| Статистические показатели | Частоты | Общий | ||||
| тета | альфа | бета | ||||
| общий | 9–10 кол/с | 11 – 12 кол/с | ||||
| Средние для группы с высокой динамичностью торможения (6 человек) | 119 | 398 | 151 | 247 | 123 | 640 |
| Средние для группы с низкой динамичностью торможения (6 человек) | 30 | 231 | 36 | 195 | 100 | 361 |
| Разность средних | 89 | 167 | 115 | 52 | 23 | 279 |
| Критерий / | 7,64 | 3,44 | 4,80 | 1,66 | 1,37 | 4,72 |
| Вероятность нулевой гипотезы р | <0,001 | <0,01 | <0,001 | >0,05 | >0,05 | <0,001 |
Таким образом, если деление по признаку динамичности возбуждения выявляет различия в навязывании главным образом более высоких частот (от 11 до 22 имп/с), то деление по динамичности торможения приводит к выявлению различий в основном в зоне низкочастотной стимуляции, охватывающей полосы тета- и альфа-ритмов, причем по второму признаку испытуемые дифференцируются гораздо отчетливее. Статистический анализ значимости отмеченных различий указывает, что они в большинстве своем не являются случайными и особенно это относится к различиям по динамичности тормозного процесса. Оценка этих различий дана в табл. 13, из которой видно, что разность средних ЭИНР для низких частот стимуляции (до 10 имп/с включительно) статистически весьма значима, а разность средних ЭИНР для более высоких частот (11 имп/с и более) не достигает уровня значимости.
Различия при низких частотах стимуляции столь велики, что разность общих ЭИНР (суммарных для всех применяемых частот) тоже оказывается статистически высокозначимой. Оценка различий ЭИНР в зависимости от динамичности возбуждения (табл. 14) показывает, что эти различия большей частью либо значимы на минимальном 5 %-ном уровне, либо вовсе незначимы, как, например, при сравнении общих ЭИНР для альфа-полосы.
| \ Статистические \ показатели | Частоты | Общий | ||||
| тета | альфа | бета | ||||
| общий | 9–10 кол/с | 11–12 кол/с | ||||
| Средние для группы с высокой динамичностью возбуждения (6 человек) | 26 | 322 | 84 | 238 | 112 | 460 |
| Средние для группы с низкой динамичностью возбуждения (6 человек) | 60 | 358 | 21 | 337 | 268 | 680 |
| Разность средних | 34 | 36 | 63 | 99 | 156 | 220 |
| Критерий t | 2,77 | – | 4,49 | 1,47 | 3,05 | 2,61 |
| Вероятность нулевой гипотезы р | <0,05 | – | <0,01 | >0,05 | <0,05 | <0,05 |
На основании статистического анализа можно сделать следующие выводы:
1) от динамичности тормозного процесса весьма существенным образом зависит навязывание частот тета-полосы и граничащих с Статистическая оценка различий между средними суммарными индексами навязывания ритма для групп с высокой и низкой динамичностью возбудительного процесса ними частот зоны альфа (9 – 10 имп/с); навязывание частот бета-полосы (в изучаемом нами диапазоне 16 – 22 имп/с) и прилежащих к ним частот зоны альфа, как показывает статистическая обработка, от динамичности торможения не зависит;
2) зависимость навязывания ритма всех частот от динамичности возбудительного процесса выражена вообще гораздо слабее и при статистической оценке находится в среднем где-то на грани значимости;
3) направление зависимостей в целом таково, что высокие индексы навязывания низких частот принадлежат главным образом лицам с высокой динамичностью торможения, а высокие индексы навязывания высоких (в исследованных пределах) частот свойственны, как правило, – правда, с меньшей определенностью – лицам с низкой динамичностью возбуждения. На языке традиционных терминов это означает, что лучшее навязывание всех частот чаще наблюдается у «неуравновешенных» индивидов, характеризующихся преобладанием тормозного процесса.
Факт, говорящий о связи между навязыванием ритма в полосе низких частот и динамичностью торможения, может быть сопоставлен с данными различных авторов, указывающих на связь между биоэлектрической активностью в полосе медленных ритмов, и в частности тета-ритма, и развитием коркового торможения при выработке угашения (Р. Эрнандес-Пеон, 1962; Г. Гасто, А. Роже и др., 1957), дифференцировки (Г. Гасто, А. Роже и др., 1957; Н. Иошии и др., 1962; М.Я. Рабинович, 1961) или запоминания (А. Юс и К. Юс, 1954; Н. Иошии и др., 1962), а также при введении наркотических веществ (Т.Ш. Хапажев, 1962). Некоторые авторы (Н.Н. Данилова, 1959; Н.Н. Зислина, 1955) сообщают и о связи между развитием торможения и навязыванием в области медленных ритмов. Наши данные стоят здесь несколько особняком, так как они касаются не развития тормозного состояния, а индивидуальных различий в динамичности торможения. Однако основа для толкования приведенных литературных и наших экспериментальных данных, касающихся навязывания медленных ритмов, по-видимому, остаётся одной и той же. Как литературные, так и наши собственные данные, видимо, свидетельствуют о том, что медленная ритмика коры – как фоновая, так и при навязывании ритма – есть в определенной степени выражение преобладающей тормозной тенденции.
Конкретные выводы по материалам данной главы делать, как нам кажется, еще преждевременно. Хотя и существует, видимо, прямое влияние такого параметра нервной деятельности, кан динамичность нервных (особенно тормозного) процессов, на ряд показателей электроэнцефалограммы покоя и ее изменений в уровнях ритмической световой стимуляции. Это влияние, как можно было видеть, далеко не является однозначным и однонаправленным как в отношении различных ритмов (и соответственно частот стимуляции), так и в отношении различных индикаторов, характеризующих эти ритмы.
На данном этапе исследования ЭЭГ коррелятов основных свойств нервной системы более важное значение имеет принципиальный вывод о том, что такие корреляты уже при первоначальном подходе все же удается обнаружить и охарактеризовать, пусть пока только с помощью статистических критериев. Всякое проявление индивидуальности в функциях центральной нервной Системы не может в конечном счете не определяться той или иной комбинацией свойств нервной системы. Мы глубоко убеждены, что это в полной мере относится и к динамике биопотенциалов мозга. Дальнейшая экспериментальная работа поможет вскрыть и в деталях охарактеризовать те зависимости и соотношения, которые связывают основные свойства нервной системы и индивидуальные особенности электрического ритма коры больших полушарий.
Глава 6. Динамичность возбуждения и ее электроэнцефалографические корреляты у детей[29]
Необходимость изучения свойств нервной системы у детей вытекает из стремления к генетическому подходу в исследовании высшей нервной деятельности человека, имеющему своей целью раскрытие интимных механизмов формирования как самих функций нервной системы в ходе ее развития, так и факторов, обусловливающих индивидуальные вариации проявления этих функций.
Онтогенетический анализ неврологических факторов индивидуальности дает в перспективе возможность вплотную подойти к решению таких вопросов, как относительная роль наследственного, врожденного и приобретенного в психофизиологическом облике индивида, причины возникновения той или иной недостаточности его Психофизиологической организации, и главным образом вопроса о путях и методах активного воздействия на те или иные параметры этой организации с целью преодоления ее недостатков и развития ее достоинств, – короче говоря, с целью ее оптимизации, – с учетом того, что детская нервная система, несомненно, обладает более высоким уровнем пластичности, чем уже сформировавшаяся морфологически и функционально нервная система взрослого человека.
Шаги, предпринятые различными авторами (А.Г. Иванов-Смоленский, 1935; Н.С. Лейтес, 1956 б, 1960; А.А. Волохов, А.П. Крючков, 1959; З. Н. Брике, 1956; Л.И. Уманский, 1958; И.О. Майер, 1963; В.Э. Чудновский, 1963; и др.) для изучения основные свойств нервной системы ребенка, многое прояснили в сложной картине их генеза, взаимоотношений и внешних проявлений, однако думать, что разработка этой комплексной проблемы близка к своему завершению, еще рано. В частности, еще далек от своего разрешения вопрос, с которого, собственно, следует начинать, – вопрос о выборе адекватной методики исследования свойств детской нервной системы.
Наиболее широкое хождение здесь имеет методика двигательных реакций на речевом подкреплении, но эта методика становится тем менее пригодной, чем старше возраст испытуемых детей, чем большей становится с возрастом роль тормозящих и регулирующих влияний со стороны второй сигнальной системы; кроме того, эта методика непригодна для определения силы нервной системы, т. е. одного из важнейших ее свойств. Другие же методики сколько-нибудь значительного распространения не получили, и, таким образом, задача создания комплекса приемов для определения свойств нервной системы у детей фактически до сих пор не решена. Очевидно, принципиальные основы разработки такого методического комплекса должны быть те же, что и для взрослых: методики или, по крайней мере, индикаторы должны быть непроизвольными, они должны иметь количественное выражение и должны удовлетворять требованиям «валидности» и статистической надежности.
Конкретные подходы к решению этой большой методической задачи могут быть, конечно, различными. В настоящей главе мы изложим первые результаты попыток использовать для этой цели возможности, предоставляемые электроэнцефалографической методикой, которая на взрослых, как показывает опыт, изложенный в предшествующих главах, может быть с успехом использована в качестве основы для разработки целого ряда показателей свойств нервной системы.
Электроэнцефалографическая методика имеет свою возрастную специфику, которая определяется тем, что в процессе онтогенетического развития и созревания мозга частотно-амплитудная структура его биоэлектрической активности претерпевает закономерные изменения. При этом основная тенденция заключается в увеличении частоты доминирующего ритма и в повышении его регулярности. Этот факт можно считать твердо установленным, однако имеется лишь небольшое количество данных по вопросу о возрастных особенностях той функции, на которой основаны описанные выше приемы определения свойств нервной системы у взрослых, – функции блокады обычно доминирующего альфа-ритма при различных сенсорных воздействиях; и совсем мало данных, содержащих точные количественные показатели возрастного изменений этой функции.
Работа И.В. Равич-Щербо и дипломницы М.К. Трифоновой, на результатах которой мы сейчас кратко остановимся, имела своей основной целью как раз определение возрастной динамики реактивности ЭЭГ в ответ на предъявление некоторых сенсорных; раздражений, а кроме того, и выяснение онтогенеза некоторых параметров ЭЭГ покоя, таких, как альфа-индекс, частота и суммарная энергия отдельных биоэлектрических ритмов (1959).
В этой работе были исследованы три возрастные группы ‑5, 10 и 13 лет. Авторы рассматривают (на наш взгляд, вполне справедливо) биоэлектрические составляющие детской ЭЭГ как прямые генетические проекции соответствующих частот ЭЭГ зрелого мозга – к альфа-ритму они относят частоты от 8 до 13 кол/с, к тета-ритму – от 4 до 7 кол/с и т. д. Производя с помощью полосовых фильтров автоматический анализ ЭЭГ и подсчитывая показатели частотных полос по трем указанным возрастам, И.В. Равич-Щербо и М.К. Трифонова нашли, что средняя частота и суммарная энергия в пределах альфа- и бета-диапазонов с возрастом растут, а в пределах тета-диапазона, наоборот, уменьшаются; в то же время показатели альфа-индекса имеют явную тенденцию к уменьшению – в среднем от 71,3 в 5‑летнем возрасте до 48,3 в возрасте 13 лет.
Что касается реакций ЭЭГ на различные сенсорные воздействия, то здесь оказалось, что зрительные раздражители у всех детей, которым они пятикратно предъявлялись, т. е. у детей с достаточно выраженным альфа-ритмом, вызывали неизменную реакцию десинхронизации, причем средняя длительность ее у старших детей была значительно выше, чем у пятилеток, составляя более 10 с.
Реакция десинхронизации при подаче звука была в целом менее выраженной, колеблясь в среднем по своей длительности от одной до двух с небольшим секунд и к тому же имея у многих детей тенденцию к угашению; возрастной анализ показал, что большие значения длительности десинхронизации и меньшая склонность к угашению этой реакции при пятикратном предъявлении звука имеют место у 13‑летних детей. Авторы отмечают также значительное сокращение с возрастом латентных периодов реакции десинхронизации на зрительные стимулы.
Большинство данных, приводимых И.В. Равич-Шербо и М.К. Трифоновой, как это отмечают и сами авторы, видимо, свидетельствует о том, что в исследованном ими возрастном диапазоне происходит постепенное развитие свойства динамичности возбудительного процесса; одновременно, с созреванием синаптических передач, повышается скорость проведения возбуждения по структурам ретикулярной формации и сокращаются латентные периоды реакции десинхронизации, что может говорить о повышении уровня лабильности нервной системы.
Важно отметить, что уже начиная с 5‑летнего возраста (а может быть, и еще раньше) у детей может быть зарегистрирована достаточно устойчивая реакция подавления доминирующего ритма в ответ на предъявление зрительных раздражителей. Это дает возможность, используя эту реакцию в качестве «безусловной», подкрепляющей, так же как и у взрослых, вырабатывать на ее основе условные реакции и манипулировать ими с целью получения референтных, а не только косвенных индикаторов различных свойств детской нервной системы. И.В. Равич-Щербо и М.К. Трифонова не ставили себе специальной целью разработку таких индикаторов, однако они показали, что в детской ЭЭГ для этого имеется достаточно хорошая основа. В работе же К. Войку (1964) задача получения количественного индикатора одного из свойств нервной системы, именно динамичности возбуждения, была исходной и одной из центральных. От успеха в ее решении зависело выполнение других задач, стоявших перед автором. Таких задач было несколько: а) выяснить роль динамичности возбудительного процесса в протекании электрокорковых ориентировочных реакций, б) установить возможные зависимости между динамичностью возбуждения и рядом показателей детской электроэнцефалограммы покоя (фона ЭЭГ), в) изучить зависимость между динамичностью возбуждения и показателями реакции навязывания ритма как функции частоты и интенсивности световой стимуляции.
Кроме того, представляло интерес выяснение особенностей соотношения фоновых показателей ЭЭГ между собой, а также соотношения реакций ЭЭГ, вызванных стимулами различных сенсорных модальностей и различных сенсорных характеристик.
Методика этой работы была в общем почти полностью подобна той, которая применялась нами в работе со взрослыми и описана выше, поэтому мы не будем на ней останавливаться. Укажем только, что в работе использовались полосовой анализатор частот с пятью фильтрами и шестиканальный интегратор и что программа работ включала определение реакции ЭЭГ на звуковые и световые стимулы, а также их сочетания и на мелькающие световые раздражители. Была принята следующая последовательность опыта:
1) угашение ориентировки на звук, 2) испытание действия светового раздражителя, 3) выработка условнорефлекторной депрессии альфа-ритма с простым подкреплением, 4) выработка условнорефлекторной депрессии альфа-ритма с активирующим подкреплением, 5) навязывание ритма при варьировании частоты и интенсивности стимула, 6) определение фоновых показателей ЭЭГ.
Эта процедура сохранялась одной и той же для всех испытуемых – 20 детей (мальчиков и девочек), школьников в возрасте 10 – 11 лет.
Полученные в численном выражении результаты подвергались статистическому анализу, проведенному при помощи электронно-вычислительной машины.
Сначала остановимся вкратце на тех результатах, которые были получены при внутренних сопоставлениях фоновых параметров ЭЭГ и реакции ЭЭГ на сенсорные стимулы, безотносительно к проблеме их возможной зависимости от динамичности возбудительного процесса.
Корреляции между показателями ЭЭГ покоя даны в табл. 15. Отметим, прежде всего, корреляцию между альфа-индексом и частотой тета-ритма: большему числу альфа-колебаний в фоне соответствует большая частота тета-ритма. Положительная, но незначимая связь наблюдается также между альфа-индексом и частотой бета-ритма (полоса 14 – 20 кол/с) и дельта-ритма, а корреляция альфа-индекса с частотой альфа-ритма имеет отрицательный знак.
Таблица 15
Интеркорреляции фоновых показателей детской ЭЭГ
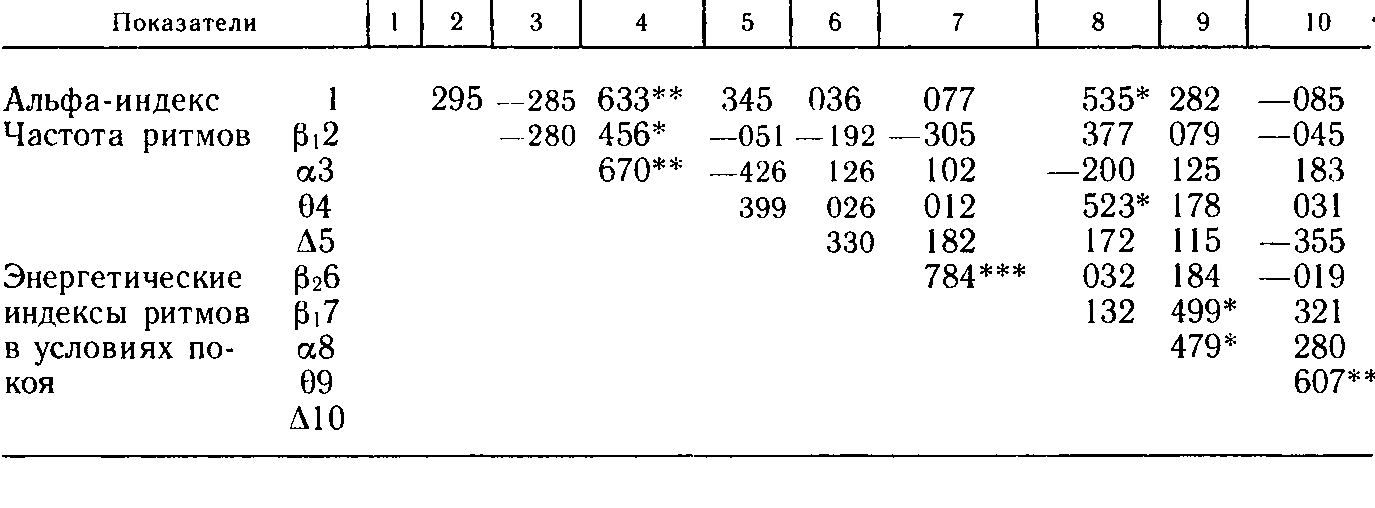
Примечание. • р < 0,05; ** р < 0,01; ***р' < 0,001. 150
Заслуживают внимания соотношения, наблюдаемые между показателями частоты биоэлектрических ритмов в частотных полосах Pi (14 – 20 кол/с), а, 9 и А. Частота бета-волн коррелирует положительно с частотой альфа-ритма. Частота альфа-ритма коррелирует отрицательно с частотой тета-ритма и проявляет тенденцию вступить в отрицательную корреляцию с частотой дельта-волн. Таким образом, частота альфа-волн коррелирует отрицательно с частотой всех других ритмов, а все другие показатели частоты проявляют заметную тенденцию к положительной корреляции (исключением является взаимная корреляция бета- и дельта-ритмов). Показатели суммарной биоэлектрической энергии разных частотных полос в состоянии покоя коррелируют между собой по принципу соседства. На очень высоком уровне значимости коррелируют показатели суммарной энергии в частотных полосах р2 (21 – 23 кол/с) и Pi, но они слабо коррелируют с энергетическими индексами других частотных полос. Правда, суммарная энергия Pi коррелирует с энергетическим индексом частотной полосы тета-ритма на уровне р < 0,05. Энергетический индекс полосы альфа-ритма коррелирует только с суммарной энергией тета-диапазона (р < 0,05).
Последний показатель коррелирует, таким образом, с показателями суммарной энергии всех других ритмов.
Что можно сказать по поводу соотношений, наблюдаемых между показателями ЭЭГ покоя?
Пожалуй, наибольший интерес вызывает здесь характер корреляций между показателями частоты биоэлектрических составляющих ЭЭГ: дельта-, тета-, альфа- и бета-ритмов. Все эти показатели (кроме крайних – дельта- и бета-частот) на том или ином уровне значимости коррелируют между собой и, таким образом, обусловливаются, видимо, действием одного и того же фактора мозговой деятельности, однако, как уже отмечалось, частота альфа-ритма с частотами других ритмов коррелирует отрицательно. Следовательно, фактор, лежащий в основе корреляций между частотными показателями ритмов, имеет биполярную природу; на одном конце его находится альфа-ритм, а на другом – все другие ритмы, как более медленные, так и более частые (во всяком случае, до 20 кол/с). Этот факт представляется чрезвычайно интересным. По-видимому, он не является случайным или специфическим только для детского возраста, так как подобные же соотношения наблюдаются и у взрослых (как показано в предыдущей главе). По данным настоящей работы, различие между частотами альфа-ритма и других ритмов сохраняется и при коррелировании частотных показателей с индикаторами рефлекторной динамики (табл. 19). Корреляции последних с частотой альфа-ритма имеют положительный знак, а с частотами других составляющих – отрицательный (это не относится к показателям ориентировки на свет, корреляции которых с частотами ритмов колеблются около нуля).
Обобщая материалы наблюдений над взрослыми и детьми, можно высказать общее предположение о том, что, вероятно, существуют раздельно работающие мозговые генераторы частот зоны альфа и частот остальных диапазонов, но что в то же время эти генераторы работают согласованно и – настолько, насколько это может быть вскрыто статистическим подходом, – вероятно, по принципу вычитания, реципрокно: повышенной активности (и большей частоте на выходе) одного генератора соответствует (статистически) пониженная активность (и меньшая частота на выходе) другого генератора. Дальнейшие исследования, с привлечением более широкого материала для сопоставлений, помогут пролить свет на эту проблему.
Довольно сложны отношения между энергетическими индексами исследованных диапазонов частот. Эти индексы коррелируют большей частью по принципу соседства, однако индексы альфа- и низкочастотного бета-ритма между собой практически не взаимосвязаны. Возможно, конечно, что этот момент имеет чисто случайное происхождение, и тогда представляется, что энергетические индексы всех ритмов ЭЭГ покоя обусловлены действием одного и того же фактора. Но возможно также, что высокая корреляция показателей суммарной энергии двух бета-полос (0,784) и отсутствие связи этих показателей с энергетическими индексами других диапазонов (кроме корреляции β1 и θ) указывают на существование одного фактора, обусловливающего энергетические показатели высокочастотных ритмов, и что относительно высокие взаимные корреляции индексов более медленных ритмов говорят о существовании другого фактора, обусловливающего энергетические показатели зоны низкочастотных ритмов.
Не лишены интереса корреляции между показателями частоты биоэлектрических составляющих и альфа-индексом. Во-первых, направление связей между этими параметрами снова отражает различия в отношениях частот альфа-ритма и других частотных диапазонов ЭЭГ: корреляция альфа-индекса с частотами (β1 и θ-, Δ-ритмов положительная, а с частотой альфа-ритма отрицательная, хотя и незначимая (напомним, что отрицательные соотношения между частотой и индексом альфа-ритма наблюдаются также и у взрослых). Во-вторых, наблюдается довольно высокая степень связи между альфа-индексом и частотой тета-ритма.
Если учесть, что альфа-индекс хорошо себя проявляет в качестве индикатора баланса нервных процессов по динамичности и, в частности, ниже у тех лиц, которые обладают высокой 'динамичностью возбуждения (по ЭЭГ показателям), то следует ожидать, что у испытуемых с высокодинамичным возбудительным процессом частота тета-ритма тоже будет ниже, чем у испытуемых с низкой динамичностью возбуждения. Далее мы увидим, что эксперимент вскрывает именно такого характера отношения между частотой тета-ритма и показателями динамичности возбуждения. То же самое, кстати, наблюдается и у взрослых.
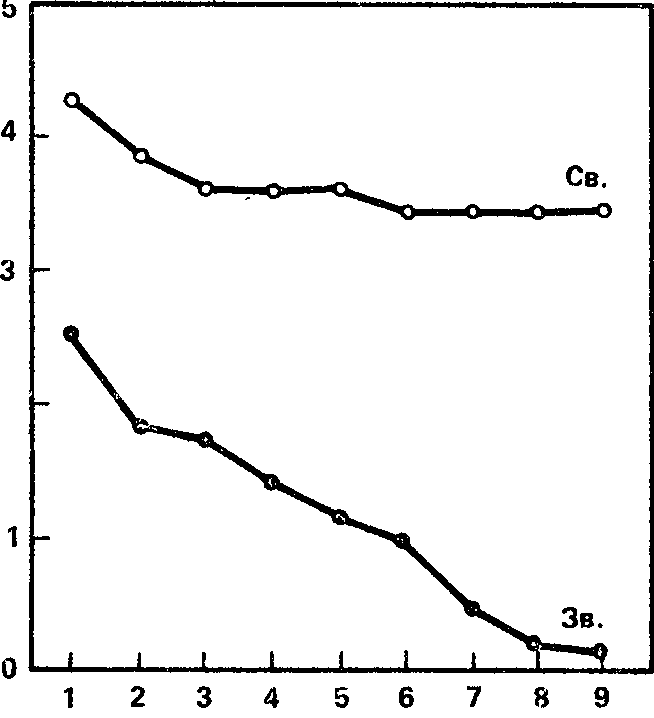
Рис. 24. Динамика ориентировочных элек трокорковых реакций у детей-десятилеток (в среднем для всей выборки). Ось абсцисс – порядковый номер предъявления раздражителей; ось ординат – длительность реакции десинхронизации (в с).
Динамика ориентировочных реакций на звук и свет (в среднем для всего контингента испытуемых детей) показана на рис. 24, где на оси абсцисс отложен порядковый номер предъявления (от 1 до 9), а на оси ординат – средняя длительность блокады альфа-ритма. Как видно из графика, максимальная длительность блокады альфа-ритма на звук наблюдается при первом предъявлении стимула. Потом, по мере повторения стимула, длительность блокады постепенно уменьшается и приближается к нулю. Процесс снижения длительности ориентировочной реакции начинается уже со второгоч предъявления стимула. Все это весьма близко к динамике ориентировочной реакции на звук у взрослых, за исключением того факта, что абсолютные значения длительности реакции активации у взрослых, особенно при первых предъявлениях, все же. значительно выше.
То же можно сказать и о реакции на свет. Максимальная длительность депрессии на свет (как и на звук) получается при первом предъявлении светового раздражителя. При следующих предъявлениях длительность реакции постепенно снижается, но гораздо менее круто, чем на звук, и поэтому в общем кривая является очень пологой и далеко не достигает оси асбцисс. Судя по степени наклона кривой, можно думать, что для получения полного исчезновения реакции активации на свет (если оно вообще возможно) необходимо намного больше повторных предъявлений раздражителя, чем это было в работе и чем требуется при применении звуковых раздражителей.
Анализ коэффициентов корреляции индикаторов ориентировочной деятельности (табл. 16) показывает, что индикаторы ориентировки на звук хорошо, на 1 %-ном уровне, коррелируют между собой, но совершенно не коррелируют с показателями ориентировки на свет. Кроме того, показатели ориентировки на свет на 5 %-ном уровне коррелируют между собой, но не коррелируют с показателями ориентировки на звук.
Таблица 16
Дата: 2018-12-28, просмотров: 701.