| Частота стимуляции | 3 | 4 | 6 | 9 | 11 | 14 | 18 | 22 | 25 | 28 |
| 3 | 300 | 285 | -065 | –075 | –272 | –319 | –216 | 136 | 257 | |
| 4 | 647** | 302 | 385* | 288 | 275 | 186 | 319 | 335 | ||
| 6 | 339 | 235 | –030 | 085 | 065 | 221 | 243 | |||
| 9 | 667** | –020 | –029 | 152 | 128 | 150 | ||||
| 11 | 159 | 161 | 102 | 208 | 131 | |||||
| 14 | 951*** | 886*** | 739*** | 655** | ||||||
| 18 | 925*** | 795*** | ||||||||
| 22 | 848*** | 796*** | ||||||||
| 25 | 956*** | |||||||||
| 28 |
Примечание. • р < 0,1; ** р < 0,01; ***р < 0,001.
Показатели реакции навязывания ритма, взятые суммарно, по частотным полосам, сохраняют ту же самую тенденцию, что и показатели реакции навязывания по отдельным частотам. Очень высоко коррелируют показатели этой реакции в частотных полосах β2 (21 – 30 кол/с) и β1 (r = 0,792; р < 0,001). Что касается индексов навязывания в частотных полосах дельта-, тета- и альфа-ритмов, то они на достаточном статистическом уровне значимости между собой не коррелируют.
Таким образом, выявляется тенденция навязанных ритмов к распределению по двум группам, из которых одна соответствует низким, а другая – высоким частотам световой стимуляции. Отсюда следует, что реакция навязывания ритма у детей при исследованных частотах стимуляции определяется, по крайней мере, двумя основными и, видимо, независимыми друг от друга факторами мозговой деятельности.
Этот вывод весьма близок к тем заключениям, к которым мы пришли, анализируя наши и Э.А. Голубевой данные относительно факторов, проявляющихся в реакции навязывания ритма у взрослых индивидов. Напомним, что, согласно этим заключениям, в реакциях навязывания ритма на разные частоты световой стимуляции отмечается действие нескольких факторов, достаточно независимых один от другого.
Единственное различие, которое отмечается между нашими результатами, полученными на взрослых и на детях, относится к корреляциям индексов навязывания в частотной полосе альфа-ритма. У детей индексы навязывания на частоты, входящие в зону альфа, лучше коррелируют с показателями навязывания на частоты тета-ритма и дельта-ритма. У взрослых же показатели реакции навязывания на частоты альфа-ритма занимают более определенное промежуточное положение между показателями навязывания на низкие и высокие частоты световой стимуляции. Но это различие, возможно, имеет чисто методические причины. Его можно, пожалуй, объяснить тем, что в исследованиях на взрослых использовался более широкий диапазон частот, входящих в зону альфа-ритма (4 частоты), а на детях – только 2. При наличии более широкого ряда частот внутренние корреляции для альфа-полосы, возможно, выступили бы более отчетливо.
Что же касается реакции навязывания ритма у детей как функции интенсивности стимула, то на ее характеристике в целом мы остановимся далее, в гл. 9. Укажем только, что зависимость, полученная на детях, оказалась весьма близка к той, которая была установлена для взрослых: в обоих случаях ее графическое изображение представляло собой асимптотическую функцию.
Рассмотрим теперь результаты сопоставления только что изложенных ЭЭГ характеристик с референтным индикатором динамичности возбуждения – средней длительностью условной реакции активации. Но сначала остановимся кратко на особенностях самого процесса образования условных электрокорковых реакций у детей – ведь задача получения референтного показателя динамичности возбуждения у детей была в излагаемой работе исходной и основной. В общем, оказалось, что эти особенности достаточно схожи с теми, которые наблюдаются у взрослых.
В случае простого, или нейтрального, подкрепления (свет на белом экране) в динамике образования условных связей отмечаются весьма значительные индивидуальные различия. У некоторых испытуемых условнорефлекторная блокада вырабатывается очень быстро и требует только нескольких сочетаний; у других, наоборот, условная депрессия альфа-ритма формируется очень медленно и требует большого числа сочетаний. У этих испытуемых подавление альфа-ритма на изолированное действие звука носит неустойчивый характера: в некоторых пробах оно имеется, в других – отсутствует. Увеличение числа сочетаний в некоторых случаях может привести к выработке и упрочению условной связи. В других случаях увеличение числа сочетаний, наоборот, приводит к полному и окончательному угашению условной связи. Наконец, в отдельных случаях (таких было три) условные связи не выработались совсем.
В опытах же с активирующим подкреплением выработка условно-рефлекторной активации у большинства испытуемых протекала очень быстро, требуя всего нескольких сочетаний, после которых она оставалась устойчивой до конца опыта. (Исключением из этого правила явились лишь трое испытуемых, у которых выработка условнорефлекторной депрессии с активирующим подкреплением протекала медленно и была получена только к концу принятой серии сочетаний. В опытах с простым подкреплением у двоих из этих испытуемых выработка условной депрессии вообще не удалась, а у третьего происходила с большим трудом). Поскольку, однако, активирующее подкрепление является в некотором смысле искусственным приемом, в значительной мере сглаживающим индивидуальные особенности процесса обусловливания, хотя и необходимым для исследования таких явлений, как угашение с подкреплением или выработка дифференцировки, в качестве референтного индикатора динамичности возбуждения были приняты, как и у взрослых, показатели выработки условных связей при простом, нейтральном подкреплении. При этом индексом испытуемого служило не число сочетаний до достижения критерия выработки, а средний условный эффект из всех 7 – 8 проб условного раздражителя, предъявлявшихся данному испытуемому. Его значения колебались от 0,5 до 4,7 с, составляя в среднем несколько более 2 с. Эти значения и были сопоставлены с индивидуальными величинами прочих электроэнцефалографических характеристик.
Один из способов установления возможных зависимостей между референтным и остальными индикаторами состоит в статистическом сравнении среднегрупповых значений сопоставляемых показателей для групп, сформированных на основе референтного индикатора. Испытуемые дети по динамичности возбуждения могли быть разбиты на две группы.
Первая группа (с низкой динамичностью возбуждения) включала тех испытуемых, у которых индивидуальный индекс по данному показателю равен иди ниже 2,1 с. В эту группу попали те испытуемые (9 человек), у которых условнорефлекторная блокада альфа-ритма либо совсем не вырабатывалась, либо вырабатывалась после большого числа сочетаний.
Вторую группу (с высокой динамичностью возбуждения) составили те испытуемые, индивидуальный индекс которых по взятому показателю больше 2,1 с (11 человек); у этих испытуемых условнорефлекторная депрессия альфа-ритма выработалась быстрее и была более устойчивой.
В табл. 18 представлены соответствующие средние, их разности, значения критериев различия и оценки уровня их статистической значимости.
Таблица 18
Статистическая оценка различий между средними арифметическими ряда ЭЭГ показателей для групп детей, различающихся по уровню динамичности возбудительного процесса (К. Войку, 1964)
| Статистические показатели | Альфа-индекс | Частота, кол/с | Суммарная энергия ритмов при записи ЭЭГ покоя, усл. ед. | Суммарная энергия ритмов при действии света, уел ед. | ||||||||||||
| β2 | α | θ | Δ | |||||||||||||
| β2 | β1 | α | θ | Δ | β2 | β1 | α | θ | Δ | |||||||
| Средние для группы с низкой динамичностью возбуждения (9 испытуемых) | 88,5 | 16,4 | 9,4 | 7,0 | 2,7 | 56,2 | 43,1 | 181,0 | 48,6 | 54,2 | 50,6 | 33,7 | 124,9 | 47,2 | 59,9 | |
| Средние для группы с высокой динамичностью возбуждения (11 испытуемых) | 80,0 | 15,9 | 9,7 | 6,3 | 2,7 | 72,0 | 57,9 | 156,5 | 48,5 | 54,0 | 50,0 | 36,5 | 110,2 | 46,6 | 53,5 | |
| Разность средних | 8,5 | 0,5 | 0,3 | 0,7 | 0 | 15,8 | 14,8 | 24,5 | 0,1 | 0,2 | 0,6 | 2,8 | 14,7 | 0,6 | 6,4 | |
| Критерий t | 3,05 | 1,86 | 1,0 | 2,38 | – | 2,14 | 3,20 | 3,19 | ||||||||
| Вероятность нулевой гипотезы р | <0,01 | >0,05 | >0,05 | <0,05 | – | <0,05 | <0,01 | <0,01 | ||||||||
Продолжение
| Статистические показатели | Длительность блокады альфа-ритма при 1‑м предъявлении звука, с | Число предъявлений звука до угашения ориентировки | Длительность блокады при 1‑м предъявлении света, с | Средняя длительность блокады из 7 – 8 предъявлений света, с | Средняя длительность условнорефлекторной блокады при простом подкреплении, с | Средняя длительность блокады при активирующем подкреплении, с |
| Средние для группы с низкой динамичностью возбуждения (9 испытуемых) | 2,2 | 5,0 | 3,9 | 3,6 | 1,4 | 1,6 |
| Средние для группы с высокой динамичностью возбуждения (11 испытуемых) | 2,9 | 7,1 | 4,5 | 3,6 | 3,2 | 2,6 |
| Разность средних | 0,7 | 2,1 | 0,6 | 0 | 1,8 | 1,0 |
| Критерий / | 2,23 | 3,11 | – | – | 6,15 | 3,0 |
| Вероятность нулевой гипотезы р | <0,05 | <0,01 | 8 | <0,001 | <0,01 |
Из фоновых показателей ЭЭГ статистически значимые различия при сравнении средних и, следовательно, зависимость от динамичности возбуждения обнаружили следующие показатели.
1. Альфа-индекс; разность его средних арифметических статистически значима на уровне р < 0,01. Типологическое значение альфа-индекса, как это явствует из настоящего сравнения, одинаково с тем, которое было установлено ранее нами в исследованиях на взрослых. Дети с более высокой динамичностью возбуждения характеризуются более низким, а с более низкой динамичностью этого процесса – более высоким альфа-индексом.
2. Частота тета-ритма; разность средних статистически значима на уровне р < 0,05. Дети с высокодинамичным возбуждением имеют меньшую частоту тета-ритма по сравнению с детьми с низкой динамичностью возбуждения. Этот факт, между прочим, хорошо согласуется с приведенными выше наблюдениями И.В. Равич-Щербо и М.К. Трифоновой, согласно которым уменьшение с возрастом альфа-индекса (которое можно интерпретировать как следствие повышения динамичности возбуждения) сопровождается уменьшением частоты тета-ритма. Что касается частоты других биоэлектрических составляющих, то соответствующие критерии различия не достигают 5 %-ного уровня значимости, однако заметим, что частота альфа-ритма в среднем все же выше у второй группы (с высокодинамичным возбуждением), а частота бета-ритма (в зоне низкочастотного бета-ритма) – больше у первой группы детей, в то время как частота дельта-ритма одинакова у обеих групп испытуемых.
3. Суммарная энергия частотных полос β2 и β1 (соответственно р < 0,05 и <0,01). У детей с пониженной динамичностью возбуждения суммарная биоэлектрическая энергия бета-ритма (во всяком случае, низкочастотного, до 30 кол/с) в среднем меньше, чем у детей с высокой динамичностью возбуждения, и наоборот.
4. Суммарная энергия частотной полосы альфа-ритма; разность средних значима на уровне р < 0,01. Значение этого показателя в аспекте динамичности возбуждения одинаково со значением альфа-индекса и противоположно значениям суммарных энергий обеих полос бета-активности: у детей с высокой динамичностью возбуждения количество биоэлектрической энергии в альфа-полосе в состоянии покоя меньше, чем, у детей с высокодинамичным возбуждением.
Остальные фоновые показатели ЭЭГ статистически значимой зависимости от динамичности возбуждения не обнаруживают, хотя для некоторых показателей, может быть, и намечаются определенные тенденции к связи (табл. 18).
5. Из показателей ориентировочной деятельности на звук и на свет в статистически значимой зависимости от динамичности возбуждения оказались только индикаторы ориентировки на звук. Разность средних величин скорости угашения этой реакции статистически значима на уровне р < 0,01. У детей с высокой динамичностью возбудительного процесса угашение ориентировки на звук требует больше предъявлений стимула, чем у детей противоположной группы.
Другой показатель ориентировки на звук – длительность блокады альфа-ритма на его первое предъявление – также обнаруживает некоторую зависимость от динамичности возбуждения. Его значения выше у детей второй группы (с высокой динамичностью возбуждения), а разность средних значима на уровне р < 0,05.
6. Из показателей, относящихся к явлению навязывания ритма, ни один не оказался в статистически значимой зависимости от динамичности возбудительного процесса. Это, видимо, объясняется прежде всего тем, что дисперсия индивидуальных индексов навязывания вокруг среднего арифметического является очень большой. Тем не менее здесь все же могут быть выявлены некоторые тенденции.
Так, анализ средних кривых навязывания ритма как функции частоты стимуляции (рис. 25) показывает, что если в полосе самых низких частот результаты по обеим группам приблизительно одинаковы и равны примерно нулю, то при частотах, соответствующих альфа-полосе, навязывание в группе с высокодинамичным возбуждением заметно хуже, а при частотах бета-полосы, особенно при самых высоких из них, напротив, несколько лучше, чем в противоположной группе детей – с низкой динамичностью возбуждения.
Правда, говорить о лучшем или худшем навязывании этих более высоких частот, видимо, не имеет смысла, поскольку, как это видно из графиков (рис. 25), навязывание частот 25 и 28 имп/с даже у «лучшей» группы колеблется около нуля, а у другой группы индексы этой реакции находятся в отрицательной зоне графика: имеет место тенденция группы в целом к «отрицательному» навязыванию, о котором мы говорили в предшествующей главе. Это означает, что у детей с низкой динамичностью возбуждения прерывистая стимуляция с указанными частотами световой импульсации ведет к большему подавлению фоновой ритмики бета-полосы, «разбивает» ее в большей степени, чем непрерывное световое раздражение, в то время как у детей с высощдинамичным возбуждением эффект прерывистой стимуляции, если не превышает (в среднем) эффекта непрерывного света, то, по крайней мере, хотя бы равен ему.
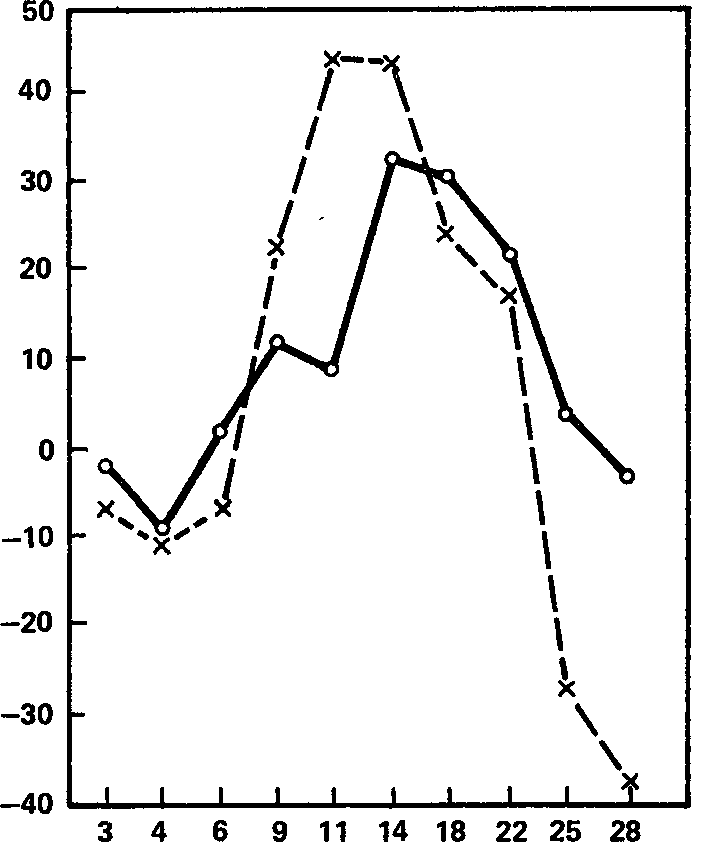
Рис. 25. Реакция навязывания ритма как функция частоты стимуляции в среднем для групп детей-десятилеток с высокой (сплошная линия) и низкой (штриховая линия) динамичностью возбудительного процесса. Ось абсцисс – частота стимуляции (имп/с); ось ординат – эффект навязывания (усл. ед.).
Итак, влияние фактора динамичности возбудительного процесса на ЭЭГ характеристики у детей может быть ясно замечено в отношении, по крайней мере, некоторых электроэнцефалографических показателей. Этот вывод, основанный на результатах применения критерия t, подтверждается и при использовании техники корреляционного анализа.
В приведенной выше табл. 16 уже были представлены коэффициенты корреляции между рядом показателей ориентировки на звук и свет и индикаторами динамичности возбуждения. Поскольку мерой этого свойства нервной системы в данной работе является легкость замыкания связей типа «звук плюс свет» с эффекторным выражением в виде блокады альфа-ритма, корреляций с динамичностью возбуждения можно было с равными основаниями ожидать и от показателей ориентировки на звук, и от показателей ориентировки на свет. Однако опыт показывает, что индикаторы ориентировки на звук имеют довольно тесную связь с показателями динамичности, а индикаторы ориентировки на свет такой связи не обнаруживают: между показателями реактивности на световое воздействие и показателями условнорефлекторных реакций нет ни одной значимой корреляции (табл. 19).
Таблица 19
Коэффициенты корреляции между рефлекторными показателями и индикаторами фона ЭЭГ у детей (К. Войку, 1964)
| Рефлекторные показатели | Показатели фона ЭЭГ | |||||||||
| альфа-индекс | частота | энергетические индексы покоя | ||||||||
| β2 | α | θ | Δ | β2 | α | θ | Δ | |||
| Длительность блокады альфа-ритма при 1‑м предъявлении звука | _604*** | –412 * | 145 | –175 | –287 | –188 | –225 | –625 *** | –441 * | –147 |
| Число предъявлений до угашения ориентировки | –459 ** | –284 | 332 | –337 | –205 | 172 | 123 | –357 | –047 | –200 |
| Длительность блокады альфа-ритма при 1‑м | ||||||||||
| предъявлении света | –135 | 005 | –031 | –087 | –043 | 170 | 192 | –182 | 233 | 064 |
| Средняя длительность блокады альфа-ритма из 9 – 10 предъявлений света | 076 | –175 | –135 | 105 | –154 | 056 | 013 | 058 | 195 | 113 |
| Средняя длительность условнорефлекторной блокады альфа-ритма при простом подкреплении | –464 ** | –465 ** | 152 | –446 ** | 000 | 435* | 312 | –535 ** | –077 | –161 |
| Средняя длительность условнорефлекторной блокады альфа-ритма при активирующем подкреплении | –423 * | –184 | 413* | –544 ** | –211 | 191 | –044 | –530 ** | –093 | –282 |
Примечание. • р < 0,1; ** р < 0,05; ***р < 0,01.
Столь же ясное разделение между звуковой и световой ориентировками можно заметить при сопоставлении их индикаторов с показателями ЭЭГ покоя. Из табл. 19 видно, что показатели ориентировки на звук вступили в корреляцию с некоторыми фоновыми показателями, в частности с альфа-индексом и энергетическим индексом альфа-полосы, которые, как мы немного позже увидим, коррелируют с динамичностью процесса возбуждения. Показатели же ориентировки на свет с фоновыми индикаторами совершенно не коррелируют.
Таким образом, функция реактивности биотоков мозга на световой раздражитель и здесь сохраняет свою специфичность, будучи обусловлена, видимо, особым фактором церебральной организации восходящих активирующих влияний. Что же касается ориентировки на звук, то у детей обусловливающие ее процессы, очевидно, находятся в достаточно тесной зависимости от параметра динамичности возбуждения. Заметим, что эта зависимость является значительно более тесной, чем у взрослых, у которых, как мы видели выше, и длительность блокады на первый звуковой раздражитель, и в особенности скорость угашения ориентировки при повторном предъявлении звука связаны, скорее, с параметром динамичности торможения. Возможно, это обусловлено тем, что у детей при общей меньшей дифференцированности у них мозговых функций большую роль играет генеральный фактор ориентировочной блокады альфа-ритма, о существовании которого мы будем предположительно говорить позже, в гл. XI. Однако этому, видимо, противоречит наличие достаточно четко – более четко, чем у взрослых, – организованной и отграниченной функции реактивности альфа-волн на световой раздражитель.
Рассмотрим, наконец, корреляционные отношения между показателями динамичности и показателями фона детской ЭЭГ.
Из группы фоновых показателей статистически значимые корреляции с показателями динамичности (табл. 19) имеют альфа-индекс и энергетический индекс альфа-волн. Согласно характеру корреляций, существующих между этими показателями, выработка условной депрессии происходит легче у тех испытуемых, которые обладают более низким альфа-индексом и меньшими величинами суммарной энергии альфа-полосы. Точно такой же характер соотношений между этими показателями наблюдается и у взрослых.
Из фоновых показателей ЭЭГ с показателями выработки условной блокады альфа-ритма коррелируют и показатели, относящиеся к частоте биоэлектрических волн в разных частотных полосах. Судя по характеру коэффициентов корреляции, более быстрой выработки условнорефлекторной депрессии следует ожидать от тех испытуемых, у которых ниже частота бета- и тета-волн и в известной степени выше частота альфа-волн. Эти соотношения в такой же форме отмечались и при групповом анализе результатов. В менее выраженной форме они имеют место и у взрослых. Как уже говорилось, эти корреляции имеют, по-видимому, парадоксальный характер. По нашим результатам, более высокой частотой бета-волн характеризуются испытуемые не с высокой динамичностью возбуждения, как можно было бы ожидать, а с низкой динамичностью этого процесса. Что касается корреляций показателей выработки условной блокады с частотой альфа-волн, то они в общем весьма невысоки. Следует заметить, что и у взрослых зависимость между частотой альфа-ритма и динамичностью возбуждения практически отсутствует, а имеет место зависимость между частотой альфа-ритма и динамичностью тормозного процесса.
Итак, динамичностью возбудительного процесса определяется у детей-десятилеток довольно широкий комплекс электроэнцефалографических показателей, куда входят, во-первых, индикаторы ориентировочных реакций на предъявление звуковых (но не световых!) раздражителей, а во-вторых, группа показателей фона ЭЭГ, таких, как альфа-индекс, суммарная энергия альфа-полосы и частота некоторых входящих в ЭЭГ ритмов, главным образом тета-ритма. Эти показатели в своей совокупности образуют, таким образом, своеобразный синдром биоэлектрических проявлений динамичности возбудительного процесса. Синдром этот во многих своих чертах напоминает соотношения, наблюдаемые на взрослых.
О чем может говорить этот факт? Во-первых, очевидно, о надежности получаемых в эксперименте данных. Во-вторых, видимо, о том, что ЭЭГ метод в равной степени и у взрослых, и у детей пригоден для изучения свойств нервной системы; с помощью этого метода у детей, так же как у взрослых, можно получить большое число количественных показателей, дающих при определении соотношений между ними ценную нейрофизиологическую информацию. Хотя в данной главе речь шла только о динамичности возбудительного процесса, нет никаких оснований думать, что ЭЭГ метод не может быть использован для исследования любых других свойств нервной системы ребенка и для того, чтобы пролить хотя бы частичный свет на одну из наиболее интересных и сложных проблем дифференциальной психофизиологии – проблему роли онтогенетического развития в формировании окончательной организации свойств нервной системы взрослого человека.
Дата: 2018-12-28, просмотров: 761.