С помощью ДНК-РНК-гибридизации показано, что в ЦНС моллюска-кальмара экспрессируется 46% уникальных последовательностей генома, что достаточно для кодирования нескольких десятков тысяч различных мРНК. Следовательно, генетическая сложность клеточных элементов в ЦНС высокоорганизованных беспозвоночных сопоставима с таковой в ЦНС млекопитающих. В отличие от млекопитающих, однако, у кальмара не обнаружено специфической для ЦНС популяции молекул поли+РНК.
Уникальную возможность для исследования экспрессии генов в индивидуальных нейронах представляет ЦНС брюхоногих моллюсков, состоящая из небольшого числа крупных, во многих случаях легко идентифицируемых нейронов, сосредоточенных в нескольких ганглиях. У аплизии некоторые нейроны достигают размеров 0,5 мм и содержат - 0,25 мкг ДНК и - 5 нг поли+РНК, что вполне достаточно для анализа методами молекулярного клонирования.
С помощью процедуры дифференциальной гибридизации клонированы гены и мРНК, специфически зкспрессирующиеся в отдельных нейронах ЦНС аплизии. В их число входят гены, продукты которых хорошо идентифицированы; например ген, кодирующий гормон откладки яиц, и четыре родственных ему гена. Будучи активными в разных нейронах, они кодируют синтез нескольких физиологически активных пептидов, секреция которых индуцирует стереотипный поведенческий репертуар откладки яиц. При этом один и тот же пептид может действовать как нейрогормон на клетки соматических тканей и нейромедиатор на определенные нейроны. Экспрессия других генов этого семейства в клетках атриальных желез приводит к синтезу нейроактивных пептидов, вызывающих активацию сумчатых клеток, а также секретируемых во внешнюю среду пептидов, обладающих активностью половых феромонов. Таким образом, кодируемые этим семейством генов пептиды регулируют различные компоненты одного сложного поведенческого репертуара.
Очень интересен также наплизии, участвующий в регуляции водно-соленого баланса. Он кодирует две мРНК, образующиеся в результате альтернативного сплайсинга. Одна из них является преобладающим продуктом экспрессии гена в одном из нейронов, тогда как укорочения форма мРНК преимущественно синтезируется в некоторых других нейронах. Следовательно, у аплизии существуют механизмы выбора различных путей сплайсинга в разных нейронах.
Изучение экспрессии генов в индивидуальных нейронах этого моллюска позволяет сделать некоторые принципиальные выводы, которые могут быть применимыми и к ЦНС позвоночных животных. Главный из этих выводов состоит в том, что присутствие множества "редких" молекул РНК в суммарных препаратах поли*РНК, изолированных из целой ЦНС или ее крупных отделов, является следствием активной экспрессии этих РНК в небольших популяциях нервных клеток, а не их экспрессии на одинаково низком уровне в обширных популяциях клеток.
Сравнение популяций мРНК, синтезируемых в нейронах, использующих один и тот же классический медиатор, но функционально различных, показывает, что такие нейроны обычно различаются экспрессией нескольких мРНК, каждая из которых специфична лишь для одного из сравниваемых нейронов. Очевидно, общее разнообразие синтезируемых в ЦНС мРНК складывается из перекрывающихся, но не одинаковых популяций мРНК, образуемых в индивидуальных нейронах. Остается добавить, что рассмотренные исследования охватывают лишь относительно часто встречающиеся молекулы мРНК. Не исключено, что в действительности различия в популяциях синтезируемых мРНК между индивидуальными нейронами носят более сложный характер.
К популярным объектам нейрогенетики из числа беспозвоночных относится и одна из свободно живущих нематод - Caenorhabditis elegans. Одним из важных результатов ее исследования является обнаружение так называемых селекторных генов. Последние играют ключевую роль в онтогенезе нейронов нематоды: их продукты индуцируют включение серии "генов-реализаторов", формирующих фенотип нейронов. Примером селекторного гена может служить ген INS-4, детерминирующий специфичность синаптических контактов идентифицируемого мотонейрона VA. При мутациях этого гена мотонейрон VA образует синаптические контакты, в норме ему не свойственные. Никаких других заметных нарушений в развитии ЦНС при этом не обнаруживается. Продуктом гена INS-4 является гомеобокс-содержащий ДНК-связывающий белок. Очевидно, он регулирует транскрипцию группы генов-реализаторов, непосредственно участвующих в установлении специфических для данного мотонейрона синаптических контактов.
Наконец, классический объект генетических исследований - дрозофила - позволил получить обширную информацию о сложных процессах онтогенеза центральной нервной системы. Эту информацию можно обобщенно представить в виде модели многоступенчатой генетической детерминации нейрогенеза, основные положения которой, по-видимому, сохраняют силу и для позвоночных животных. В соответствии с этой моделью существует четыре класса генов, регулирующих последовательные этапы нейрогенеза. Первый, наименее изученный класс, включает гены, ответственные за преобразование части клеток недифференцированной вентральной эктодермы в предшественники нервных клеток. Ко второму классу относятся гены, детерминирующие свойства клеток-предшественников в соответствии с их положением в той или иной части ЦНС. Среди этих генов - многие из так называемых гомейотических или сегментарных генов, которые детерминируют также и формирование общего плана строений организма в целом. При этом одни и те же гены обычно детерминируют и формирование определенного сегмента организма, и специфические свойства локализованного в нем сегмента ЦНС. Заметим, однако, что сегментарная и нейроспецифическая экспрессия гомейотических генов контролируется разными механизмами. Гены третьего класса детерминируют свойства индивидуальных нейробластов в зависимости от их локализации внутри данного сегмента ЦНС. Представителем этого класса, по-видимому, является ген prospero, экспрессирующийся только в определенных нейробластах каждого сегмента ЦНС, но не в образующихся из них нейронах. Мутации этого гена приводят к выпадению из развивающейся нервной системы нескольких специфических клеточных линий. Наконец, гены четвертого класса детерминируют индивидуальные свойства каждого из нейронов, образующегося при делении одного нейробласта. Представителями этого класса являются гены fushi tarazu и even-skipped. Интересно, что один и тот же ген может участвовать в контроле разных этапов нейрогенеза. Так, ген ftz является типичным гомейотическим геном, участвующим в формировании общего сегментарного плана строения ЦНС и организма в целом, т.е. входит во второй класс генов. На более поздних этапах он участвует в детерминации свойств индивидуальных нейронов: в каждом сегменте ЦНС он экспрессируется в группе из 30 идентифицируемых нейронов, являющихся потомками - 8 нейробластов. При этом ни в одном из нейробластов ген ftz не экспрессируется, а процент экспрессирующих его клеток в потомстве индивидуальных нейробластов варьирует от 0 до 100%. Заметим, однако, что сегментарная и нейроспецифическая экспрессия гена ftz контролируется разными механизмами. Еще один фундаментальный вывод описанных исследований состоит в том, что индивидуальная специфичность элементов нервной системы определяется не уникальными генами, количество которых заведомо меньше числа самих элементов, а уникальными комбинациями взаимодействующих генов, надежность которых обеспечивается чертами функциональной избыточности.
Гены, ответственные за формирование специфического фенотипа функционально зрелых нейронов, изучены главным образом для относительно просто устроенных нейронных структур, например сетчатки глаза дрозофилы. Каждый элемент сетчатки - омматидий - состоит из 8 четко различимых по морфологии и локализации фоточувствительных нейронов и 12 ненейрональных клеток. Дифференцировка нейронов в развивающихся омматидиях протекает в строго фиксированной последовательности. Недифференцированные клетки в зачатках омматидиев являются эквипотенциальными, а судьба дифференцирующихся клеток определяется не их происхождением, а индуцирующими стимулами со стороны соседствующих с ними ранее дифференцированных клеток.
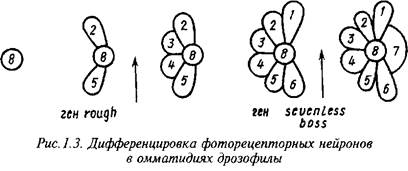
Некоторые из генов, ответственных за генерацию и рецепцию этих стимулов, хорошо изучены: ген seveniess кодирует мембранный рецептор, необходимый для индукции дифференцировки нейрона R7, а ген bride of sevenless - за образование в нейроне R8 лиганда, действующего на этот рецептор. Ген seven in absentia кодирует ядерный белок, интерпретирующий сигнал, воспринимаемый рецептором sevenless. Ген rough кодирует транскрипционный фактор, необходимый для возникновения в нейронах R2 и R6 сигнала, индуцирующего дифференцировку соседних клеток в нейроны R3 и R4. Другой транскрипционный фактор, необходимый для дифференцировки нейронов Rl, R3, R4 и R6, кодируется геном seven-up. Ключевым элементом в реализации генетических программ функционального созревания фоторецепторных нейронов является ДНК-связывающий белок, кодируемый геном glass, а специфическими для индивидуалъньгх фоторецепторных нейронов компонентами этих программ - гены, кодирующие разные формы фоторецепторных белков.
Важную роль в окончательной морфологической дифферен-цировке нейронов играют гены, кодирующие молекулы избирательной межклеточной адгезии. Одной из разновидностей таких молекул у насекомых являются фасциклины, обеспечивающие взаимное узнавание и ассоциацию растущих аксонов в развивающейся ЦНС и, в конечном итоге, формирование стереотипной системы аксонных пучков. В эмбриональной ЦНС разные фасциклины экспрессируются в разных популяциях нейронов и их отростков: каждый фасцикяин экспрессируется в аксонах - 5 из - 30 аксонных пучков передних и задних комиссур каждого ганглия и - 15 нейронах каждого полусегмента. В эмбриогенезе экспрессия фасциклинов наблюдается уже в отдельных клетках морфологически недифференцированной вентральной нейроэктодермы, сохраняясь в образующихся из этих клеток нейробластах, ганглионарных материнских клетках и нейронах. По-видимому, фасциклины входят в число наиболее ранних молекулярных маркеров индивидуальных нейрональньгх линий. Обнаруженные закономерности при исследовании относительно просто устроенной ЦНС беспозвоночных не всегда находят прямые аналогии в более сложно организованных ЦНС высших позвоночных животных. Тем не менее с известной осторожностью их можно использовать при интерпретации данных по экспрессии генов в мозге позвоночных.
Дата: 2019-05-29, просмотров: 433.