1. Рефлекс (от лат. reflexus — отражённый) — стереотипная реакция живого организма на раздражение рецепторов, проходящая с участием нервной системы. Рефлексы существуют у многоклеточных живых организмов, обладающих нервной системой, осуществляются посредством рефлекторной дуги.[1] Рефлекс — основная форма деятельности нервной системы.
Положения рефлекторной теории, разработанные И. М. Сеченовым. И. П. Павловым и развитые Н. Е. Введенским. А. А. Ухтомским. В. М. Бехтеревым, П. К. Анохиным и другими физиологами, являются научно-теоретической основой советской физиологии и психологии. Эти положения находят свое творческое развитие в исследованиях советских физиологов и психологов.
Рефлекторная теория, признающая рефлекторную сущность деятельности нервной системы, основывается на трех главных принципах:
1) принципе материалистического детерминизма;
2) принципе структурности;
3) принципе анализа и синтеза.
Принцип материалистического детерминизма означает, что каждый нервный процесс в головном мозге обусловливается (вызывается) действием определенных раздражителей.
Принцип структурности заключается в том, что различия функций разных отделов нервной системы зависят от особенностей их строения, а изменение строения отделов нервной системы в процессе развития обусловливается изменением функций. Так, у животных, которые не имеют головного мозга, высшая нервная деятельность отличается значительно большей примитивностью по сравнению с высшей нервной деятельностью животных, у которых есть головной мозг. У человека в ходе исторического развития головной мозг достиг особенно сложного строения и совершенства, что связано с его трудовой деятельностью и общественными условиями жизни, требующими постоянного речевого общения.
Принцип анализа и синтеза выражается в следующем. При поступлении в центральную нервную систему центростремительных импульсов в одних нейронах возникает возбуждение, в других — торможение, т. е. происходит физиологический анализ. Результатом является различение конкретных предметов и явлений действительности и процессов, происходящих внутри организма.
Одновременно при образовании условного рефлекса устанавливается временная нервная связь (замыкание) между двумя очагами возбуждения, что физиологически выражает собой синтез. Условный рефлекс есть единство анализа и синтеза.
Рефлекс - понятие, его роль и значение в организме
Рефлексами называют ответные реакции организма на раздражение рецепторов. В рецепторах возникают нервные импульсы, которые по чувствующим (центростремительным) нейронам поступают в центральную нервную систему. Там полученная информация обрабатывается вставочными нейронами, после чего возбуждаются двигательные (центробежные) нейроны и нервные импульсы приводят в действие исполнительные органы — мышцы или железы. Вставочными называют нейроны, тела и отростки которых, не выходят за пределы центральной нервной системы. Путь, по которому проходят нервные импульсы от рецептора до исполнительного органа, называется рефлекторной дугой.
18. Рефлекторная дуга, её звенья. Представление об обратной связи.
Рефлекторная дуга (нервная дуга) — путь, проходимый нервными импульсами при осуществлении рефлекса Рефлекторная дуга состоит из: рецептора — нервное звено, воспринимающее раздражение; афферентного звена
— центростремительное нервное волокно — отростки рецепторных нейронов, осуществляющие передачу импульсов от чувствительных нервных окончаний в центральную нервную систему;
центрального звена — нервный центр (необязательный элемент, например для аксон-рефлекса); эфферентного звена — осуществляют передачу от нервного центра к эффектору, эффектора — исполнительный орган, деятельность которого изменяется в результате рефлекса.
Различают: — моносинаптические, двухнейронные рефлекторные дуги; — полисинаптические рефлекторные
дуги (включают три и более нейронов).
Полисинаптическая рефлекторная дуга: нервный импульс от рецептора передаётся по чувствительному (афферентному) нейрону в спинной мозг. Клеточное тело чувствительного нейрона расположено в спинальном ганглии вне спинного мозга. Аксон чувствительного нейрона в сером веществе мозга связан посредством синапсов с одним или несколькими вставочными нейронами, которые, в свою очередь, связаны с дендритами моторного (эфферентного) нейрона. Аксон последнего передаёт сигнал от вентрального корешка на эффектор (мышцу или железу).
Простейшая рефлекторная дуга у человека образована двумя нейронами — сенсорным и двигательным (мотонейрон). Примером простейшего рефлекса может служить коленный рефлекс. В других случаях в рефлекторную дугу включены три (и более) нейрона — сенсорный, вставочный и двигательный. В упрощенном виде таков рефлекс, возникающий при уколе пальца булавкой. Это спинальный рефлекс, его дуга проходит не через головной, а через спинной мозг. Отростки сенсорных нейронов входят в спинной мозг в составе заднего корешка, а отростки двигательных нейронов выходят из спинного мозга в составе переднего. Тела сенсорных нейронов находятся в спинномозговом узле заднего корешка (в дорсальном ганглии), а вставочных и двигательных — в сером веществе спинного мозга. Простая рефлекторная дуга, описанная выше, позволяет человеку автоматически
(непроизвольно) адаптироваться к изменениям окружающей среды, например, отдергивать руку от болевого раздражителя, изменять размеры зрачка в зависимости от условий освещенности. Также она помогает регулировать процессы, протекающие внутри организма. Все это способствует сохранению постоянства внутренней среды, то есть поддержанию гомеостаза. Во многих случаях сенсорный нейрон передает информацию (обычно через несколько вставочных нейронов) в головной мозг. Головной мозг обрабатывает поступающую сенсорную информацию и накапливает её для последующего использования. Наряду с этим головной мозг может посылать моторные нервные импульсы по нисходящему пути непосредственно к спинальным мотонейронам; спинальные мотонейроны инициируют ответ эффектора.
Взаимосвязь, определяющая процессы возбуждения и торможения, т.е. саморегуляции функций организма, осуществляется при помощи прямых и обратных связей между центральной нервной системой и исполнительным органом. ОБРАТНАЯ СВЯЗЬ ("обратная афферентация" по П.К.Анохину), т.е. связь между исполнительным органом и центральной нервной системой, подразумевает передачу сигналов с рабочего органа в центральную нервную систему о результатах его работы в каждый данный момент.
Согласно обратной афферентации, после получения исполнительным органом эфферентного импульса и выполнения рабочего эффекта, исполнительный орган сигнализирует центральной нервной системе о выполнении приказа на периферии.
Существование такой замкнутой кольцевой, или круговой, цепи рефлексов центральной нервной системы и обеспечивает все сложнейшие коррекции протекающих в организме процессов при любых изменениях внутренних и внешних условий (В.Д. Моисеев, 1960). Без механизмов обратной связи живые организмы не смогли бы разумно приспособиться к окружающей среде.
Следовательно, вместо прежнего представления о том, что в основе строения и функции нервной системы лежит разомкнутая рефлекторная дуга, теория информации и обратной связи ("обратной афферентации") дает новое представление о замкнутой кольцевой цепи рефлексов, о круговой системе эфферентно-афферентной сигнализации. Не разомкнутая дуга, а сомкнутый круг - таково новейшее представление о строении и функции нервной системы.
19. Понятие о нервном центре. Особенности проведения возбуждения по нервным центрам.
Нервным центром называют совокупность нервных клеток, необходимых для осуществления какой-либо функции. Эти центры отвечают соответствующими рефлекторными реакциями на внешнее раздражение, поступившее от связанных с ними рецепторов. Клетки нервных центров реагируют и на непосредственное их раздражение веществами, находящимися в протекающей через них крови (гуморальные влияния). В целостном организме имеется строгое согласование — координация их деятельности.
Проведение волны возбуждения от одного нейрона к другому через синапс происходит в большинстве нервных клеток химическим путем — с помощью медиатора, а медиатор содержится лишь в пресинаптической части синапса и отсутствует в постсинаптической мембране. Поэтому важной особенностью проведения возбуждения через синоптические контакты является одностороннее проведение нервных влияний, которое возможно лишь от пресинаптической мембраны к постсинаптической и невозможно в обратном направлении. В связи с этим поток нервных импульсов в рефлекторной дуге имеет определенное направление от афферентных нейронов к вставочным и затем к эфферентным — мотонейронам или вегетативным нейронам.
Большое значение в деятельности нервной системы имеет другая особенность проведения возбуждения через синапсы — замедленное проведение. Затрата времени на процессы, происходящие от момента подхода нервного импульса к пресинаптической мембране до появления в постсинаптической мембране потенциалов, называется синаптической задержкой. В большинстве центральных нейронов она составляет около 0.3 мс. После этого требуется еще время на развитие возбуждающего постсинаптического потенциала (ВПСП) и потенциала действия. Весь процесс передачи нервного импульса (от потенциала действия одной клетки до потенциала действия следующей клетки) через один синапс занимает примерно 1.5 мс. При утомлении, охлаждении и ряде других воздействий длительность синаптической задержки возрастает. Если же для осуществления какой-либо реакции требуется участие большого числа нейронов (многих сотен и даже тысяч), то суммарная величина задержки проведения по нервным центрам может составить десятые доли секунды и даже целые секунды.
При рефлекторной деятельности общее время от момента нанесения внешнего раздражения до появления ответной реакции организма — так называемое скрытое или латентное время рефлекса определяется в основном длительностью проведения через синапсы. Величина латентного времени рефлекса служит важным показателем функционального состояния нервных центров
20. Классификация синапсов в ЦИС: возбуждающие и тормозящие; их медиаторы.
Синапс - специализированный контакт между нервными клетками или нервными клетками и другими возбудимыми образованиями, обеспечивающий передачу возбуждения с сохранением его информационной значимости. С помощью синапсов осуществляется взаимодействие разнородных по функциям тканей организма, например, нервной и мышечной, нервной и секреторной. Синаптическая область характеризуется специфическими химическими свойствами. Понятие «синапс» ввел в 1897 г. английский физиолог Шеррингтон, обозначив так соединение аксона одной нервной клетки с телом другой.
Все синапсы имеют принципиально общие черты строения. Пресинаптическое окончание аксона нейрона при подходе к иннервируемой клетке теряет миелиновую оболочку, что несколько снижает скорость распространения волны возбуждения. Небольшое утолщение на конце волокна, называемое синаптической бляшкой, содержит синаптические пузырьки с медиатором -веществом, способствующим передаче возбуждения в синапсе.
Классификация синапсов
В основу классификации синапсов положены три основных принципа. В соответствии с морфологическим принципом синапсы подразделяют:
- аксоаксональные синапсы (между двумя аксонами);
- аксодендритические синапсы (между аксоном одного нейрона и дендритом другого);
- аксосоматические синапсы (между аксоном одного нейрона и телом другого);
- дендродендритические (между дендритами двух или нескольких нейронов);
- нервно-мышечные синапсы (между аксоном мотонейрона и исчерченным мышечным волокном);
- аксоэпителиальные синапсы (между секреторным нервным волокном и гранулоцитом);
- межнейронные синапсы (общее название синапсов между какими-либо элементами двух нейронов).
Кроме этого, все синапсы делят на центральные (в головном и спинном мозге) и периферические
(нервно-мышечные, аксоэпителиальные и синапсы вегетативных ганглиев).
В соответствии с нейрохимическим принципом синапсы классифицируют по виду химического вещества - медиатора, с помощью которого происходит возбуждение и торможение эффекторной клетки. В адренер-гическом синапсе медиатором является адреналин, в холинергическом синапсе - ацетилхолин, а в гамкергическом синапсе - гамма-аминомасляная кислота и др.
По способу передачи возбуждения синапсы подразделяют на три группы. Первую составляют синапсы с химической природой передачи посредством медиаторов (например, нервно-мышечные); вторую - синапсы с передачей электрического сигнала непосредственно с пресинаптической - на постсииаптическую мембрану (например, синапсы в клетчатке глаза). По сравнению с химическими синапсами они отличаются большей скоростью передачи сигнала, высокой надежностью и возможностью двусторонней передачи возбуждения. Третья группа представлена «смешанными» синапсами, сочетающими элементы как химической, так и электрической передачи.
По конечному физиологическому эффекту, а также по изменению потенциала постсинаптической мембраны различают возбуждающие и тормозные синапсы. В возбуждающих синапсах в результате деполяризации постсинаптической мембраны генерируется возбуждающий постсинапти-ческий потенциал (ВПСП). В тормозных синапсах возможны два варианта процесса:
- в пресинаптических окончаниях выделяется медиатор, гиперполя-ризующий постсииаптическую мембрану и вызывающий в ней тормозной постсинаптический потенциал (ТПСП);
- тормозной синапс является аксоаксональным, т.е. еще до перехода возбуждения на область синапса обеспечивает пресииаптическое торможение.
21. Суммация возбуждения в ЦНС, её виды (И.М. Сеченов).

 В нервном волокне каждое одиночное раздражение (если оно не подпороговой и не свехпороговой силы) вызывает один импульс возбуждения. В нервных же центрах, как показал впервые И.М.Сеченов, одиночный импульс в афферентных волокнах обычно не вызывает возбуждения, т.е. не передается на эфферентные нейроны. Чтобы вызвать рефлекс необходимо быстрое нанесение допороговых раздражений одно за другим. Это явление получило название временной или последовательной суммации. Ее сущность состоит в следующем. Квант медиатора, выбрасываемого окончанием аксона при нанесении одного допорогового раздражения, слишком мал для того, чтобы вызвать возбуждающий постсинаптический потенциал, достаточный для критической деполяризации мембраны. Если же к одному и тому же синапсу идут быстро следующие один за другим допороговые импульсы, происходит суммирование квантов медиатора, и наконец его количество становится достаточным для возникновения возбуждающего постсинаптического потенциала, а затем и потенциала действия. Кроме суммации во времени, в нервных центрах возможна пространственная суммация. Она характеризуется тем, что если раздражать одно афферентное волокно раздражителем допороговой силы, то ответной реакции не будет, а если раздражать несколько афферентных волокон раздражителем той же допороговой силы, то возникает рефлекс, так как импульсы, приходящие с нескольких афферентных волокон, суммируются в нервном центре.
В нервном волокне каждое одиночное раздражение (если оно не подпороговой и не свехпороговой силы) вызывает один импульс возбуждения. В нервных же центрах, как показал впервые И.М.Сеченов, одиночный импульс в афферентных волокнах обычно не вызывает возбуждения, т.е. не передается на эфферентные нейроны. Чтобы вызвать рефлекс необходимо быстрое нанесение допороговых раздражений одно за другим. Это явление получило название временной или последовательной суммации. Ее сущность состоит в следующем. Квант медиатора, выбрасываемого окончанием аксона при нанесении одного допорогового раздражения, слишком мал для того, чтобы вызвать возбуждающий постсинаптический потенциал, достаточный для критической деполяризации мембраны. Если же к одному и тому же синапсу идут быстро следующие один за другим допороговые импульсы, происходит суммирование квантов медиатора, и наконец его количество становится достаточным для возникновения возбуждающего постсинаптического потенциала, а затем и потенциала действия. Кроме суммации во времени, в нервных центрах возможна пространственная суммация. Она характеризуется тем, что если раздражать одно афферентное волокно раздражителем допороговой силы, то ответной реакции не будет, а если раздражать несколько афферентных волокон раздражителем той же допороговой силы, то возникает рефлекс, так как импульсы, приходящие с нескольких афферентных волокон, суммируются в нервном центре.
Явление суммации возбуждения в ЦНС открыл И.М.Сеченов (1868) в опыте на лягушке: раздражение конечности лягушки слабыми редкими импульсами не вызывает реакции, а более частые раздражения такими же слабыми импульсами сопровождается ответной реакцией - лягушка совершает прыжок.
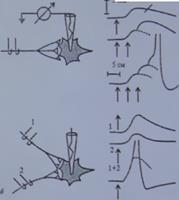
Различают временную (последовательную) суммацию и пространственную суммацию (рис. 4.6).
 Временная суммация. На рис. 4.6 слева показана схема для экспериментального тестирования эффектов, вызываемых в нейроне ритмической стимуляцией аксона. Запись вверху позволяет видеть, что если ВПСП быстро следуют друг за другом, то они суммируются благодаря своему относительно медленному временному ходу (несколько миллисекунд), достигая в конце концов порогового уровня. Временная суммация обусловлена тем, что ВПСП от предыдущего импульса еще продолжается, когда приходит следующий импульс. Поэтому данный вид суммации называют также последовательной суммацией. Она играет важную физиологическую роль, потому что многие нейронные процессы имеют ритмический характер и, таким образом, могут суммироваться, давая начало надпороговому возбуждению в нейронных объединениях нервных центров.
Временная суммация. На рис. 4.6 слева показана схема для экспериментального тестирования эффектов, вызываемых в нейроне ритмической стимуляцией аксона. Запись вверху позволяет видеть, что если ВПСП быстро следуют друг за другом, то они суммируются благодаря своему относительно медленному временному ходу (несколько миллисекунд), достигая в конце концов порогового уровня. Временная суммация обусловлена тем, что ВПСП от предыдущего импульса еще продолжается, когда приходит следующий импульс. Поэтому данный вид суммации называют также последовательной суммацией. Она играет важную физиологическую роль, потому что многие нейронные процессы имеют ритмический характер и, таким образом, могут суммироваться, давая начало надпороговому возбуждению в нейронных объединениях нервных центров.
Пространственная суммация (см. рис. 4.6, б). Раздельная стимуляция каждого из двух аксонов вызывает подпороговый ВПСП, тогда как при одновременной стимуляции обоих аксонов возникает ПД, что не может быть обеспечено одиночным ВПСП. Пространственная суммация связана с такой особенностью распространения возбуждения, как конвергенция.
22. Принцип доминанты в ЦНС (А.А.Ухтомский), её роль в рефлекторной деятельности.
Известный русский физиолог А. А. Ухтомский (1875-1942) сформулировал принцип доминанты, который является одним из основных принципов деятельности ЦНС;
Доминанта — это господствующие в ЦНС очаги возбуждения, которые изменяют и «подчиняют» себе в данный момент деятельность других центров.
Доминанта формируется под влиянием различных адекватных стимулов внешней и внутренней среды организма (истощение питательных веществ, накопление в крови половых гормонов и др.). Все эти стимулы повышают возбудимость соответствующих нервных центров (пищевого, полового и т.д.). За счет суммации с посторонними (индифферентными) стимулами разной модальности в данном центре формируется стойкое и длительное возбуждение, сопряженное с торможением других, связанных с ним, центров. Так возникают пищевая, половая, оборонная и другие доминанты, которые в значительной мере определяют осуществления именно данного рефлекса, поведение человека и животных в данный момент.
Можно любой очаг возбуждения, возникшего в ЦНС, рассматривать как доминантный? Какие характерные черты доминанты? Доминантный очаг возбуждения (любой центр) характеризуется следующими свойствами: 1) Повышение возбудимости, 2) устойчивость возбуждения, 3) способность к суммации возбуждения (накапливать возбуждения за счет сторонних раздражений) 4) способность тормозить другие нервные центры и системы рефлексов, 5 ) способность длительное время удерживать возбуждения по окончании раздражение, которое его обусловило (инерция доминанты).
"Примерами доминанты могут быть следующие наблюдения. Раздражение у животных некоторых участков коры большого мозга, которое обычно сопровождается сгибанием конечностей, во время акта дефекации или мочеиспускания такой реакции не вызывает. Наоборот, происходит усиление этих рефлекторных актов. Это обусловлено тем, что в данный момент центры дефекации или
мочеиспускания находятся в состоянии доминантного возбуждения, который «притягивает» к себе посторонние возбуждения и оказывает сочетанное торможение других нервных центров.
Доминантный состояние может возникать и под влиянием гормонов, которые повышают возбудимость соответствующих центров. Например, у лягушек-самцов в весенний период заметно усиливается половой (охватывающий) рефлекс под влиянием посторонних (для полового центра) раздражений (например, при щипки кавычки). В то же время возникает защитного изгибающего рефлекса этой конечности.
Явления, которые напоминают доминанту, приходится наблюдать и в клинической практике, когда источники возбуждения, возникшие в ЦНС, подобно магниту, «притягивают» к себе различные посторонние раздражения, усиливаясь за их счет. Так, при язвенной болезни желудка и двенадцатиперстной кишки в стадии доминантного возбуждения находится пищевой центр, при гипертонической болезни - сосудодвигательный, вследствие чего различные возбуждение (особенно эмоциональные) приводят к повышению желудочной секреции или уровня системного артериального давления. После травмы нервных стволов нередко в состоянии доминантного возбуждения находятся центры болевой чувствительности в таламокортикальиых структурах МОАК, вследствие чего усиливается боль в раненой конечности (каузалгия) при различных посторонних раздражениях (сильном зуде, притрагивании к любой части тела и т.п.).
Прекращаться доминанта может разными путями: I) вследствие устранения раздражения или удовлетворения потребностей (биологической, социальной), на основании которых сформировалась доминанта, 2) при возникновении в ЦНС сильной доминанты, тормозящей ту, которая является, 3) вследствие развития в доминантном центре вторичного торможения.
Благодаря доминанте в ЦИС создаются не стабильны, а динамические координационные отношения между нервными центрами и системами рефлексов. Доминанта играет важную роль в психической деятельности человека и животных, в значительной мере определяет поведенческие акты в определенный момент.
Открытие центрального торможения. Классический опыт И.М.Сеченова на таламической лягушке.
И. М. Сеченов (1862) открыл торможение в центральной нервной системе. Он показал, что при раздражении области зрительных чертогов лягушки происходит торможение моторных спинномозговых рефлексов, так как весьма значительно увеличивается их латентный период. Явление центрального торможения было подтверждено учениками И. М. Сеченова и на животных с постоянной температурой тела (Л. Н. Симонов, 1866). Головной мозг не только тормозит спинномозговые рефлексы, но при определенных условиях усиливает их (И. Г. Березин, 1866, В. В. Пашутин, 1866).
Значение открытия центрального торможения для дальнейшего развития физиологии
И. М. Сеченов впервые доказал влияние ретикулярной формации мозгового ствола на спинной мозг. Открытие И. М. Сеченова явилось отправным пунктом для работ школы И. П. Павлова по изучению закономерностей взаимоотношения возбуждения и торможения в головном мозге и работ школы Н. Е. Введенского по изучению природы торможения и единства возбуждения и торможения.
Во всех видах центрального торможения, вызываемого импульсами, поступающими по афферентным волокнам, и осуществляемого эфферентными импульсами по пирамидным путям, участвуют вставочные нейроны. Различают первичное торможение, вызванное активацией тормозных синапсов и возникающее без предварительного возбуждения, и вторичное торможение, как результат предшествовавшего возбуждения.
К первичному торможению относятся постсинаптическое, включающее возвратное торможение моторных нейронов клетками Реншоу, и пресинаптическое. К вторичному торможению относятся индукционное торможение после возбуждения при реципрокной иннервации и пессимальное торможение Н. Е. Введенского, не обнаруженное в центральной нервной системе в норме.
1. Постсинаптическое торможение, при котором возникают тормозные постсинаптические потенциалы (ТПСП) в тормозных синапсах 2-го типа. В спинном мозге ТПСП появляются в моторных нейронах и нейронах Реншоу при определенных условиях притока афферентных импульсов, в головном мозге — корзинчатых и других тормозных нейронах. В спинном мозге латентный период ТПСП 0,3 мс, они достигают максимума через 0,8 мс и продолжаются около 2.5 мс. В нейронах головного мозга они продолжаются значительно дольше, 100-200 мс. Частота разряда ТПСП до 1000 имп/с. Они также суммируются в пространстве и во времени, как и ВПСП ТПСП - почти зеркальное отражение ВПСП (ТПСП противодействует ВПСП, препятствует возникающей деполяризации, так как при ТПСП возникает гиперполяризация постсинаптической мембраны. Когда раздражение афферентного нерва, вызывающее торможение и появление ТПСП, предшествует ВПСП, то последний подавляется. При действии тормозного раздражителя во время проведения импульсов ВПСП они становятся реже или исчезают. Результат торможения зависит от соотношения амплитуд ВПСП и ТПСП и количества участвующих возбуждающих и тормозных синапсов.
Сеченов решил доказать существование в мозгу специальных центров, задерживающих отраженные (рефлекторные, вызванные раздражением) движения и определить их местонахождение. После долгих размышлений он наметил три главные серии исследований: перерезка мозга в различных точках; раздражение мозга физическими (при помощи электричества) или химическими агентами; возбуждение нервных центров физиологическими путями.
Все эксперименты он проводил на классическом объекте исследований физиологов того времени — лягушках. С ювелирной точностью производил разрезы мозга, а затем подолгу наблюдал, какие изменения происходили под влиянием каждого разреза в отражательных движениях животных. Полученные факты говорили сами за себя: угнетение отраженной деятельности возникает лишь после разрезов мозга непосредственно перед зрительными буграми и в них самих.
24. Виды торможения в ЦНС. Тормозные синапсы и их медиаторы.
Для обеспечения приспособления и адекватного поведения необходимы не только способность к выработке новых условных рефлексов и их длительное сохранение, но и возможность к устранению тех условно-рефлекторных реакций, необходимость в которых отсутствует. Исчезновение условных рефлексов обеспечивается процессами торможения. По И.П.Павлову, различают следующие формы коркового торможения: безусловное, условное и запредельное торможение.
Безусловное торможение
Этот вид торможения условных рефлексов возникает сразу в ответ на действие постороннего раздражителя, т.е. является врожденной, безусловной формой торможения. Безусловное торможение может быть внешним и запредельным. Внешнее торможение возникает под влиянием нового раздражителя, создающего доминантный очаг возбуждения, формирующего ориентировочный рефлекс. Биологическое значение внешнего торможения состоит в том, что, затормаживая текущую условно- рефлекторную деятельность, оно позволяет переключить организм на определение значимости и степени опасности нового воздействия.
Условное торможение (внутреннее)
Оно возникает, если условный раздражитель перестает подкрепляться безусловным. Его называют внутренним, потому что оно формируется в структурных компонентах условного рефлекса. Условное торможение требует для выработки определенного времени. К этому виду торможения относятся: угасательное, дифференцировочное, условный тормоз и запаздывающее.
условный тормоз — это разновидность дифференцировочного торможения. Возникает в том случае, если положительный условный раздражитель подкрепляется безусловным, а комбинация из условного и индифферентного раздражителей не подкрепляется. Например, условный раздражитель свет подкрепляется безусловным раздражителем, а комбинация свет и звонок не подкрепляется.
Первоначально эта комбинация вызывает такой же условный ответ, но в дальнейшем она утрачивает свое сигнальное значение и на нее условная реакция возникать не будет, в то время как на изолированный условный раздражитель (свет) она сохраняется. Звонок же приобретает значение тормозного сигнала. Его подключение к любому другому условному раздражителю затормаживает проявление условного рефлекса.
Запредельное торможение
Этот вид торможения отличается от внешнего и внутреннего по механизму возникновения и физиологическому значению. Оно возникает при чрезмерном увеличении силы или продолжительности действия условного раздражителя вследствие того, что сила раздражителя превышает
работоспособность корковых клеток. Это торможение имеет охранительное значение, так как препятствует истощению нервных клеток. По своему механизму оно напоминает явление "пессимума", которое было описано Н.Е.Введенским.
Запредельное торможение может вызываться действием не только очень сильного раздражителя, но и действием небольшого по силе, но длительного и однообразного по характеру раздражения. Это раздражение, постоянно действуя на одни и те же корковые элементы, приводит их к истощению, а следовательно, сопровождается возникновением охранительного торможения. Запредельное торможение легче развивается при снижении работоспособности, например, после тяжелого инфекционного заболевания, стресса, чаще развивается у пожилых людей.
Дата: 2019-03-05, просмотров: 468.