
| [17] |
| Типы (механизмы) проведения возбуждения по миокарду: 1. С участием специализированной проводящей системы сердца. 2. Передачей возбуждения от одного рабочего миокардиоцита к другому. |
| Скорость проведения возбуждения по миокарду: · предсердий составляет порядка 1 м/с · желудочков - 0,8 м/с · по проводящей системе - до 4-5 м/с |
Т.е. возбуждение по проводящей системе сердца в 5 раз быстрее скорости распространения возбуждения по рабочему миокарду.
Напомню, что проведением возбуждения по нервным волокнам составляет 0,5 – 120 м[Б47] ·с-1.
|
| [18] |
Во всех случаях проведение возбуждения осуществляется за счет наличия нексусов - специальных «щелевых контактов» [Б48] (рис[Б49] . 810132328).
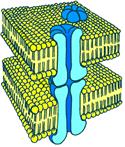
Рис[Б50] . 810132328. Нексус. Объяснение в тексте
Нексусы, как правило, соединяют группы синхронно функционирующих клеток, образующих функциональный синцитий.
Как устроены нексусы? В каждой из двух соседних клеточных мембран находятся регулярно распределенные коннексоны, пронизывающие всю толщу мембраны. Они расположены так, что в месте контакта клеток находятся друг против друга и их просветы оказываются на одной линии. У образованных таким образом каналов крупные внутренние диаметры и, значит, высокая проводимость для ионов. Через них могут приходить даже относительно крупные молекулы с молекулярной массой до 1000 (около 1,5 нм в поперечнике). Коннексон состоит из субъединиц числом до шести с молекулярной массой примерно 25000 каждая.
Но главное через нексусы свободно проходят ионы.
Принцип передачи возбуждения через нексус показан на рис[Б51] . 810132331.
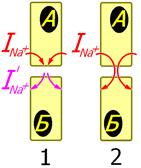
Рис[Б52] . 810132331. Натриевые токи (INa+) при передаче возбуждения в химическом синапсе (1), электрическом синапсе или между миокардиоцитами (2).
Принцип передачи возбуждения между миокардиоцитами аналогичен принципу передачи возбуждения в электрическом синапсе и существенно отличается от принципа передачи возбуждения в химическом синапсе.
При возбуждении клетки А натриевый ток (INa+) входит в нее через открытые потенциалуправляемые натриевые каналы. При этом часть тока входит через участок мембранного контакта в клетку Б, вызывая ее деполяризацию. Уровень деполяризации клетки Б гораздо ниже, однако он может оказаться выше критического уровня деполяризации мембраны клетки Б и в ней генерируется потенциал действия.
В химическом синапсе входящие натриевые токи клетки А прерываются на пресинаптической мембране. Деполяризующие входящие натриевые токи на постсинаптической мембране клетки Б возникают при активации медиаторами рецепторуправляемых натриевых каналов. При достижении критического уровня деполяризации мембраны клетки Б открываются потенциалуправляемые натриевые каналы.
Таким образом, деполяризующие токи в клетке Б в химическом синапсе образуются в самой клетке Б, а в электрическом синапсе и при контакте миокардиоцитов деполяризующие токи в клетку Б входят из клетки А.
Для [Б53] миокарда важна регулируемость щелевых контактов. Каналы миокардиоцитов закрываются при снижении pH или повышении концентрации Са2+. Это неизбежно происходит в случае повреждения клеток или глубокою нарушения обмена. За счет такого механизма пораженные места изолируются от остальной части функционального синцития, и распространение патологии ограничивается (например, при инфаркте миокарда).
В миокарде, как функциональном синцитии, возбуждение одной точки миокарда неизбежно вызывает возбуждение всех остальных областей миокарда. Это означает, что сердечная мышца отвечает на возбуждение по правилу "все или ничего", а градуальная зависимость, типичная для скелетной мышцы, здесь при нормальных условиях не наблюдается[Б54] .
Благодаря проводящей системе сердца волна возбуждения синхронно возбуждает рабочие миоциты.
Однако в атриовентрикулярном узле волна возбуждения на участке длиной 1,5‑2 мм задерживается, бежит с малой скоростью (2-5 см/с). Тем самым обеспечивается так называемая атриовентрикулярная задержка, благодаря которой возбуждение желудочков и их сокращение начинается через 100 мс после начала сокращения предсердий, а не раньше. Эта зона расположена в верхней части атриовентрикулярного узла.
Полагают, что снижение скорости проведения в этой зоне обусловлено особенностями контакта миоцитов - ход волокон перпендикулярен направлению волны возбуждения, что и обусловливает замедление ее движения по этому участку.
Важно отметить, что проведение волны возбуждения через атриовентрикулярный узел осуществляется лишь в том случае, если одновременно возбуждаются несколько миоцитов этого узла. Это защитный механизм от возникновения аритмий и появление эктопических очагов возбуждения.
Как правило, вначале возбуждается правое предсердие, а затем - с небольшим интервалом - левое предсердие.
Проводящая система сердца
|
| [19] |
В наиболее развитом виде проводящая система представлена в сердце млекопитающих и человека (рис. 810140007).
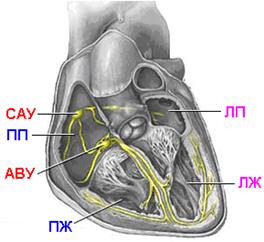 Рис. 810140007. Проводящая система сердца. САУ – синоатриальный узел, ПП – правое предсердие, ЛП – левое предсердие, АВУ – атриовентрикулярный узел, ПЖ – правый желудочек, ЛЖ – левый желудочек. Объяснение в тексте.
Рис. 810140007. Проводящая система сердца. САУ – синоатриальный узел, ПП – правое предсердие, ЛП – левое предсердие, АВУ – атриовентрикулярный узел, ПЖ – правый желудочек, ЛЖ – левый желудочек. Объяснение в тексте.
|
Ткани проводящей системы обладают рядом особенностей по сравнению с рабочим миокардом. Они могут использовать энергию как аэробного, так и анаэробного гликолиза. Последнее свойство создает повышенную устойчивость проводящей системы к гипоксии.
Другой особенностью проводящей системы является ее устойчивость к повышенной концентрации внеклеточного K+. Это обеспечивает проведение возбуждения в ней в таких условиях, когда кардиомиоциты оказываются невозбудимыми.
|
По функциональным особенностям, по специфике морфологии и ультраструктуры проводящую систему высших позвоночных и человека делят на следующие отделы [Б55] [Б56] :
1. Узловую ткань 1.1. Синусовый узел (Киса-Фляка) 1.2. атриовентрикулярный узел (Ашоффа-Тавара); 2. Собственно проводящие пути 2.1. Проводящие пути предсердий 1.2.1. Межпредсердный путь Бахмана 1.2.2. Межузловые проводящие пути предсердий 2.1.2.1. Бахмана 2.1.2.2. Венкебаха 2.1.2.3. Тореля 1.2.3. Пучок Гиса, 1.2.4. Ножки пучка Гиса 2.1.4.1. Правая 2.1.4.2. Левая 2.1.4.2.1. Передняя ветвь 2.1.4.2.2. Задняя ветвь 1.2.5. Субэндокардиальная сеть волокон Пуркинье.
|
Синоатриальный узел
|
| [21] |
Синонимы – синусовый узел, синусный узел, узел Киса-Фляка.
С[Б57] иноатриальный узел расположен в верхней части правого предсердия в устье верхней полой вены. Длина узла составляет 1,5 — 2,5 см, ширина — 0,4 — 0,7 см. Различают головку синусового узла, расположенную субэпикардиально, и хвост, располагающийся субэндокарднально.
В синусовом узеле выделяют четыре морфологических типа клеток: пейсмекеры (Р-клетки[Б58] ), недифференцированные, промежуточные, волокна Пуркинье.
Пейсмекеры расположенные главным образом в центре узла. Недифференцированные клетки миокарда находятся преимущественно в периферических отделах узла. Волокна Пуркинье располагаются по краям синусового узла и подходят к предсердным проводящим путям. Промежуточные клетки обладающие морфологическими признаками пейсмекеров и недифференцированных клеток.
Кровоснабжение синусового узла осуществляет ramus cristae terminalis, артерии, отходящей от правой (иногда от левой) венечной артерии.
Проводящие пути предсердий
|
| [22] |
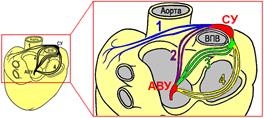 Рис[Б59] . 810071559 [Б60] . Проводящие пути предсердий (вид сверху и сзади).
1 – межпредсердный путь Бахмана,
2 – передний межузловой путь Бахмана,
3 – средний межузловой путь Венкебаха,
4 – задний межузловой путь Тореля.
СУ – синоатриальный узел,
АВУ – атриовентрикулярный узел.
ВПВ – верхняя полая вена.
Рис[Б59] . 810071559 [Б60] . Проводящие пути предсердий (вид сверху и сзади).
1 – межпредсердный путь Бахмана,
2 – передний межузловой путь Бахмана,
3 – средний межузловой путь Венкебаха,
4 – задний межузловой путь Тореля.
СУ – синоатриальный узел,
АВУ – атриовентрикулярный узел.
ВПВ – верхняя полая вена.
|
Передний тракт исходит из переднего края синусового узла, огибает спереди верхнюю полую вену и делится на две части. Одна из них идет к левому предсердию, образуя межпредсердный пучок Бахмана (Bachman), другая часть возвращается к межпредсердной перегородке и достигает гребня атриовентрикулярного узла.
Средний межузловой трактВенкебаха (Wenckebach) исходит из задней части синусового узла, огибает сзади верхнюю полую вену и, спускаясь по правой стороне межпредсердной перегородки, подходит к гребню атриовентрикулярного узла.
Задний предсердный тракт Тореля (Thorel) отходит от задней части синусового узла, следуя вдоль crista terminalis, обходит основную часть атриовентрикулярного узла и подходит к его нижней части. Некоторая часть волокон заднего тракта принимает участие в формировании пучка Гиса.
В физиологических условиях импульсы, проходящие по более коротким переднему и среднему трактам, быстрее достигают атриовентрикулярного узла. Межпредсердный тракт обеспечивает практически синхронную работу правого и левого предсердий.
Атриовентрикулярный узел
|
| [23] |
Синонимы – атриовентрикуляное соединение, узел Ашоффа-Тавара. В последние годы вместо термина «атриовентрикулярный узел» часто употребляют более широкое понятие — «атриовентрикулярное соединение». Этим термином обозначают анатомическую область, включающую в себя атриовентрикулярный узел и специализированные клетки предсердий, лежащие в области узла.
Атриовентрикулярный узел расположен в нижней части межпредсердной перегородки справа, кпереди от коронарного синуса[Б61] .
Атриовентрикулярный [Б62] узел часто разделяют на три функциональные зоны (этажа): AN (atrium-nodus), N (nodus), NH (nodus-His) (рис. 810140040).
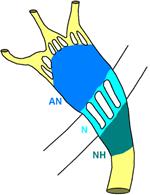 Рис[Б63] . 810140040. Атриовентрикулярный узел. Объяснения в тексте.
Рис[Б63] . 810140040. Атриовентрикулярный узел. Объяснения в тексте.
|
Выделяют четыре типа клеток атриовентрикулярного узла, аналогичных клеткам синусового узла: пейсмекеры, переходные и сократительные клетки, клетки Пуркинье.
Пейсмекеров немного и он располагаются главным образом в области перехода атриовентрикулярного узла в пучок Гиса. Переходные клетки составляют основную массу атриовентрикулярного узла. Клетки сократительного миокарда, располагающиеся главным образом у атрионодального края.
Известны два участка, в которых задерживается проведение импульса: первый — в области предсердного края атриовентрикулярного узла, второй — в месте соединения узла с пучком Гиса. Предпосылкой для возникновения этого феномена является существование двух или нескольких каналов проведения — импульсов в атриовентрикулярном узле (рис. 810140040). [Б64]
Дата: 2019-02-02, просмотров: 440.