредственно проводятся к структурам лимбическои системы как по врожденным путям, так и через временные связи, образованные в прошлом. Был поставлен интересный опыт. Электрическое раздражение зрительной области коры, которое само по себе не дает какой-либо внешней реакции, сочеталось с раздражением палеокортекса (поясной извилины), вызывающим эмоциональную реакцию страха. После ряда сочетаний уже одно электрическое раздражение зрительной коры начинало вызывать реакцию страха у животного [13]. Этот факт подтверждает точку зрения о том, что в результате сочетания возбуждений интегративного механизма эмоций в лимбическои системе и воспринимающей области неокортекса образуются временные связи между этими нервными образованиями.
Можно думать, что в случаях длительных и сильных переживаний отрицательных эмоций, как это наблюдается у наших исследуемых, находящихся продолжительное время в тяжелой конфликтной жизненной ситуации, наибольшие пластические изменения происходят в нейронных кругах, связанных с эмоциональным поведением. Вследствие этого резко повышается возбудимость временных связей между сенсорными элементами неокортекса, воспринимающими сигнальные эмоциональные раздражители, и структурами лимбической системы, в которых интегрируются нервные механизмы отрицательной эмоции. Это приводит к тому, что даже при очень небольшой афферентной импульсацин от воздействия физически слабого эмоционального раздражителя, как это было, например, в наших экспериментах с очень короткой экспозицией зрительных словесных стимулов, происходит возбуждение упомянутых временных связей и развитие соответствующей эмоции с комплексом вегетативных и биоэлектрических реакций. При этом раздражитель может не опознаваться субъектом-
Согласно нашей гипотезе, для процесса осознания словесного сигнала решающее значение имеет активация временных связей сенсорных элементов неокортекса с моторной речевой областью и возбуждение последней. Это предположение, как нам кажется, находит фактическое подтверждение в психологических исследованиях людей, подвергшихся операции "расщепление мозга" [52]. Если какое-либо слово, например "ключ", "вилка", "кольцо", предъявляется в левом поле зрения, т. е. зрительная афферентная импульеация поступает только в правое полушарие, а у лиц с "расщепленным мозгом" связь между полушариями, как известно, отсутствует, то исследуемый не может вербализовать, осознать это слово, но он в состоянии согласно предварительной инструкции ощупью отобрать соответствующий предмет. На вопрос, что именно исследуемый выбрал левой рукой, он мог дать неправильный ответ, хотя выбор был сделан правильно.
iB настоящее время имеется достаточно подобного рода данных, которые показывают, что в "немом" правом полушарии имеются гностические зоны, воспринимающие зрительную и слуховую речь. Невозможность вербализации, надо думать, не обусловлена неспособностью правого полушария воспринимать и анализировать, т. е. "понимать речь", в известных, весьма ограниченных пределах, конечно. Для вербализации и тем самым для осознания стимула (простого или словесного) существенно, по-видимому, необходимо поступление информации о нем в левое полушарие, которое четко латерализовано относительно моторной речи.
Таким образом создается представление, по которому отсутствие в правом полушарии представительства моторной речи определяет невозможность вербализации и осознания сигнала. В гностической речевой зоне обоих полушарий осуществляется высший анализ и синтез словесных сигналов, но они не будут осознаваться, если нервные импульсы из этой зоны не поступят в моторную речевую область. Можно думать, что активация двигательной речевой области является решающим звеном в структурно-функциональной организации головного мозга, обеспечивающей осознание раздражителя.
Тесная и необходимая связь сознания и, в частности осознания "ближайшей чувственно воспринимаемой среды", с языком неоднократно подчеркивалась основоположниками диалектического материализма: "Язык так же древен, как и сознание: язык есть практическое, существующее и для других людей и лишь тем самым существующее также и для меня самого, действительное сознание..." [11].
Все вышесказанное о связи сознания с функционированием специализированного механизма моторной речи и о роли лимбической системы в осуществлении эмоциональных реакций позволяет представить себе физиологический механизм действия на человека неосознаваемых им эмоциональных стимулов, в частности словесных. Очевидно, этот "бессознательный" эффект эмоциональных слов обусловлен разностью в порогах активации временных связей, составляющих функциональную систему, которая отражает неприятную или угрожающую ситуацию. Как уже упоминалось, порог активации структур лимбической системы в случаях длительных или сильных эмоциональных переживаний может значительно и на длительное время понижаться. В этих случаях словесные раздражители, сигнализирующие о конфликтной ситуации, при определенных условиях могут вызвать возбуждение временных связей между воспринимающей речевой зоной неокортекса и структурами лимбической системы без активации связей этой корковой зоны с моторной речевой областью.
Возбуждение лимбических структур без активации моторной речевой области приводит к тому, что возбуждаются гипоталамические и стволовые механизмы эмоциональных реакций без осознания раздражителя, вызвавшего эти реакции. Согласно нашей гипотезе, таков физиологический механизм безотчетных эмоций, т. е. реакций человека, воспроизводимых на основе лишь условнорефлекторной эмоциональной памяти, без участия специфически человеческой словесно-логической памяти.
Лимбическая система при ее непосредственном раздражении электрическим током или активации условным раздражителем, з свою очередь, действует на неокортекс, понижая или повышая его возбудимость. Тормозное или облегчающее влияние лимбилеской системы на новую кору было показано в целом ряде экспериментов на животных [2; 18; 37; 39]. Эти восходящие влияния могут изменять - улучшать или ухудшать - восприятие внешних раздражителей. В большинстве случаев, очевидно, это - тормозное влияние, поэтому наблюдается повышение порога опознания.
С целью экспериментального подтверждения нашей гипотезы о том, что эмоциональный словесный раздражитель при определенных условиях может вызывать по механизму временной связи возбуждение лимбической системы, что, в свою очередь, оказывает влияние на восприятие этого раздражителя, нами, совместно с Ю. Л. Арзумановым, были проведены исследования особенностей вызванных потенциалов коры головного мозга на осознаваемые и неосознаваемые эмоциональные слова. Выяснение связи субъективных аспектов восприятия с биоэлектрическими феноменами, в частности с теми или иными компонентами вызванной электрической активности коры головного мозга, представляет важную проблему современной нейро- и психофизиологии. Отражается ли процесс осознания внешнего раздражителя в определенных компонентах вызванных потенциалов коры или же последние являются лишь биоэлектрическим эпифеноменом по отношению к процессу восприятия? По этому вопросу нет единой точки зрения. С одной стороны, можно привести довольно пессимистическое высказывание А. И. Ройтбака [12], считающего, что при расположении электродов на поверхности коры отводится электрическая активность корковых пирамидных нейронов. А так как, по И. С. Беритову [3], психическая деятельность коры (возникновение ощущений, образов) обусловлена, в первую очередь, возбуждением звездчатых нейронов, на которых оканчиваются специфические афференты, то, следовательно, по мнению Ройтбака, мы не регистрируем самого интересного.
В пользу такого мнения говорят факты, полученные в опытах, в которых путем гипнотического внушения изменялось субъективное восприятие стимула. Так, например, здоровым людям в гипнозе в одних случаях внушалось, что световой стимул тусклый, в других - яркий, хотя интенсивность стимула объективно оставалась без изменений. Величина вызванного потенциала коррелировала с физической интенсивностью стимула и не менялась в зависимости от внушенного субъективного восприятия [15]. При вызванной в гипнозе кожной анестезии не было отмечено изменений величины или формы вызванных потенциалов коры на электрокожное раздражение [28]. На фоне внушенной глухоты с вертекса продолжали регистрировать такие же вызванные потенциалы на звук, как и до гипноза [28]. Сходные результаты были получены у больных с истерической гемианастезией [34; 16].
При помощи специальных приемов (подача конкурирующего изображения на второй глаз или стабилизация зрительного образа на сетчатке) у бодрствующего здорового человека добивались исчезновения субъективного восприятия зрительного стимула [46; 47]. В этих случаях со зрительной области коры регистрировались вызванные электрические ответы, которые не отличались по величине и форме от реакций на эти же стимулы, но осознаваемые исследуемым.
Все вышеприведенные факты дают основание думать, что осознание внешнего раздражителя и электрические потенциалы коры головного мозга, вызываемые этим раздражителем, отражают нервные процессы, которые связаны с деятельностью разных функциональных уровней.
С другой стороны, делаются попытки доказать, что в форме вызванных потенциалов коры головного мозга человека можно обнаружить признаки кодирования специфической сенсорной информации [23; 31]. Либэ [35; 36] у бодрствующего взрослого человека субдурально с соматосенсорной коры регистрировал вызванный электрический ответ на слабые раздражения кожи, которые не ощущались исследуемым. Этот потенциал по своей конфигурации отличается от коркового ответа на ощущаемые раздражения: начальный позитивно-негативный комплекс регистрируется, но явно меньшей амплитуды, а поздние положительные и отрицательные волны отсутствуют. Совпадение появления поздних компонентов с возникновением ощущения дает основание Либэ считать, что осознание внешнего стимула связано с развитием поздне-латентных корковых потенциалов. Таким образом, по данным Либэ, осознание внешнего раздражения обеспечивается соответствующей пространственно-временной конфигурацией нейрональной активности в коре, которую можно регистрировать в виде определенной формы вызванного потенциала. Критическим фактором для осознания, таким образом, по Либэ, является развитие поздних, "длиннолатентных" компонентов вызванного потенциала коры.
Эта довольно стройная схема Либэ связывает факт осознания стимула с подключением в корковые процессы влияний неспецифической ретикулярной формации, так как поздние компоненты вызванной активности коры развиваются при ее непременном участии. Она как будто соответствует и данным, наблюдаемым на животных, у которых поздние "вторичные" потенциалы регистрируются при бодрствовании животного и исчезают с развитием наркотического или сонного состояний. Со времен работы Мэгуна и Моруцци [40] стало общепринятым положение о том, что для восприятия внешнего раздражителя необходимо поступление в кору импульеации по обеим афферентным системам (специфической и неспецифической) и конвергенция их на соответствующих воспринимающих элементах. Таким образом, точка зрения Либэ хорошо соответствует существующим нейрофизиологическим концепциям о механизмах бодрствования.
Мы получили, однако, помимо фактов, о которых уже было сказано выше, также многочисленные другие данные, опровергающие представление о том, что критическим фактором для осознания раздражителя являются длиннолатентные (до 500 миллисекунд) нейрональные процессы в коре, отражающиеся в виде поздних вызванных потенциалов.
У взрослого здорового человека на слабый звуковой сигнал, неосознаваемый исследуемым, со скальпа удается отводить усредненный вызванный потенциал в виде позднего низкоамплитудного положительного колебания. Большой скрытый период этой волны (свыше 400 миллисекунд) и тот факт, что она отводится с вертекса, указывают на ее связь с поступлением в кору неспецифической информации. Таким образом, слабый неосознаваемый звук может вызвать восходящую неспецифическую импульеацию, что отражается в коре в виде регистрируемой в наших опытах поздней положительной волны.
В экспериментах с регистрацией усредненной вызванной электрической активности коры на словесные стимулы применялась та же методика предъявления слов на экране, что и в вышеописанных исследованиях порога опознания. Исследуемыми также являлись психопатические личности, находящиеся в тяжелой конфликтной жизненной ситуации. В серии с неосознаваемыми нейтральными и эмоциональными словами экспозиция всех слов на экране равнялась 15 мсек, а освещенность 0,05 лк±20%. Все исследуемые в этой серии во всех пробах не осознавали словесный раздражитель и воспринимали его как тусклую вспышку света на экране, хотя из словесного отчета можно было видеть, что они, как правило, очень старались распознать слово.
После регистрации вызванной электрической активности коры на неосознаваемые нейтральные и эмоциональные слова во второй серии экспериментов производилась запись вызванных потенциалов на те же слова, но уже предъявляемые с другими параметрами (экопозиция 200 мсек, освещенность 0,1 лк±20%), которые давали возможность исследуемому легко прочесть слово. Таким образом мы могли сравнивать усредненные вызванные потенциалы на одни и те же слова, но в одних случаях неосознаваемые исследуемым, в других - осознаваемые. Примененная нами методика регистрации усредненных вызванных потенциалов уже описана подробно [10].
Анализ величины вызванного потенциала показывает, что различие амплитуд поздних положительных волн (компонент Р 300) в затылочной области, возникающих при неосознаваемых нейтральных и неосознаваемых эмоциогенных словах, столь же высоко достоверно (Р<0,001), как при осознаваемых. Как можно видеть на рис. 2, в том и другом случаях амплитуда этой волны на эмоциональные стимулы явно больше.
На рис. 2 можно также видеть и существенное различие, которое выражается в степени распространенности "эмоционального" облегчения вызванного коркового электрического ответа. На осознаваемое эмоциональное слово наблюдается увеличение амплитуды компонента Р 300 только в затылочной области, что дало нам основание говорить об относительно локальном характере дополнительной активации, связанной с эмоциогенным свойством стимула [9]. На неосознаваемые эмоциональные слова не только с затылочной области, но и с вертекса отводится поздняя положительная волна достоверно большей величины, чем на нейтральные слова (Р<0,05).
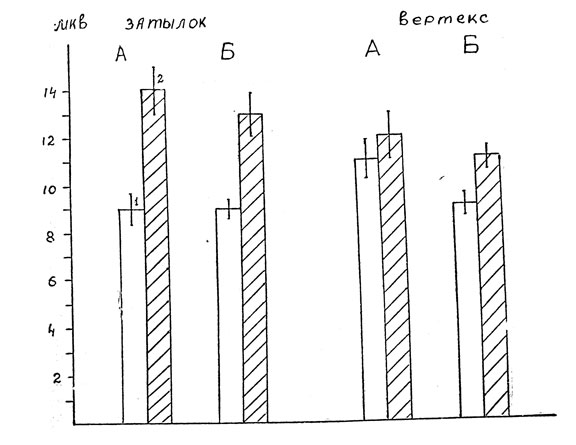
Рис. 2. Изменения амплитуды компонента Р 300 на осознаваемые и неосознаваемые эмоциональные слова. А - осознаваемые, Б - неосознаваемые, 1 - нейтральные стимулы; 2 - эмоциональные
Приведем иллюстрации этого, на наш взгляд, интересного факта. На рис. 3 можно видеть, что у исследуемого Б. А. (психопатическая личность, на почве ревности убил свою жену) на неосознаваемое нейтральное слово "кастрюля" регистрируется позднее положительное колебание (компонент Р300), амплитуда которого в области затылка равняется 10,1 мкв, а в вертексе-10,8 мкв (кадр А). На эмоциональное слозо "расправа", которое также не осознается исследуемым, амплитуда этого компонента электрического ответа коры значительно больше в обоих отведениях: в затылочном - 15,6 мкв; в вертексе - 16,3 мкв (кадр Б).
У этого же исследуемого Б. А. в другой опытный день на нейтральное неосознаваемое слово "трава" амплитуда компонента Р 300 в затылочной области составляет 7,1 мив, а вертексе - 6,9 мкв (рис. 4, А). Эмоциональное же слово "обида" вызывает положительные колебания значительно большей амплитуды: в затылочной области 11,8 мкв, в вертексе - 11,4 мкв (рис. 3, Б).
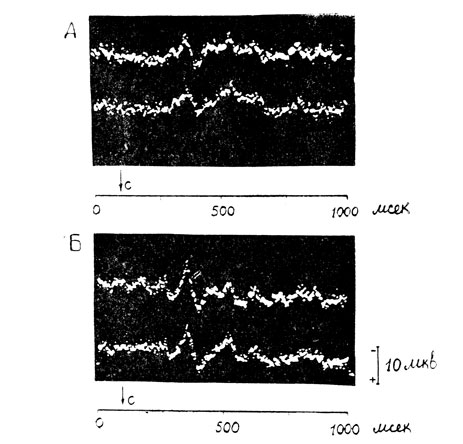
Рис. 3. Увеличение амплитуды позднего вызванного ответа на неосознаваемое эмоциональное слово. Исследуемый Б. А. А - потенциал на нейтральное слово 'кастрюля'; Б - на эмоциональное слово 'расправа'. Стрелка и буква С - момент раздражения. Остальные обозначения: верхняя кривая - вертекс, нижняя - затылочная область. Отклонение луча вверх - негативность. Момент раздражения совпадает с началом кривой. Эпоха анализа - вся кривая - 1000 мсек
Следовательно, в случаях, когда слово, связанное с отрицательным эмоциональным переживанием, не осоэнается человеком, оно вызывает в каре более диффузную активацию, чем при действии этого же раздражителя, но осознаваемого им. Таким образом, одной из отличительных черт вызванной электрической активности коры, связанной с осознанием или неосознанием словесного стимула, является степень распространенности по коре облегчения поздней положительной волны.
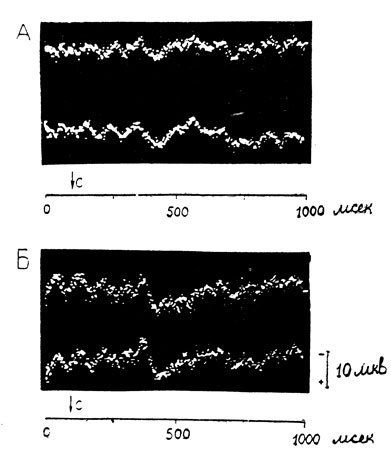
Рис. 4. Увеличение вызванного электрического ответа коры в вертексе и затылочной области на неосознаваемое эмоциональное слово. Исследуемый Б. А. А - на нейтральное слово 'трава'; Б - эмоциональное слово 'обида'. Остальные обозначения те же, что на рис. 3
Другая особенность вызванных потенциалов на неосознаваемые эмоциональные слова связана с поздним отрицательным компонентом N200. Этот потенциал существенно не различается ни по форме, ни по величине при осознаваемых нейтральных и эмоциональных словах. В то же время в пробах, где эти же слова не осознавались исследуемым, отрицательный компонент N 200 на эмоциональные стимулы был явно большей амплитуды как в затылочной области, так и в вертексе (Р<0,01). Усредненные результаты измерения этой волны на осознаваемые и неосознаваемые нейтральные и эмоциональные слова приведены на рис. 5. На графиках ясно видна разница в величине потенциала N 200 между нейтральными и эмоциональными словами в зависимости от того, осознает или не осознает их исследуемый. Увеличение компонента N200 на неосознаваемые эмоциональные слова хорошо видно также на рис. 3 и 4.
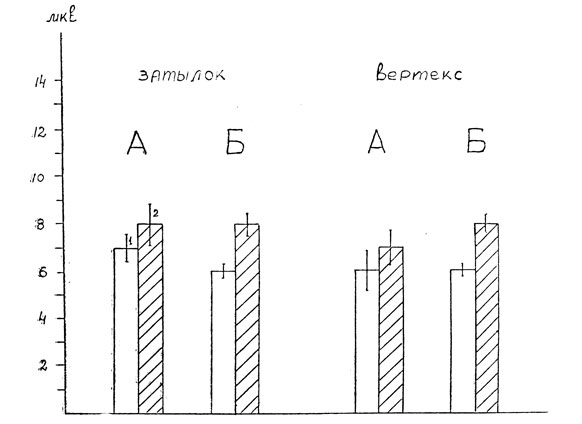
Рис. 5. Изменения амплитуды позднего отрицательного потенциала 200 на неосознаваемые эмоциональные слова. Обозначения те же, что на рис. 2
Вся эта разница между осознаваемыми и неосознаваемыми словами прежде всего подтверждает наше представление о кортикофугальном механизме вовлечения влияний лимбической системы в корковые реакции на эмоциональные стимулы. Она ясно показывает, что характер восходящей "неапецифической" импульсации определяется кортикофугальными импульсами. При осознании стимула эта дополнительная "эмоциональная" активация коры более локальна в пространстве и времени, больше приурочена к корковым структурам, воспринимающим данный раздражитель. Надо думать, что возбуждение более высоких структурно-функциональных уровней коры, обеспечивающее осознание внешнего раздражителя, каким-то образом изменяет характер нисходящих регулирующих влияний на лимбическую систему, а через нее и на корковые нейроны.
Психофизиологические исследования значительно чаще выявляют повышение порога опознания эмоциональных слов, чем их понижение, то сравнению с нейтральными словами. Однако в опытах с неосознаваемыми словами мы наблюдаем, как правило, облегчение вызванных потенциалов на эмоциональные слова. Это противоречие между психофизиологическими и биоэлектрическими данными в настоящее время можно объяснить только тем, что связанные с эмоциональным компонентом слова восходящие импульсы со стороны лимбической системы оказывают неодинаковое влияние на разные структурно-функциональные корковые уровни.
По-видимому, одна и та же импульсация, поступающая в кору из "неспецифических" систем, может тормозить функциональную систему, обеспечивающую вербализацию внешних явлений, и тем самым повышать порог опознания словесного раздражителя; с другой стороны, эта же импульсация повышает активность определенных корковых элементов, с которыми связана, в основном, генерализация вызванных потенциалов. О том, что источник биоэлектрических и перцептивных изменений-один-дополнительная импульсация из лимбической системы, - говорят наши исследования с применением антихолинергического вещества амизила [9]. Это вещество, действующее активно на лимбическую систему, временно нивелировало пороги опознания эмоциональных и нейтральных слов и устраняло разницу в амплитудно-временных параметрах вызванных потенциалов коры.
Каков физиологический смысл подобных разнонаправленных изменений корковых функций? По этому поводу в настоящее время могут быть высказаны лишь предположения. В случаях действия эмоционально неприятного стимула порог его осознания повышается, чем и достигается "психологическая защита". Вместе с тем, одновременно происходит активация корковых нейронов и, как мы видели, более диффузная, чем при действии осознанных эмоциональных раздражителей. Последним, по-видимому, кора головного мозга как бы подготавливается к осуществлению необходимых защитных реакций на еще не вполне опознанный стимул.
Итак, согласно нашим данным, зрительный словесный раздражитель, который не осознается исследуемым, так как он предъявляется ему на очень короткое время и очень слабый по яркости, в состоянии вызвать в коре головного мозга электрический ответ. Корковый ответ на такого рода стимул регистрируется обычно в виде усредненного позднего негативно-позитивного колебания, так называемых компонентов N 200 и Р 300. Существенная разница в амплитуде вызванного потенциала на нейтральные и эмоциональные слова дает нам основание считать, что и в случаях, когда словесный стимул не осознается, в коре головного мозга происходит анализ и синтез его семантических свойств. Ведь только после анализа и "опознания" предъявляемого слова корковыми элементами, воспринимающими зрительную речь, может произойти та дополнительная активация корковых нейронов, связанная с эмоциональным значением стимула, которую мы наблюдали в виде увеличения потенциала.
Следовательно, для возбуждения временных связей между новой корой и структурами лнмбической системы не обязательно, чтобы эмоциональный словесный раздражитель осознавался субъектом. Добавочная восходящая "неспецифическая" импульсация из лимбической системы к неокортексу, связанная с эмоциональным компонентом словесного стимула, может возникать еще до того, как слово будет осознано.
В опытах с вызванными потенциалами коры мы получили еще одно доказательство того, что при определенных условиях человек может воспринимать отдельные слова и реагировать на их смысловое содержание без их осознания. Но главное не в том, что получено еще одно подтверждение этого удивительного факта. Результаты регистрации вызванных потенциалов на неосознаваемые слова доказывают справедливость высказанной нами гипотезы о нейрофизиологическом механизме явления "психологической защиты" [7; 8]. Как уже описывалось выше, согласно этой точке зрения, повышение порога опознания эмоционального слова обусловлено тем, что оно еще на "досознательном уровне" воспринимается и "опознается" корой головного мозга. Будучи еще не осознанным, эмоциональный словесный стимул может возбуждать временные связи между воспринимающими элементами неокортек-са и лимбической системой. Кортикофугальное возбуждение структур лимбической системы приводит к возникновению неспецифической импульсации, которая изменяет уровень возбудимости коры головного мозга.
Этот корковый механизм саморегуляции, действующий по принципу обратной связи, благодаря которому кора головного мозга регулирует активность собственных нейронов, адекватно сигнальному значению действующих в данный момент раздражителей, и является, по нашему мнению, основой изменений порогов осознания эмоциональных раздражителей, в частности явления "психологической защиты".
54. Physiological Mechanisms of "Psychological Defence" and Unaccountable Emotions. E. A. Kostandov
Research Institute of Forensic Psychiatry, Moscow
Summary
A difference between the recognition thresholds of neutral and meaningful words has been found in persons in the state of emotional stress. With an increase in the threshold of recognition of affective words the subliminal effect was observed (autonomic and bioelectrical reactions). Visually evoked potentials (VEP) were recorded to subliminal words. Recording of VEP to two successive stimuli shows that associations can be elaborated between two unrecognized stimuli. The role of additional unspecific impulses from the limbic system in the neural mechanisms of changes of the thresholds of recognition of affective words is discussed.
Литература
1. Аствацатуров М. И., Современные неврологические данные о сущности эмоций. В сб.: Советская невропсихиатрия, Л., 1936, т. I, 33.
2. Баклаваджян О. Г., Вегетативная регуляция электрической активности мозга, 1967, Л., 84.
3. Беритов И. С, Нервные механизмы поведения высших позвоночных животных, М., 1961.
4. Бериташвили И. С, Память позвоночных животных, ее характеристика и происхождение, Тбилиси, 1968.
5. Беритов И. С, Структура и функции коры большого мозга, М., 1969.
6. Бериташвили И. С, Характеристика и происхождение памяти позвоночных животных. В сб.: Гагрские беседы, т. VI, Тбилиси, 1972, 165.
7. Костандов Э. А., Эффект неопознаваемых "эмоциональных" словесных раздражителей. Журн. высш. нервн. деят. им. И. П. Павлова, 1968, 18, 3, 371.
8. Костандов Э. А, Ориентировочный рефлекс. В кн.: Физиология высшей нервной деятельности, часть I, M., 1970, 206.
9. Костандов Э. А., Арзуманов Ю. Л., Изменения корковых вызванных потенциалов на эмоциональные зрительные стимулы под влиянием амизила у человека. Журн. высш. нервн. деят. им. И. П. Павлова, 1971, 21, 6, 1247.
10. Костандов Э. А., Арзуманов Ю. Л., Вызванные корковые потенциалы на эмоциональные неосознаваемые слова. Журн. высш. нервн. деят. им. И. П. Павлова, 1974, 24, 3, 465.
11. Маркс К. и Энгельс Ф., Сочинения, т. 3. М., 1955, 29.
12. Ройтбак А. И., Вызванные потенциалы коры больших полушарий. В кн.: Современные проблемы электрофизиологических исследований нервной системы. М., 1974, 164.
13. Тевзадзе В. Г., Реакции, вызванные прямым раздражением поясной извилины, и их влияние на некоторые функции. Сообщения АН ГССР, 1965, 43, 2, 487.
14. Хачапуридзе Б. И., Об отражательной функции установки в связи с проблемой воздействия невоспринимаемых раздражителей. Труды Тбилисского государственного университета, 1966, 124, 25.
15. Beck, Е. G; Dustman, R.E., Beiter. E. G., Hypnotic suggestion and visually evoked potentials. Electroenceph. Clin. Neurophysiol., 1966, 20, 4, 397.
16. Bergamini, L., Bergamasco, В., Possibility of the clinical use of sensory evoked potentials transcranially recorded in man. Electroenceph. Clin. Neurophysiol., Suppl., 1967, 26, 114.
17. Brady, J. V. (Брейди Дж.), Палеокортекс и мотивация поведения. В кн.: Механизмы целого мозга, М., 1963, 138.
18. Chi, С. С, Flynn, J. P., The effects of hypothalamic and reticular stimulation [of evoked responses in the visual system of the cat. Electroenceph. Clin. Neurophysiol., 1968, 24, 4, 343.
19. Dixon, N. F., Lear, Т. Е., Incidence of theta rhythm prior of awareness of a visual stimulus. Nature, 1964, 203, 4941, 167.
20. Edwards, A. E., Subliminal tachistoscopic perception as a function of threshold method. J. Psychol., 1960, 50, 139.
21. Eriksen, С W., Subception: fact or artifact? Psychol. Rev., 1956, 63, 74.
22. Fangel, Ch., Kaada, B. R., Behavior "attention" and fear induced by cortical stimulation in the cat. EEG Clin. Neurophysiol., 1960, 12, 3, 575.
23. Garcia Austt, E., Vanzulli, A., Evoked responses and perception changes. Electroenceph. Clin. Neurophysiol.. 1970, 28, 5, 521.
24. Gellhorn, E., Loofbourrow, G. (Гельгорн Э., Луфборроу Дж.), Эмоции и эмоциональные расстройства, М., 1966.
25. Goldiamond, I., Indicators "of perception: I. Subliminal perception, subception, unconscious perception: An analysis in terms of psychophysical indicator methodology. Psychol. Bull., 1958, 55, 6, 373.
26. Goldstein, M. J., Barthol, R. P., Fantasy responses to subliminal stimuli. J. Abnorm. Soc. Psychol., 1960, 60, 1, 22.
27. Goldstein, M. J., Himmelfarb, S. Z., The effects of providing knowledge of results upon the perceptual defense effect. J. Abnorm., Soc. Psychol., 1962, 64, 143.
28. Halliday, A. M., Mason, A. A., Cortical evoked potentials during hypnotic anaesthesia. Electroenceph. Clin. Neurophysiol., 1964, 16, 3, 314. '29. HOWES, D., SOLOMON, R. L., A Note on McGinnies, "Emotionality and Perceptu? Defence". Psychol. Rev., 1950, 57, 4, 229.
30. Howes, D., Solomon, R. L., Visual duration threshold as a function of word probability. J. Exp. Psychol., 1951, 41, 401.
31. John, E. R., Herrington, R. N., Sutton, S., Effects of visual form on the evoked response. Science, 1967, 155, 3768, 1439.
32. Johnson, R. C. Frincke, G., Martin, L.,Meaningfulness, frequency, and affective character of words as related to visual duration threshold. Canad. J. Psychol., 1961, 15(4), 199.
33. Lazarus, R. S., McCleary, R. A., Autonomic discrimination without awareness: A Study of subception. Psychol. Rev., 1951, 58, 113.
34. Levy, R., Behrman, J., Cortical evoked responses in hysterical hemianaesthesia, Electroenceph. Clin. Neurophysiol., 1970, 29, 4, 400.
35. Libet, В., Brain stimulation and the threshold of conscious experience. In: Brain and Conscious Experience, Berlin, Heidelberg, N.Y, 1966, 165.
36. Libet, В., Alberts, W. W., Wright, E. W., Feinstein, В., Responses of human somatosensory cortex to stimuli below threshold for conscious sensation, Science, 1967, 158, 1597.
37. Lorens, S.A., Brown, Th. S., Influence of stimulation of the septal area on visual evoked potentials. Exp. Neurol., 1967, 17, 1, 86.
38. McGinnies, E., Discussion of Howes' and Solomon's Note on "Emotionality and perceptual defense". Psychol. Rev., 1950, 57, 4, 235.
39. MacLean, P. D., The limbic brain in relation to the psychoses. In: Physiological Correlates of Emotion, N. Y., London, 1970, 130.
40. Moruzzi, G., Magoun, H., Brain stem reticular formation and activation of the EEG. EEG Clin. Neurophysiol., 1949,1,3,455.
41. Newbigging, P. L., The perceptual redintegration of frequent and infrequent words. Canad. J. Psychol., 1961, 15(3 ), 123. o42. NEWBIGGING, P. L., The perceptual redintegration of words which differ inconnotative meaning. Canad. J. Psychol., 1961, 15 (3), 133.
43. Olds, J. (Олдс Дж.), Выявление подкрепляющих систем головного мозга методом самораздражепия. В кн.: Механизмы целого мозга, М., 1963, 199.
44. Phares, E. J., Perceptual threshold decrements as a function of skill and chance expectancies. J. Physiol., 1962, 53, 399.
45. Penfield, K., Jasper, H. (Пенфилд У., Джаспер Г.), Эпилепсия и функцио -нальная анатомия головного мозга человека, М., 1958.
46. Riggs, L., Progress in the recording of human retinal and occipital potentials. J. Opt. Soc. Amer., 1969, 59, 12, 1558.
47. Riggs, L. A., Whittle, P., Human occipital and retinal potentials evoked by subjectively faded visual stimuli. Vision Res., 1967, 7, 441.
48. Smith, G. L., Spence, D. P., Klein, G. S., Subliminal effect of verbal stimuli. J. Abnorm. Soc. Psychol., 1959, 59, 2, 167.
49. Smith, G. J., Spence, D. P.. Klein. G. S., Subliminal effects of verbal stimuli. J. Abnorm. Soc. Psychol., 1959, 59, 2, 167.
50. Spence, D. P., Subliminal perception and perceptual defense: Two sides of a single problem. Behav Sci., 1967, 12, 3, 183.
51. Spence, D. P., Ehrenberg, B. Effects of oral deprivation on responses to subliminal and supraliminal verbal food stimuli. J. Abnorm. Soc, Psychol., 1964, 69, 10.
52. Sperry, R. W., Brain bisection and mechanisms of consciousness. In: Brain and Conscious Experience, Berlin, Heidelberg, N. Y., 1935, 298.
Дата: 2019-07-24, просмотров: 428.