Возникновение ЦНС из недифференцированных клеток эктодермы до настоящего времени остается одним из самых загадочных процессов в эмбриогенезе позвоночных. Считается, что исходно клетки эктодермы не имеют самостоятельного нейрогенного потенциала, а их дифференцировка в нервные клетки индуцируется какими-то сигналами, поступающими из нижележащих клеток эмбриональной мезодермы. Молекулярная природа этих сигналов и их первичных эффектов в клетках эктодермы до последнего времени была совершенно неизученной. Совсем недавно обнаружено, что, например, компонентом такого первичного эффекта в эмбрионах шпорцевых лягушек является индукция гена Х1НЬохб, содержащего характерную для многих генов, регулирующих онтогенез, эволюционно консервативную последовательность, так называемый гомеобокс. Он тесно связан с определением локализации групп клеток и тканей, с "географией" клеток и функций в организме.
Экспрессия этого гена характерна для наиболее ранних стадий нейрогенеза. Ген ХШЬохб имеет четко выраженный сегментарный характер экспрессии в эмбриональной ЦНС: его транскрипты обнаруживаются только в задних двух третях нервной трубки. Подобно многим гомейотическим генам дрозофилы ген ХШЬохб перестает экспрессироваться на поздних стадиях эмбриогенеза.
Ген XlHboxl экспрессируется в спинном мозге, причем передняя граница экспрессирующей его области - очень резкая и в точности совпадает с границей спинного и головного мозга. При инактивации кодируемого этим геном белка специфическими антителами нарушается развитие передних отделов спинного мозга: они трансформируются в структуру, морфологически идентичную продолговатому мозгу.
Очевидно, кодируемые этими генами белки являются специфическими транскрипционными факторами, регулирующими генетическую детерминацию клеточных линий в определенных отделах эмбриональной ЦНС. Они обеспечивают реализацию позиционной информации в общем процессе нейрогенеза.
В геноме мыши обнаружено четыре комплекса тесно сцепленных гомеобокс-содержащих генов, Нох-1, - 2, - 3, и - 4. Индивидуальные гены каждого комплекса аналогично гомейотическим генам дрозофилы экспрессируются в различных относительно передне-задней оси тела областях ЦНС. Морфологически различимые сегменты в эмбриональной ЦНС позвоночных обнаруживаются лишь на ранних стадиях развития заднего мозга в виде так называемых ромбомеров, тогда как на более поздних стадиях и в мозге взрослых животных рудиментом сегментарного строения остается лишь распределение ядер черепно-мозговых нервов.
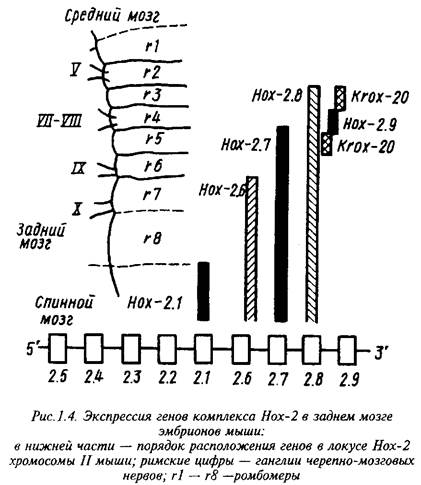
Индивидуальные гены Нох-2 комплекса имеют общую область экспрессии в каудальных отделах эмбриональной нервной трубки, тогда как передние границы экспрессирующей области у них различны: чем ниже данный ген расположен в Нох-2 локусе хромосомы, тем выше расположена эта граница.
Сейчас интенсивно накапливаются и обобщаются новые данные в этой области. Эволюционная консервативность регуляторных генов, экспрессируемых в сегментарных отделах эмбриональной ЦНС позвоночных, и явные аналогии в характере их экспрессии с гомейотическими генами дрозофилы делают весьма привлекательной гипотезу об их роли в детерминации общего плана строения и идентичности отдельных клеточных линий в развивающейся ЦНС.
Ни один из исследованных генов Нох комплексов не экс-прессируется в передних отделах головного мозга позвоночных в эмбриогенезе. Эти отделы не проявляют явных признаков сегментарного строения. Являясь относительно эволюционно новыми приобретениями, они, по-видимому, имеют специфичные механизмы генетической детерминации. Показано, в частности, что в детерминации отделов переднего и промежуточного мозга существенную роль играет ген Dlx, гомологичный гомейотическому гену D11 дрозофилы. Аналогично, в детерминации среднего мозга и, возможно, передних отделов мозжечка участвует протоонкоген Wnt-1 и гомеобокс-содержащие гены Еп-1 и - 2.
Имеются данные, заставляющие по-новому взглянуть на вопрос о функциях гомеобокс-содержащих генов у высших животных. Клонирование генов, кодирующих транскрипционные факторы Oct-1, Oct-2 и Pit-1, показало, что они являются членами особого подсемейства гомеобокс-содержащих генов, в которое входит также ген Unc-86 нематод. Общим признаком, объединяющим эти гены, является длинная консервативная последовательность, POU-домен, частью которой является гомеобокс. Ген Unc-86 участвует в детерминации нейронных линий у нематод. Несколько генов POU-подсемейства специфически экспрессируются в отделах мозга млекопитающих: Вгп-1, - 2, - 3 и Tst-I. Все четыре гена экспрессируются в эмбриональной нервной системе, включая клетки герминативных зон.
Динамика экспрессии индивидуальных генов в развивающемся мозге очень различна и по мере развития отделов мозга приобретает черты, отражающие их паттерн экспрессии в мозге взрослых животных. Экспрессия этих генов в клетках герминативных зон может означать, что судьба индивидуальных клеточных линий в нервной системе предопределена уже до начала их миграции из герминативной зоны.
По-видимому, кодируемые гомеобокс-содержащими генами транскрипционные факторы могут регулировать экспрессию генов не только в формирующихся клеточных линиях в раннем эмбриогенезе млекопитающих, но и в дифференцированных клетках взрослого организма, причем на очень разных уровнях клеточной специфичности.
Наконец, особого рассмотрения заслуживает тот факт, что в мозге млекопитающих есть ряд генов, особенно быстро реагирующих на разнообразные экстремальные воздействия. Их включение в ряде ситуаций, в частности, сопровождающихся повышением уровня внутриклеточного Са2+, служит как бы первым относительно неспецифичным звеном реакции генома нейрона. Далее следует включение более специфичных участков генома. В число таких генов "первоочередного реагирования" входят гены, открытые ранее в связи с исследованием протоонкогенов. В частности, гены c-fos и c-jun кодируют факторы - регуляторы транскрипции других генов, - белки Fos и Jun. Последние входят, в свою очередь, в состав белка АР-1, связывающегося с определенными локусами ДНК, - например возле 5'-конца гена, кодирующего фактор трофики нейронов - NGF. В экстремальных ситуациях - при эпилептических судорогах, требующих срочного усиления трофики нейронов, - этот механизм включает синтез NGF. Аналогичные механизмы с участием генов c-fos и c-jun срабатывают под влиянием болевых импульсов и других экстремальных воздействий на ЦНС, включая целые каскады транскрипции генов, кодирующих, например, такие противоболевые факторы, как опиодные пептиды и другие регуляторы. В последнее время c-fos и другие гены раннего реагирования нередко называют "третьим мессенджером". Наконец, появляется все больше данных об участии генов этого типа в механизмах запоминания.
Выводы
1. Большинство нейронов ЦНС позвоночных являются диплоидными; небольшая доля нейронов в некоторых отделах ЦНС может содержать избыточное по сравнению с диплоидным количество ДНК.
2. Репликативный синтез ДНК в дифференцированных нейронах отсутствует; в мозге взрослых млекопитающих репликация ДНК связана главным образом с ограниченными процессами размножения глиальных клеток.
3. В клетках мозга млекопитающих имеются активно функционирующие системы репарации ДНК, поддерживающие целостность и эффективность генетического аппарата.
4. Хроматин нервных клеток имеет типичную для эукариотических клеток нуклеосомную организацию. Особенностями хроматина нейронов неокортекса млекопитающих являются необычно короткие нуклеосомные единицы, присутствие редких вариантов гистонов, высокое разнообразие негистоновых белков и высокая матричная активность.
5. В мозге млекопитающих экспрессируется несколько десятков тысяч уникальных генов, из которых не менее половины имеют мозгоспецифический характер экспрессии.
6. Огромное разнообразие экспрессируемых в мозге генов складывается из перекрывающихся, но не одинаковых популяций генов, экспрессируемых в отдельных нервных клетках.
7. Наряду с разнообразнейшими мозгоспецифическими мессенджер-РНК в центральной нервной системе синтезируется ограниченное число особых малых РНК с последовательностями нуклеотидов, общими для всех отделов мозга.
8. Разнообразие белков и регуляторных пептидов, синтезируемых в мозге, определяется не только большим набором экспрессируемых генов, но и системой альтернативного сплайсинга пре-мРНК.
Дата: 2019-05-29, просмотров: 437.