В ротовой полости пищевой комок лишь измельчается. В желудке молодняка имеется липаза расщепляющая ацилглицеролы при РН 5,5. У взрослы животных рН ниже и активность этого фермента тормозится. Липиды поступают в 12-персную кишку, содержимое которого приобретает слабо щелочную реакцию. Здесь они расщепляются липазой, поступившей из поджелудочной железы.
СН2 -О-СО-С17Н35 СН2 -ОН
│ │
С17Н35 -СО-О-СН Липаза НО-СН + 3 С17Н35 - СООН
│ │
СН2-О-СО-С17Н35 + Н2О СН2-ОН Стеариновая кислота
Тристеарин Глицерол
Этому способствуют желчные кислоты, образующиеся в печени из холестерола и образующие комплексные соединения с ацилглицеролами. Секрецию желчных кислот из желчного пузыря усиливает холецистокинин, вырабатываемый стенкой кишечника. Желчные кислоты эмульгируют жиры, увеличивая площадь соприкосновения их с ферментом, активируют липазу. Отщепившиеся от ацилглицеролов жирные кислоты не растворимы в воде и поэтому образуют растворимые комплексы с желчными кислотами (холеиновые кислоты). В стенке кишечника они расщепляются и жирные кислоты всасываются. Фосфоглицериды, поступившие с пищей, в 12-персной кишке расщепляются фосфолипазами А1, А2, С и D, а эфиры холестерола и жирных кислот – холестеразой. В стенке кишечника из глицерола и жирных кислот синтезируются эндогенные ацилглицеролы. Они соединяются с холестерином, окружаются слоями из фосфоглицеридов и белков, образуя транспортные липопротеины, и поступают в кровь.
Транспорт липидов кровью.
Различают 4 разновидности транспортных липопротеинов, поступающих в кровь из стенки тонкого кишечника и печени: хиломикроны, пре-β-, β- и α-липопротеины. Наиболее высокое соотношение между уровнем в крови липидов и белков отмечается у хиломикронов, наиболее низкое – у α-липопротеинов (липоротеинов высокой плотности. У пре-β- β-липопротеинов (липоротеинов низкой плотности) оно занимает промежуточное положение. Содержание последних в сыворотке крови увеличивается при атеросклерозе. Транспортные липопротеины током крови доставляются в печень, глее расщепляются липопротеинлипазой. Освободившиеся глицерол и жирные кислоты окисляются.
Окисление глицерола в клетках.
Начинается оно с активирования этого вещества:
З-фосфоглицериновый альдегид подвергается дальнейшему окислению с участием ферментов, катализирующих реакции второго этапа гликолиза. При аэробном окислении одной молекулы глицерола генерируется 23 молекулы АТФ:
3-глицеролфосфатдегидрогеназная реакция – 3 АТФ,
3-фосфоглицеральдегиддегидрогеназная реакция – 6 АТФ,
фосфоглицераткиназная реакция – 2 АТФ,
пируваткиназная реакция – 2 АТФ,
пируватдегидрогеназная реакция – 6 АТФ,
окисление ацетил-КоА в цикле Кребса – 12 АТФ.
Поскольку 1 молекула АТФ затрачивается в глицеролкиназной реакции, то итоговый энергетический эффект всего процесса – 22 молекулы АТФ.
β-кисление жирных кислот в клетках
С наибольшей интенсивностью этот процесс протекает в печени, где жирные кислоты окисляются до ацетил-КоА. В сердце и других органах они способны окисляться до углекислого газа и воды. Протекает β-окисление жирных кислот в пространстве между внутренней и наружной мембранами митохондрий. Начинается оно с активирования молекулы жирной кислоты:

Так завершается один цикл β-окисления жирной кислоты, во время которого генерируется по одной молекуле ФАД-Н2 и НАД-Н2, окисляемых в дыхательной цепи митохондрий, и ацетил-КоА, способного окисляться в цикле Кребса.
Укоротившийся на 2 углеродных фрагмента активированный остаток пальмитиновой кислоты (ацил-КоА) таким же образом, цикл за циклом, подвергается дальнейшему β-окислению. На последнем этапе этого процесса образуется активная форма масляной кислоты (бутирил-КоА). Она также подвергается β-окислениию:
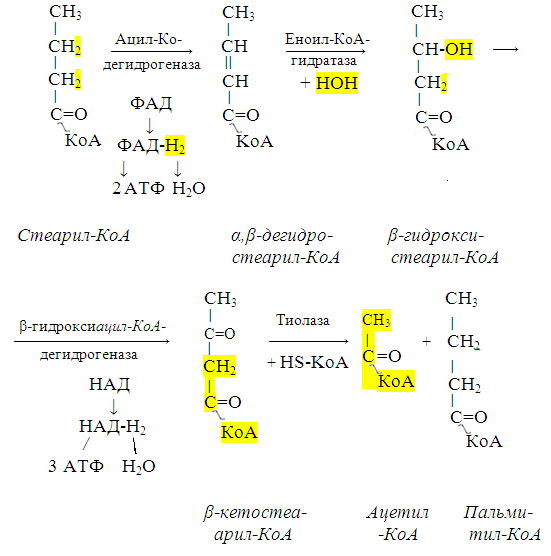
Из этого четырехуглеродной активированной жирной кислоты генерируется 2 молекулы ацетил- КоА, но не по две, а по одной молекуле ФАД-Н2 и НАД-Н2 (образовавшаяся молекула ацетил- КоА не подвергается дальнейшему β-окислению).
Энергетический эффект β-окисления жирных кислот (Х) рассчитывается по формуле: Х = n/2 X 17 – (5-1), где n – число углеродных атомов, включая группу –СООН; 17 – число молекул АТФ, генерируемых во время одного цикла: при окислении ФАД-Н2 (2 АТФ),НАД-Н2 (3 АТФ) и ацетил-КоА в цикле Кребса.(12 АТФ); 5 – количество АТФ, не образовашегося на последнем этапе β-окисления; 1 – количество АТФ, затрачиваемое на активацию жирной кислоты (образование ацил-КоА).
β-окисления жирных кислот в печени.
В этом органе основная масса жирных кислот окисляется до ацетил-КоА. Две молекулы последнего последнего соединяются между собой в результате реакции, катализируемой тиолазой. Образовавшаяся ацетоуксусная кислота током крови доставляется в мышечную и другие ткани, где обратно расщепляется до ацетил-КоА, который окисляется в цикле Кребса. Часть молекул ацетоуксусной кислоты, восстанавливаясь ионами водорода, отщепляемыми от НАД-Н2, превращается в β-гидрооксимасляную килоту. Какое-то количество молекул ацетоуксусной кислоты, отщепляя от карбоксильной группы углекислый газ, превращается в ацетон. Общее количество ацетоуксусной, β-гидрооксимасляной кислот и ацетона называют кетоновыми телами. Количество последних в тканях увеличивается (кетоз) при усиленном образовании их и недостаточно эффективном окислении.
Окисление пропионовой кислоты, образовавшейся в рубце при расщеплении микроорганизмами клетчатки, происходит после превращения ее в сукцинил-КоА (см. обмен углеводов). Последний в дальнейшем окисляется в цикле Кребса.
Лекция №12
ОБМЕН ЛИПИДОВ (продолжение)
План
1. Биосинтез глицерола и жирных кислот.
2. Биосинтез ацилглицеролов, фосфоглицеридов и холестерина.
3. Патология липидного обмена.
Биосинтез жирных кислот (путь Linean)
Этот процесс протекает в цитоплазме клеток. Суть его – постепенное удлинение молекулы жирной кислоты на 2 углеродных атома за счет молекулы ацетил-КоА и четырех ионов водорода, поставляемых НАДФ-Н2. на первом этапе происходит присоединение к ацетил-КоА углекислого газа с участием биотин-фермента.
рибозо-5-фосфат и НАДФ-Н2, генерируемые из глюкозы в реакциях пентозного цикла, а также кобаламин и фолиевая кислота, вырабатываемые микрофлорой рубца.
рибозо-5-фосфат и НАДФ-Н2, генерируемые из глюкозы в реакциях пентозного цикла, а также кобаламин и фолиевая кислота, вырабатываемые микрофлорой рубца.
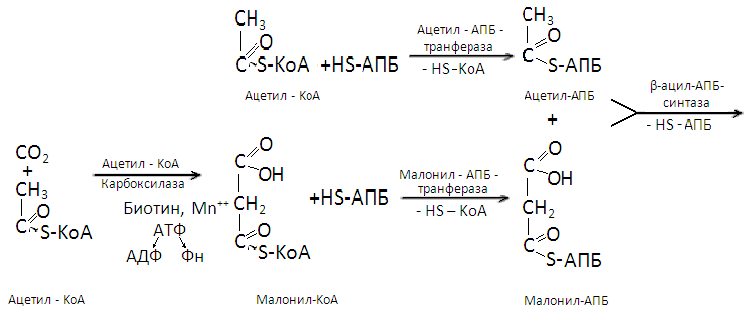
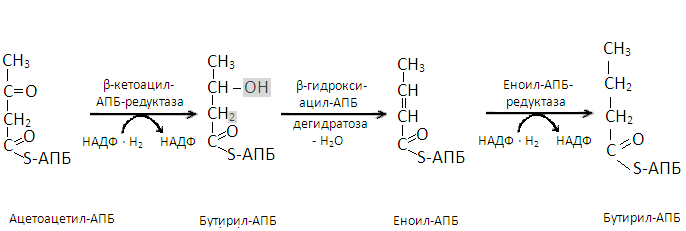
В образовавшемся малонил-КоА радикал –S-КоА заменяется на радикал ацилпереносящего белка (-АПБ). Параллельно радикал –S-КоА заменяется на радикал -АПБ в молекуле ацетил-КоА. Затем малонил-АПБ, отщепляя углекислый газ, соединяется с ацетил-АПБ и образуется ацетоацетил-АПБ. Последний восстанавливается ионами водорода, отщепляемыми от НАДФ-Н2, в β-гидроксибутирил-АПБ. После отщепления от этого вещества молекулы воды образуется кротонил-АПБ, восстанавливающийся в дальнейшем ионами водорода НАДФ-Н2 до бутирил-АПБ. Этой реакцией завершается первый цикл биосинтеза жирной кислоты.
Второй цикл начинается с соединения β-гидроксибутирил-АПБ с ацетил-АПБ с отщеплением молекулы углекислого газа. Образовавшийся капронил-АПБ таким же образом, как и ацетоацетил-АПБ, цикл за циклом, превращается в ацил-АПБ синтезируемой жирной кислоты с последующим отщеплением от последнегорадикала АПБ.
Количество молекул АТФ, затрачиваемое на биосинтез одной молекулы жирной кислоты (ХАТФ) рассчитывают про формуле: ХАТФ = n/2 – 1, где где n – число углеродных атомов, включая группу –СООН, а количество НАДФ-Н2 (ХНАДФ-Н) – по формуле: ХНАДФ-Н = n – 2.
Биосинтез глицерол-3-фосфата происходит из 3-фосфоглицеральдегида, образовавшегося из глюкозы в реакциях гликолиза:
Биосинтез ацилглцеролов.
На первом этапе его глицерол-3-фосфат взаимодействует с двумя активированными молекулами жирных кислот (например, стеарил-КоА)с образованием фосфатидной кислоты. От последней отщепляется остаток фосфорной кислоты с последующим присоединением на его место третьего радикала жирной кислоты.
СН2 –ОН СН2 –О-СО- С17Н35
1 1
НО-СН + 2 Стеарил-КоА Ацил-КоА- С17Н35–СО-О-СН Фосфатидиофосфатаза
1 трансфераза 1 + НОН
СН2-О-РО3Н2 СН2-О-РО3Н2 -Н3РО4
3-фосфоглицерол Фосфатидная кислота
СН2 -О-СО-С17Н35 СН2 -О-СО-С17Н35
1 1
С17Н35 –CO-О-СН + Стеарил-КоА Ацил-КоА- С17Н35 –CO-О-СН
1 трансфераза 1
СН2-ОН СН2 -О-СО-С17Н35
Дистеарин Тристеарин
Биосинтез фосфоглицеридов.
Вначале, как описано выше, синтезируется фосфатидная кислота, в которой к второму углеродному атому глицерола присоединен остаток арахидоновой кислоты.После этого активируется молекула холина:
НО-СН2 -СН2–N+ ≡(CH3)3 + АТФ Холинкиназа Н2О3Р-О-СН2 -СН2–N+≡(CH3)3
Холин -АДФ Фосфохолин
Фосфохолин взаимодействует с цитидинтрифосфатом (ЦТФ):
Н2О3Р-О-СН2 -СН2–N+≡(CH3)3 + ЦТФ Трансфераза Цитидилдифосфохолин
-Н4Р2О1
СН2 –О-СО- С17Н35
1
С17Н35–СО-О-СН + Цитидилдифосфохолин Трансфераза
1
СН2-О-РО3Н2
Фосфатидная кислота
СН2 –О-СО- С17Н35
1
С17Н35–СО-О-СН
1
СН2-О-РО3Н -О-СН2 -СН2–N+ ≡(CH3)3
Фосфатидилхолин.
Биосинтез холестерола.
На первом этапе этого процесса из двух молекул ацетил-КоА в результате реакции, катализируемой тиолазой, образуется ацетоацетил-КоА. После присоединения к последнему третьей молекулы ацетил-КоА, катализиремого β-гидрокси-β-метил-глутатил-КоА-синтазой, образуется β-гидрокси-β-метил-глутатил-КоА. (ГМГ-КоА). Этот фермент способны ингибировать некоторые лекарственные средства, применяющиеся для лечения больных атеросклерозом, в частности зокор. ГМГ-КоА в дальнейшем восстанавливается ионами водорода, отщепляемыми от НАДФ-Н2 в результате реакции, катализируемой ГМФ-КоА-редуктазой, до мевалоната. Последний в результате шести последовательно протекающих реакций превращается в сквален, углеводород с длинной открытой цепью:
Березов- 312 стр.
В дальнейшем сквален в результате реакции, катализируемой скваленоксидоциклазой, циклизируется в ланостерин. Последний через несколько последовательных реакций превращается в холестерин.
Дата: 2019-02-02, просмотров: 406.