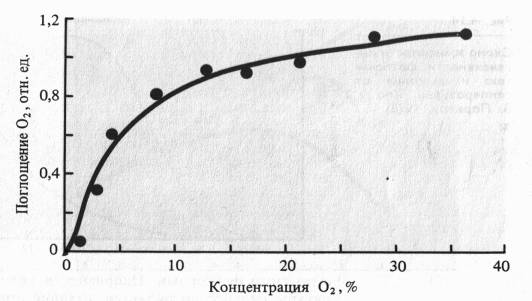
Концентрация 02. Процесс дыхания связан с непрерывным потреблением кислорода клетками и тканями растений. В то же время окислительные превращения субстратов при дыхании включают не только аэробные, но и анаэробные процессы (гликолиз, брожение). Поэтому интересен тот факт, что при снижении парциального давления 02 с 21 до 9% интенсивность дыхания тканей растений меняется незначительно. При 5% 02 у молодых растений начинает несколько снижаться поглощение 02 и в меньшей степени изменяется выделение С02. Как отмечалось, одна из важнейших конечных оксидаз аэробного дыхания — цитохромоксидаза — обладает высоким сродством к кислороду и может функционировать при его низких парциальных давлениях.
Эти факты можно объяснить как тем, что в эволюции дыхательные системы растений формировались в низкокислородных условиях, так и тем, что возможности функционирования аэробной дыхательной цепи при низких парциальных давлениях 02 — один из важных адаптивных признаков, элемент надежности энергетики растения. Это тем более важно, что газовый состав внутренних тканей органов растений может отличаться от атмосферного. Так, в листовой паренхиме сахарной свеклы содержание 02 меняется в течение суток от 7,1 до 17,4%, а С02 - от 0,9 до 5,1%; в мякоти зрелых яблок содержится 7,5 С02 и 13,9% 02, а плодов лимона — 8,5 С02 и 11,5% 02.
У растений, обитающих в условиях систематического затопления (болотные и др.), существует ряд приспособлений к перенесению недостатка кислорода: развитие аэренхимы, способность использовать кислород нитратов («нитратное» дыхание), различные способы устранения избытка продуктов брожения (этанола, молочной кислоты), а также механизмы их использования на нужды обмена веществ.
Избыток кислорода в тканях растений может возникать лишь локально. В атмосфере чистого кислорода дыхание растений снижается, а при длительном его действии растение погибает. Эффект избытка 02 связан с усилением в клетках свободнорадикальных реакций, повреждением мембран вследствие окисления их липидов и в конечном итоге — с нарушением многих процессов обмена веществ.
Диоксид углерода. Повышение концентрации С02 как конечного продукта дыхания приводит к снижению интенсивности дыхания. При повышении концентрации С02 тормозятся реакции декарбоксилирования и активность сукцинатдегидрогеназы и, следовательно, уменьшаются выделение С02 и дыхательный коэффициент. При этом наблюдается закисление тканей — ацидоз, что может приводить к вредным последствиям. С02 хорошо растворяется в липидах и таким образом может воздействовать на мембраны. По-видимому, он обладает некоторым наркотическим действием. Предполагается, что С02 способен регулировать метаболизм растений в анаэробных условиях. У листьев ингибирующее действие высоких концентраций С02 на дыхание может быть связано с закрыванием устьиц в этих условиях. Повышенное содержание С02 в тканях семян, покрытых плотной оболочкой, — один из способов поддержания состояния покоя.
Температура. Дыхание, подобно другим ферментативным процессам, зависит от температуры. В определенных температурных границах эта зависимость подчиняется правилу Вант-Гоффа (скорость химических реакций удваивается при повышении температуры на 10 °С). В интервале температур от 0 до 20 °С <210 дыхания равен 2 — 3. При температурах выше 20 °С величина Ql0 может понижаться. Одна из причин этого — уменьшение растворимости 02 в жидкостях при повышении темпепатупы. ЭАсЬект темпепатупы проявляется обычно вместе с другими факторами. Например, в условиях высокой температуры сильнее подавляется дыхание при пониженном содержании 02 или повышенной концентрации С02 и т. д.
Для дыхания каждого вида растений и его органов существуют определенные минимальные (когда дыхание еще возможно), оптимальные и максимальные температуры. В отличие от фотосинтеза дыхание у зимующих растений наблюдается при очень низких температурах (у хвои ели и сосны при — 25 °С). Температурный оптимум его у большинства видов умеренных широт лежит в пределах 35 — 40 °С, т. е. на 5 — 10 °С выше, чем для фотосинтеза. Максимальные температуры (45 — 55 °С) определяются способностью белков к денатурации.
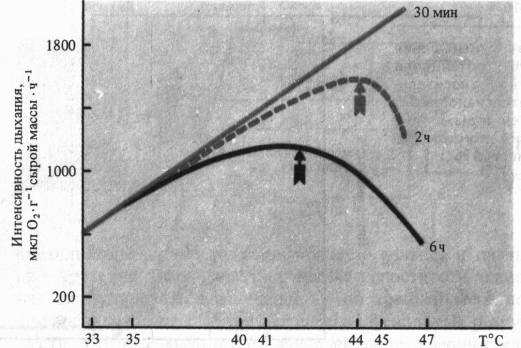
При рассмотрении влияния на дыхание крайних температур важно учитывать продолжительность действия этого неблагоприятного фактора. Так, при кратковременных экспозициях (30 мин) температурный оптимум дыхания листьев гречихи сахалинской может быть очень высок (> 47 °С), тогда как при двухчасовом действии оптимум смещается к 44 °С, а при шестичасовом—до 42 °С.
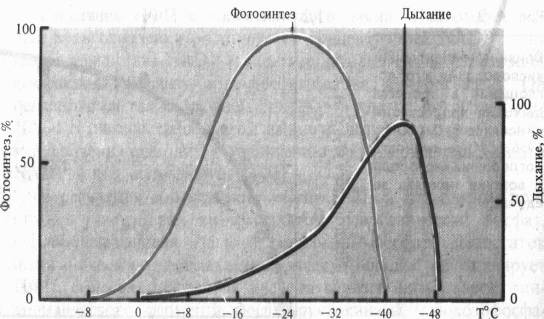
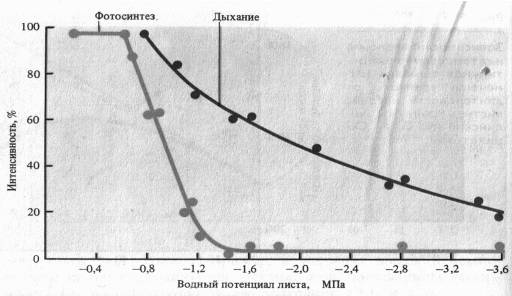
Водный режим. Изменение оводненности тканей растения отражается на интенсивности дыхания. В листьях проростков при быстрой потере воды вначале отмечается усиление дыхания — обычная кратковременная реакция на раздражение. При постепенном снижений оводненности этого не происходит. Продолжающийся водный дефицит вызывает постепенное снижение дыхания, причем более медленное, чем падение интенсивности фотосинтеза. Более раннее прекращение фотосинтетической активности при водном дефиците является следствием закрытия устьиц (уменьшение поступления С02 в лист) и потери тургорного давления в клетках.
Необходимость определенной степени оводненности тканей для процесса дыхания отчетливо проявляется при изучении дыхания семян. Дыхание воздушно-сухих семян (10—11% Н20) ничтожно мало. При повышении влажности семян до 14—15% дыхание возрастает в 4 —5 раз, а при дальнейшем повышении оводненности семян до 30 — 35% дыхание увеличивается в тысячи раз. При этом важную роль играет температура, при которой хранятся семена: при 0— 10°С влияние влажности на интенсивность дыхания значительно меньше, чем при 18 — 25 °С. Резкий подъем дыхания набухших семян сопровождается значительным выделением тепла, что может привести к их перегреву («самовозгоранию») при хранении.
Реакция растительных тканей на потерю воды зависит также от возраста, физиологического состояния, экологической принадлежности организма.
Минеральные вещества. Добавление раствора солей в воду, на которой выращивают проростки, обычно усиливает дыхание корней. Этот эффект получил название «солевого дыхания». Полагают, что солевое дыхание отражает усиление обмена веществ, необходимое для энергетической поддержки активного транспорта ионов корнем. Однако до конца это явление не изучено. Отмечены случаи, когда увеличенная интенсивность дыхания сохранялась после удаления солей. В тканях других органов этот эффект удается получить не всегда.
Свет. Действие света на дыхание зеленых органов растений из-за методических трудностей изучено недостаточно, так как одновременно с дыханием в них осуществляется фотосинтез. Освещенность, при которой интенсивность фотосинтеза равна интенсивности дыхания (по уровню поглощенного и выделенного С От), называется компенсационным пунктом.
При работе с незелеными растениями или с незелеными органами растений возможно непосредственное изучение действия света на дыхание. Дыхание (поглощение 02 и выделение С02) незеленых тканей активируется светом коротковолновой части спектра — ближним ультрафиолетом (380 нм), синими и зелеными лучами (400 — 500 нм) .
В области 380 — 600 нм лежат максимумы поглощения каротиноидов, флавинов и всех цитохромов дыхательной цепи митохондрий. Усиление дыхания может быть связано с прямым действием света на эти компоненты окислительно-восстановительных систем клеток.
Остается нерешенным вопрос о характере взаимодействия процессов фотосинтеза и дыхания в зеленых клетках в ходе фотосинтеза. Обсуждаются две точки зрения об использовании АТР, образующегося при фотофосфорилировании. Согласно одной из них на свету хлоропласта большую часть синтезируемой при фотофосфорилировании АТР используют на синтез углеводов, отдавая на нужды клетки лишь небольшую ее часть. АТР в цитоплазму поставляют митохондрии, которые используют ассимиляты. В темноте хлоропласта сами нуждаются в поступлении энергии из цитоплазмы.
По другой гипотезе АТР фотосинтеза на свету обеспечивает энергией не только фиксацию С02, но и другие энергозависимые процессы в клетке. Окислительное фосфорилирование в митохондриях на свету резко замедляется, а цикл Кребса ускоряется, так как его интермедиаты необходимы для функционирования хлоропластов.
Список использованной литературы
1.В.В. Полевой Физиология растений М.: Высшая школа 1989г.
2.Н.И. Якушкина Физиология растений М.: Просвещение 1993г.
Дата: 2019-05-29, просмотров: 336.